基于高通量测序的鲭鱼加工副产物固态厌氧发酵过程分析
2022-08-23李红玉齐祥明毛相朝郭晓华
王 瑶,李红玉,齐祥明,,毛相朝,董 浩,郭晓华
1. 中国海洋大学 食品科学与工程学院,山东 青岛 266003
2. 山东美佳集团有限公司,山东 日照 276826
鲭鱼 (Pneumatophorus japonicus) 又称鲐鱼,因富含蛋白质、钙、铁等以及Omega-3不饱和脂肪酸[1]且价格低廉而广受欢迎。自20世纪90年代起,鲭鱼已成为中国近海捕捞的主要经济鱼种之一,2020年捕捞量约3.9×105t,位居中国海洋鱼类捕捞量第四[2]。鲭鱼可被加工成鱼糜、鱼饼等产品,加工过程中产生的鱼排、内脏等副产物约占40%[3],如果不加以利用会造成资源浪费和环境污染[4]。鲭鱼属于青皮红肉鱼类,组织中含有大量游离组氨酸,在微生物中组氨酸脱羧酶的作用下易产生组胺,组胺摄入过多会引起头晕、恶心等中毒症状[5],因此鲭鱼加工副产物的利用受到了极大限制。
研究发现特定的乳杆菌能抑制组胺生成[6-7]、降低组胺含量[5-6,8],然而有关工业生产环境下 (尤其是不灭菌状态下) 采用乳杆菌进行鲭鱼加工副产物发酵的具体过程却少见报道。同时,乳杆菌可降解大分子蛋白质,形成多肽,提高食品营养价值,使食品产生独特的风味[9],并有一定的抑菌效果,抑制食品腐败变质[10]。尽管在鱼茶[6]、臭鳜鱼[11]等传统发酵食品中不灭菌发酵工艺已被广泛采用,并实践证明足够安全,但目前新开发工艺中仍很少采用该工艺,这为新型固态发酵产品的开发带来了较大的成本压力。为此,本实验选取3株代表性乳杆菌株,以豆粕为基料,在不灭菌情况下对鲭鱼加工副产物进行固态厌氧发酵,尝试生产饲料用新型活性蛋白源。实验系统监测了发酵过程中组胺、酸溶蛋白、pH 和挥发性盐基氮 (Total volatile basic nitrogen, TVB-N) 等理化指标的变化,对发酵效果较优的一组通过高通量测序进行了微生物群落分析,以探索发酵过程的生物安全性及一些风味的形成机理;对比分析了发酵前后原料及产品的脂肪酸、游离氨基酸组成和抗原蛋白降解情况,为易产组胺鱼类加工副产物的绿色综合利用与乳杆菌发酵提供工业可行的技术方案和相关基础数据支撑。
1 材料与方法
1.1 材料与试剂
鲭鱼加工副产物由山东美佳集团有限公司提供,为去头鲭鱼生产鱼片时弃除的鱼排,主要包括鱼主骨、鱼尾及其上残留的少量鱼肉。发酵所用乳杆菌 [嗜酸乳杆菌 (Lactobacillus acidophilus) HSCCLA011,L1;植物乳杆菌 (L. plantarum) HSCCLP121,L2;贝氏乳杆菌 (L. beijerinck) HSCCLB005,L3]由郑州百益宝生物技术有限公司提供;豆粕为市售豆粕;硫酸钾、硫酸铜、硼酸、盐酸等试剂均为市售分析纯。
1.2 实验方法
1.2.1 样品处理
将鲭鱼加工副产物采用绞肉机绞碎后与豆粕以质量比1∶1混合均匀后作为固态发酵基质备用。
1.2.2 菌液活化
将保存在 −20 ℃ 的冻干菌粉,加入 100 mL 的无菌温水中,放入37 ℃的培养箱中活化3 h,结合前期实验探索,确定以 1×106CFU·g−1的接种比例接入发酵基质。
1.2.3 发酵过程
上述发酵基质分别接入乳杆菌L1、L2、L3,以500 g每袋厌氧封口,并放置于30 ℃的恒温培养箱中进行固态发酵,发酵周期为30 d。灭菌组先经过121 ℃、30 min灭菌处理后接入乳杆菌L2。发酵过程中分别于第1、第3、第5、第7、第14、第21、第30天取样检测组胺、pH和TVB-N;在第1、第7、第14、第21、第30天取样检测酸溶蛋白。
1.2.4 发酵产品分析
选发酵效果较优的试验组,在发酵前和第30天取样作为原料和产品,检测其中的游离氨基酸、不饱和脂肪酸、抗原蛋白,并进行对比分析。在第1、第7、第21、第30天检测大豆球蛋白(Glycinin) 和 P-伴大豆球蛋白 (P-conglycinin) 等常见大豆抗原蛋白。在发酵第1 (为获得尽可能多样化的微生物群落)、第30和第270天 (考察产品不灭菌保藏的可行性) 取样进行高通量测序。
1.3 分析检测方法
1.3.1 酸溶蛋白检测
参考GB/T 22492—2008《大豆肽粉》以及肖志明等[12]的酸溶蛋白检测方法。
1.3.2 TVB-N 的检测
参考 GB 5009. 228—2016《食品安全国家标准食品中挥发性盐基氮的测定》和沈颖莹等[11]的TVB-N检测方法。
1.3.3 组胺的检测
参考 GB 5009. 208—2016《食品安全国家标准食品中生物胺的测定》中的分光光度法。
1.3.4 抗原蛋白检测
参考李旺军等[13]的抗原蛋白检测方法。
1.3.5 pH 检测
取 1 g样品加入 9 mL 水浸提 1 h,用 pH 计检测。
1.3.6 游离氨基酸组成分析
参考 GB 5 009.124—2016《食品安全国家标准食品中氨基酸的测定》的测定方法。
1.3.7 脂肪酸的氧化程度
脂肪酸检测参考王建和林秋萍[14]的方法。取样品10 g,冻干,于锥形瓶中加入氯仿-甲醇溶液(V∶V=4∶1)共100 mL密封,于55 ℃水浴浸提4~5 h,再抽滤获得有机浸提液,旋蒸后获得鱼油,最后用无水亚硫酸钠去除鱼油中的水分备用。
1.3.8 DNA 提取、聚合酶链式反应 (PCR) 扩增及高通量测序分析
DNA的提取参考王兴春等[15]的十六烷基三甲基溴化铵提取方法;PCR扩增方法以及引物对应区域参考文献 [16-17]的方法;高通量测序分析参考文献 [18-19]的方法。
1.4 数据处理
采用 Excel 2010、Origin 2016 软件对数据进行分析处理。数据均为3次平行。
2 结果与分析
2.1 不灭菌鲭鱼加工副产物固态发酵过程理化指标分析
有研究显示有些乳杆菌具有抑制组胺生成的能力[6-7]。因此,本研究选用3株乳杆菌进行鲭鱼加工副产物的发酵。为结合工业实际操作需求,发酵过程采取原料不灭菌工艺,发酵过程中的接种量、温度、原料配比等已在前期实验进行了充分探索。3株乳杆菌和未接种对照组的组胺、酸溶蛋白、TVB-N和pH等理化指标变化见图1。其中组胺为鲭鱼副产物发酵产品理化安全性评价的关键指标,TVB-N为理化安全性评价的通用指标。
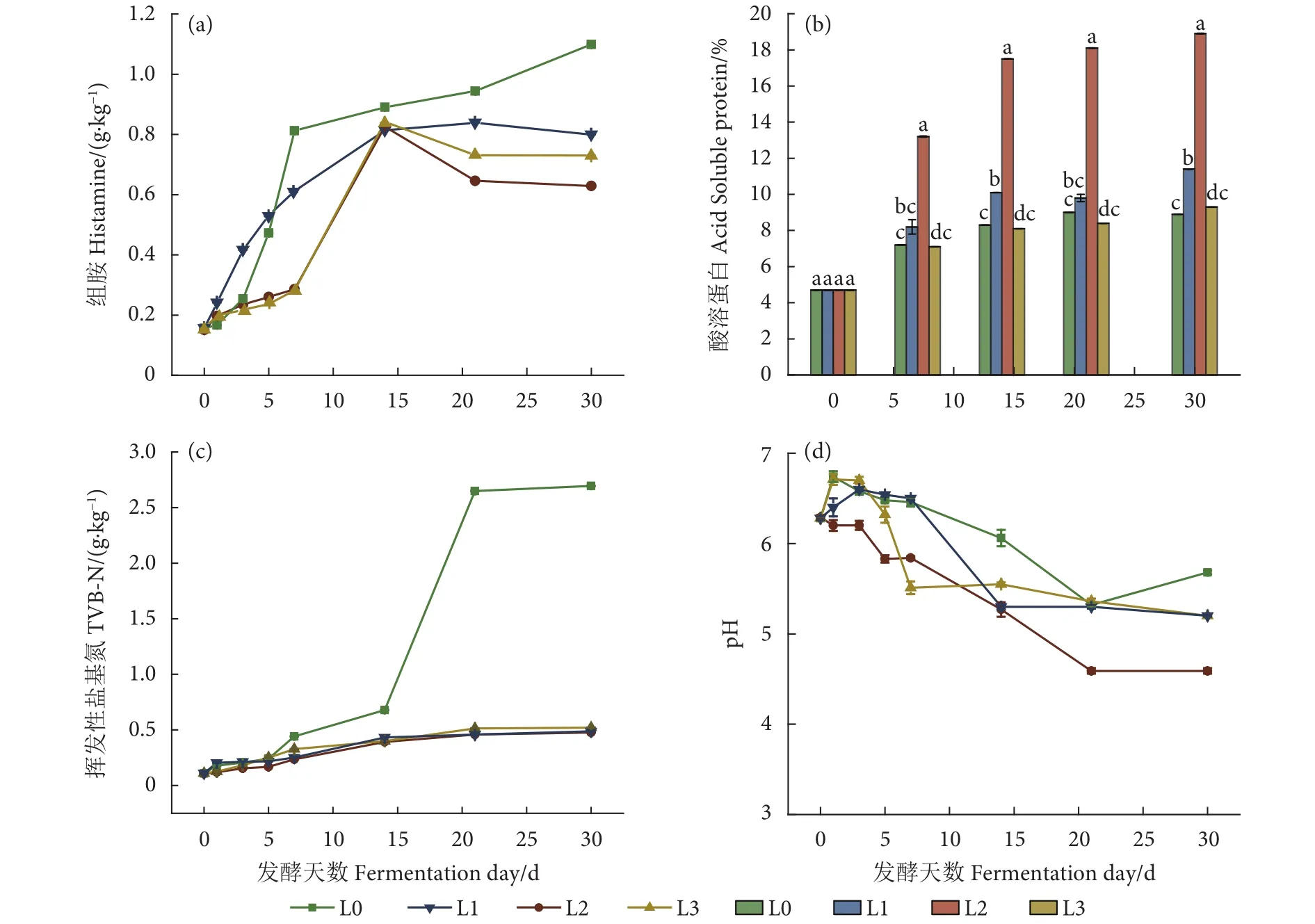
图1 不同乳杆菌菌株发酵对鲭鱼加工副产物理化指标的影响注:L0、L1、L2和L3分别为无任何处理的对照组、接种嗜酸乳杆菌、接种植物乳杆菌和接种贝氏乳杆菌的实验组;不同字母之间表示存在显著性差异 (P<0.05),图2同此。Fig. 1 Effects of different lactic acid bacteria fermentation on physicochemical indexe of processed by-products of mackerelNote:L0, L1, L2 and L3 are the control group without any treatment, groups inoculated with L. acidophilus, L. plantarum and L. beijerinck, respectively. Different letters indicate significant difference (P<0.05). The same case in Fig. 2.
3株乳杆菌抑制组胺生成的能力见图1-a。对照组的组胺含量一直呈上升趋势,第7天即超过0.800 g·kg−1,第 30 天时高达到 1.09 g·kg−1,且上升趋势未受到抑制。根据GB/T 19164—2003,此时产品的该指标只与三级鱼粉相当,未来甚至会更糟。而3组乳杆菌发酵样品中,组胺生成均被不同程度抑制。第14天后L2、L3组的组胺含量出现下降,第21天后L1组出现下降,说明样品中的组胺发生了降解。其中降解效果最好的是L2组,第30天时组胺已降至 0.629 g·kg−1,接近 GB/T 19164—2003规定的一等品指标。显著性分析结果显示,在第30天植物乳杆菌组的组胺与对照组及其他乳杆菌组差异显著 (P<0.05)。
Feng等[20]和Zhang等[6]报道了盐腌鲅鱼和草鱼鱼茶产品中植物乳杆菌致使组胺或总生物胺降解的现象,Dominggoslopes等[21]研究指出副干酪乳杆菌 (L. paracasei) 具有降解组胺的能力。这些研究表明近年来一些乳杆菌尤其是植物乳杆菌,其降解组胺的能力逐渐被发现。但有关嗜酸乳杆菌和贝氏乳杆菌降解组胺的能力则未见报道,尽管从本实验结果看其降解能力不及植物乳杆菌。
酸溶蛋白是评价发酵饲料中小肽含量的重要指标[22]。本研究将豆粕作为降低发酵原料水分的重要辅料,拟通过发酵将原料中动、植物蛋白降解为小肽成分。发酵过程中酸溶蛋白质量分数的变化见图1-b。结果显示,在整个发酵过程中,L1、L3组的酸溶蛋白质量分数与对照组相比增幅较小;而L2组与对照组及其他乳杆菌组比较均有显著差异(P<0.05),发酵第14天时酸溶蛋白质量分数达到17.5%,第30天达18.9%。从酸溶蛋白的增长趋势看,发酵14 d后增长缓慢,而从组胺抑制或降解效果看,发酵时间有必要延长至30 d。近年也有利用乳酸菌发酵豆粕尝试获得较高的酸溶蛋白含量的报道,但效果不理想,如谢全喜等[23]报道获得的酸溶蛋白质量分数仅为10.5%。本实验获得了较高的酸溶蛋白,可能是因为鱼蛋白更易被降解或菌种选择得当。
对照组从第14天开始TVB-N指标急剧上升,第 21 天时超过了 2.50 g·kg−1(图1-c),且气味表现出严重的氨味和明显的不愉快气息,显示对照组腐化变质严重。第30天,对照组的TVB-N与乳杆菌发酵组的差异极显著 (P<0.01)。乳杆菌发酵样品中的 TVB-N 均得到了有效控制 (仅约 0.50 g·kg−1),远低于GB/T 19164—2003中的特级品要求 (≤1.10 g·kg−1),且3组均无氨臭味,而是酱香气息中略带清新愉悦的酸味。上述结果说明乳杆菌能在一定程度上减轻氨基酸受到的破坏,减少氨和胺类物质的产生,并产生较好的风味。有研究指出接种乳酸菌发酵有利于形成具有清新感觉的气味活性化合物[24],这与本实验结果相似。然而其中的酱香气息并非乳酸菌特有的风味,也未见类似现象的报道;其形成原因有待探索。本实验同时检测了发酵过程中的pH变化 (图1-d)。3个发酵组的pH较对照组降低程度更高,其中以L2组降幅最大。
综上分析,各发酵组产品的理化安全指标能满足相关要求。以上结果表明,植物乳杆菌在降低产品组胺含量、降解蛋白质成小肽以及降低pH方面,均优于另外两种乳杆菌,为此后文对其作重点分析。
2.2 与灭菌发酵过程的对比
为进一步分析不灭菌发酵过程与灭菌发酵过程的差异,本研究对比考察了灭菌不接种乳杆菌(S0)、灭菌接种乳杆菌 (S1)、不灭菌接种乳杆菌(S2) 的酸溶蛋白、组胺、TVB-N和pH等指标的变化。结果意外发现,灭菌后的两组酸溶蛋白增量非常有限;不灭菌组的酸溶蛋白在整个发酵过程中保持强劲的增加趋势,与不灭菌的两组相比有显著差异 (P<0.05) (图2)。推测未灭菌原料中可能同样含有降解蛋白的酶类或微生物,这可能是本实验采用乳杆菌获得的酸溶蛋白量比谢全喜等[23]更高的另一原因。综合图2-a和图1-b中灭菌接种组和不灭菌接种组结果,发现这些内源酶或微生物与接种植物乳杆菌对蛋白的降解作用可能存在协同效应。上述发现尚未见报道。

图2 灭菌与不灭菌发酵对鲭鱼加工副产物理化指标的影响注:S0、S1、S2 分别为灭菌不接种乳杆菌、灭菌接种乳杆菌、不灭菌接种乳杆菌;组胺相对含量由每一组组胺实际含量除以第 0 天未发酵的组胺实际含量计算得出。Fig. 2 Effects of sterilization and non-sterilization fermentation on physicochemical indexes of processed by-products of mackerel Note:S0, S1 and S2 represent sterilization without inoculation of lactic acid bacteria, and non-sterilization with inoculation of lactic acid bacteria, respectively. The relative histamine content is calculated by the actual histamine content of each group by the actual histamine content of the sterilization group on 0th day.
图2-b和图2-c显示,不灭菌组的组胺和TVB-N值均高于灭菌组,推测这与灭菌组的蛋白未被充分降解有一定关系,因为组胺的产生要以组织中的游离组氨酸为底物[20],而TVB-N是以游离氨基酸为基础产生的。鲭鱼副产物中最令人关注的理化安全指标——组胺在第30天时已经从相对高位降至低于灭菌不接种组,进一步验证了植物乳杆菌具有一定的组胺降解能力。
本实验结果显示出了不灭菌发酵产品理化指标的安全性,因此灭菌组在安全指标上的优势不大。从生产实践角度看,灭菌操作在增加成本的同时降低了产品关键指标 (酸溶蛋白),显然是无法接受的。
2.3 基于高通量测序的发酵过程微生物群落分析
在不灭菌发酵过程中,微生物安全性一直是研究者关注的焦点[6,20]。有研究发现一些乳酸菌具有抑制产组胺微生物生长的能力[25-26],本实验所用的植物乳杆菌是否也有这种能力值得深究。因此,基于高通量测序对植物乳杆菌发酵组进行了不灭菌发酵过程的微生物群落分析,Alpha多样性指数结果见表1。

表1 植物乳杆菌发酵鲭鱼加工副产物的Alpha多样性指数Table 1 Alpha diversity index of processed by-products of mackerel fermented by L. plantarum
表1显示,所有样品覆盖率均为1,即操作分类单元 (Operational taxonomic units, OTUs) 的测序覆盖率均为100%,表明检测过程捕获了主要的OTUs,检测结果可信。各多样性指数显示,不接种对照组的ACE、Chao1、香农指数30 d后均上升,且ACE和Chao1指数远高于发酵组,即对照组微生物物种总数增加,初步说明鲭鱼加工副产物不经灭菌易滋生各种杂菌,从而加速腐败进程。而对照组的香农指数一直低于发酵组,说明对照组和处理组相比个体分布更不均匀,这可能因为未接种的原料中有某些优势的腐败菌生长。而发酵组数据显示,第30天时,ACE、Chao1指标出现下降,香农指数表现出比对照组更高的上升趋势,说明此时产品中的微生物物种数受到抑制而降低,但已有物种的分布均匀度还较高。
本实验进一步将样品贮存至第270天,以考察发酵产品的长期保藏性能。结果显示,ACE、Chao1、香农指数均大幅下降,即该产品在长期保存过程中样品物种总数和种类分布均匀度均有所降低,微生物生长受到进一步抑制。且发现产品在长期保存过程中乳杆菌的生长优势更明显。
本实验进一步选取了门和属水平上丰度最高的10个细菌群落做样品微生物丰富度柱状图 (图3-a和图3-b)。在门水平上,初始发酵第1天鉴定出含量较高的菌种有蓝藻菌门、变形菌门、厚壁菌门,与未接种的对照组一致。根据Zang等[27],含量最高的蓝藻菌门应是来自生鱼肉的主要菌种,门内未发现明确的危害菌类;厚壁菌门中的多数菌种为有益菌[28],本实验所用乳杆菌即属于该门;而变形菌门内的危害菌较多,如沙门氏菌、假单胞菌等均属于该门[29]。

图3 不同发酵阶段的细菌群落相对丰度注:保藏 270 d 指发酵后保存在常温下 270 d;图 4 同此。Fig. 3 Relative abundance of bacterial communities at different fermentation stagesNote:Storage of 270 d means storing at room temperature for 270 d after fermentation; the same case in Fig. 4.
30 d后,对照组中蓝藻菌门丰度基本不变,厚壁菌门有一定增加;发酵组中含乳杆菌的厚壁菌门丰度大幅上升,成为优势菌,而变形菌门丰度大幅减少 (图3-a)。对发酵产品长期保藏后 (270 d),厚壁菌门丰度上升至94.9%,占绝对优势,而变形菌门丰度进一步降低。这说明经乳杆菌发酵的产品能抑制变形菌门中危害菌的生长,因此在发酵和保藏过程中可有效防止该门危害菌的侵染。
对比图3-a和图3-b,可以认定厚壁菌门的丰度全部来自于乳杆菌属,而未分属的叶绿体菌属(UnidentifiedChloroplast) 极有可能是蓝藻门丰度的贡献者。发酵产品保存第270天时乳杆菌属丰度为92.0%,与厚壁菌门基本一致。
值得注意的是,变形菌门下并没有丰度很高的绝对优势菌,只统计到假单胞菌属(Pseudomonas)、嗜冷杆菌属 (Psychrobacter)。有研究表明假单胞菌属、嗜冷杆菌属均为产生组胺的主要菌种[30]。本研究的丰度数据显示,发酵组假单胞菌属30 d为0.645%,保存270 d后降低到0.394%;嗜冷杆菌属 30 d 为 0.075 2%,保存 270 d 后消失。说明发酵产品中组胺含量的降低除来自于自身的降解能力外,还可能来自于乳杆菌对其中产组胺菌的抑制。对照图3-b中发酵30 d的结果可知,发酵第30天时虽然乳杆菌已占优势,但其他菌种也有丰度增加的现象,这与表1中香农指数的增加一致。而经历长期贮存后,乳杆菌对嗜冷杆菌属、假单胞菌属等潜在危害菌的抑制作用会得到进一步加强。
Feng等[20]和Zhang等[6]在植物乳杆菌发酵鱼类产品实验中通过高通量测序分析了细菌群落,但所报道的其他菌类受抑制程度远低于本实验结果,这可能是由于菌种不同,也可能与原料成分不同(如盐或大米的加入) 有关。
迄今发现有关乳杆菌对真菌群落影响的报道较少,仅Liu等[31]对青贮饲料进行过类似的探索;在不灭菌发酵背景下这类分析非常必要。本研究分析了发酵过程中真菌群落的动态变化。在门水平上,担子菌门、子囊菌门为鉴定的最主要菌门(图4-a)。乳杆菌发酵组中,早期担子菌一度获得了生长优势,但长期贮存过程中出现变化,估计这与多数真菌的好氧需求有关。早期有一定氧含量时,乳杆菌可能会协同部分真菌生长,而在长期贮存过程中大量真菌进入休眠状态,群落丰富度反而有所提升。值得注意的是,著名的真菌危害菌——黄曲霉菌 (Aspergillus flavus) 即属于子囊菌门下的曲霉菌属[32]。从乳杆菌发酵30 d的结果来看,该门丰度降低,说明其中物种同样受到了抑制;而长期贮存后 (270 d),子囊菌门丰度出现大幅增加,值得进一步关注。

图4 不同发酵阶段真菌群落的相对丰度Fig. 4 Relative abundance of fungal communities at different fermentation stages
从属水平上看,长期贮存的发酵产品中不明确物种的菌种比例进一步增加 (图4-b)。对照组中属名明确、丰度在前两名的分别是耐干霉菌属 (Xeromyces) 和曲霉菌属 (Aspergillus),均属于子囊菌门[33-34]。对照组中这两个属在30 d后相对丰度轻微增加 (曲霉菌属从3.14%提升到3.17%;耐干霉菌属从3.52%提升到5.07%),但发酵组中这两属并未占优势,尤其是可能含有潜在危害菌的曲霉菌属,第1天丰度为21.5%,第30天时一度消失,第270天丰度再次轻微增至1.67%,整体呈大幅下降趋势。说明该发酵过程能抑制存在潜在危害的曲霉菌属。
另外,在属水平上,发酵组出现一些酵母属丰度大幅波动的现象。发酵第30天时,真菌中的绝对优势菌为担子菌门的丝孢酵母属 (Trichosporon)[35],占比高达76.9%;贮存第270天时,属于子囊菌门的另一种酵母属Trichomonascus则成为绝对优势真菌。上述两种酵母属为酒曲、豆豉等产品中常见的菌种,还有生香功能[36-37]。这一发现解释了前述发酵产品出现酱香的原因,之前未见类似报道。推测这一现象与酵母菌属的兼性厌氧性有关。
2.4 发酵产品游离氨基酸、脂肪酸及抗原蛋白分析
2.4.1 游离氨基酸组成
鲭鱼及其加工副产物产组胺的首要原因是鱼肉中存在大量游离组氨酸[38],从而被组胺产生菌利用生成组胺,因此本实验对产品中的游离氨基酸进行分析 (表2)。结果显示,长时间放置游离氨基酸总量均出现了一定程度的升高,多数氨基酸比例基本保持不变。然而对照组组氨酸含量有明显变化(由 2.18 降至 1.88 mg·g−1),而发酵组中组氨酸含量则略有升高。推测对照组组氨酸可能被组胺产生菌降解成了组胺,而发酵组组氨酸在生成与被降解或利用之间形成了平衡。结合图1-a发酵组的组胺变化,与菌相分析中产组胺的假单胞菌和嗜冷杆菌等虽已得到一定的抑制但尚未被完全消除的推测相互印证。结合其他研究[6-7,25],再次印证了本实验所选的乳杆菌具有抑制组胺产生菌的效果。
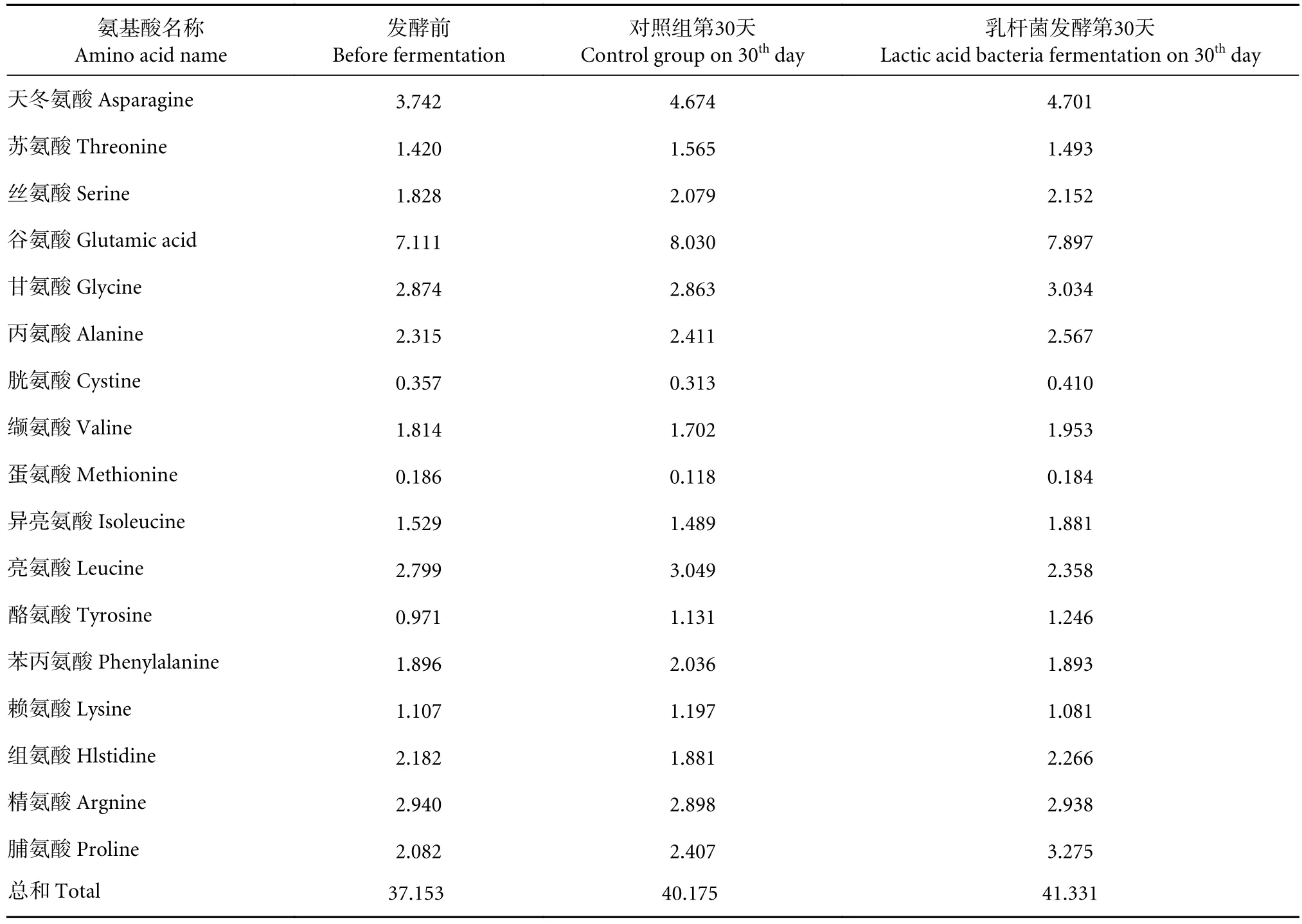
表2 乳杆菌发酵鲭鱼加工副产物游离氨基酸组成的变化Table 2 Changes of free amino acid composition of processed by-products of mackerel fermented by lactic acid bacteria mg·g−1
2.4.2 脂肪酸组成分析
鲭鱼富含脂肪,其中大量存在的不饱和脂肪酸是其重要的功能性营养成分,然而其保藏性能相对较差,易被氧化[39]。因此检测了发酵前后产品中的脂肪酸组成,结果见表3。发酵后各不饱和脂肪酸比例有小幅升高,说明未被氧化损失。这主要可能是发酵过程的厌氧环境使然,也可能是植物乳杆菌具有抗氧化能力的体现[40],还可能有乳杆菌发酵过程分解脂肪产生不饱和脂肪酸的作用[41]。

表3 乳杆菌发酵前后脂肪酸组成变化Table 3 Changes of fatty acid composition before and after lactic acid bacteria fermentation %
豆粕是本实验的主要基料,大豆中的抗原蛋白具有抗原性和致敏性[42]。因此采用十二烷基硫酸钠聚丙烯酰胺凝胶电泳 (Sodium dodecyl sulfate polyacrylamide gel electrophoresis, SDS-PAGE) 测定了发酵组和未接种对照组中豆粕抗原蛋白的降解情况 (图5)。结果显示,发酵第21天时抗原蛋白条带已消失 (分子量为22~76 kD),说明抗原蛋白如大豆球蛋白和P-伴大豆球蛋白等已被充分降解,而对照组抗原蛋白条带一直存在。

图5 乳杆菌发酵降解抗原蛋白的SDS-PAGE蛋白电泳图谱Fig. 5 SDS-PAGE electrophoretic pattern of lactobacilli protein degraded by fermentation
3 结论
本研究针对鲭鱼加工副产物的固态厌氧发酵生产饲料用新型活性蛋白源的过程,得出:
1) 发酵工艺上,植物乳杆菌HSCC-LP121在待选菌株中表现优异。综合考虑蛋白降解效果、组胺降低效果以及TVB-N等指标,发酵时间以30 d为宜,此时组胺质量分数为 0.629 g·kg−1,酸溶蛋白质量分数为 18.9%,TVB-N 为 0.476 g·kg−1,抗原蛋白条带消失。在组胺生成抑制方面,本实验所选乳杆菌均能抑制鲭鱼加工副产物中组胺的生成。
2) 不灭菌发酵的生物安全性方面,植物乳杆菌在发酵过程中能有效抑制组胺产生菌嗜冷杆菌属、假单胞菌属等潜在危害细菌及曲霉菌属等潜在危害真菌的生长。
3) 相对于灭菌发酵工艺结果,不灭菌发酵工艺的酸溶蛋白含量大幅增加,同时其组胺和TVB-N等理化安全指标也符合相关产品的国标要求,充分证明该发酵过程不灭菌的合理性。其在节能的同时保全了产品的功能性营养成分和风味,且鲭鱼副产物中不饱和脂肪酸成分基本保持不变。
长期贮存结果显示,发酵产品没有滋生杂菌或质量劣化,甚至植物乳杆菌的发酵过程还诱使多种酵母菌属在发酵及长期保存中产生了生长优势,为产品附加了酱香风味。
