不同腹脂率番鸭回肠转录组差异分析
2022-08-23王小骊王兴鑫杨彩梅肖英平吕文涛
王小骊,王兴鑫,,杨彩梅,肖英平,杨 华,吕文涛
(1.浙江省农业科学院,省部共建农产品质量安全危害因子与风险防控国家重点实验室,杭州 310021;2.浙江省农业科学院农产品质量安全与营养研究所,杭州 310021;3.浙江农林大学动物科技学院·动物医学院,杭州 311300)
番鸭原产于南美洲与中美洲热带地区,具有肉质好、脂肪少、瘦肉多、蛋白质含量高等特点。脂肪是畜禽体内重要的能量来源和能量贮备形式,但脂质氧化将影响其气味和风味,引起肉品质和营养价值急剧下降[1],但对家禽而言,超过85%的胴体脂肪是多余的并非生理需要,而腹部是脂肪的主要沉积部位[2]。过多的腹脂沉积不仅会造成饲料的浪费[3],还会影响屠宰率[4]。脂肪沉积受到脂肪酸合成酶(fatty acid synthase,FAS)和血管生成素样蛋白(angiopoietin-like proteins,ANGPTLs)等影响。研究发现,FAS的催化是脂肪酸合成中的最后一步,在脂肪代谢中起关键作用[5]。Semenkovich[6]研究表明,FAS可以通过提高甘油三酯的合成,促进动物脂肪的沉积。丰胜求[7]研究发现,Angptl1和Angptl2基因均在猪的脂肪组织中高表达。Angptl3、Angptl4、Angptl8也被证实与抑制脂肪代谢有关[8]。此外,脂肪酸结合蛋白家族(fatty acid-binding protein,FABPs)在脂肪代谢与沉积方面也起着重要作用。FABPs是一类小分子细胞内蛋白质,与细胞内脂肪酸运输有关,参与宿主体内脂肪的合成与降解[9]。王起贵[10]研究发现,鸡肝脏型-FABP(L-FABP)基因的5′-侧翼区2个多态性位点G204A和T205C与鸡的腹脂重和腹脂率有显著相关性。microRNA(miRNA)是动物体内常见的基因表达因子。通过对不同高、低腹脂含量的母鸡腹脂的miRNA表达检测及miRNA-mRNA相互作用分析发现,差异表达的miRNA包括miR-30d、miR-26a和miR-17-5p等[11]。Chen等[12]发现miR-30d与miR-30a-5p、miR-146b5p、miR-21和miR-101-3p在鸡腹部脂肪组织中大量表达,并在脂肪形成调控中发挥作用。目前对鸡的脂肪代谢方面的研究较多,但对鸭的脂肪代谢相关基因的研究比较缺乏。小肠是机体消化和吸收营养物质最重要的场所,其中位于小肠后段的回肠,在脂肪吸收的过程中发挥着作用[13-14]。因此,本试验利用转录组测序技术对高、低腹脂率的番鸭回肠进行测序分析,筛选与番鸭脂肪代谢相关的通路和基因,为进一步研究脂肪代谢途径的分子调控机制提供理论基础。
1 材料与方法
1.1 材料
1.1.1 试验动物和组织样品采集 本试验选用2 000只1日龄雄性商代种番鸭,初体重为(38.46±0.87)g,试验地点在浙江省兰溪禾旺禽业合作社,番鸭自由采食和饮水,饲粮组成与杨华等[15]试验一致。饲养70 d后,随机选取200只体况健康的番鸭屠宰后称重,采集每只番鸭的腹部脂肪并称重,同时采集所有番鸭的回肠组织,迅速置于液氮中速冻,于-80 ℃保存,以备提取样品总RNA。根据禽肉行业标准(NY/T 753—2003),计算腹脂率,将200只平均腹脂率为2.58%±0.73%的番鸭,按照腹脂率排序,取腹脂率最高的5只为H组,最低的5只为L组,对其回肠组织进行转录组分析。
1.1.2 主要试剂和仪器 TRIzol®Plus RNA纯化试剂盒和反转录试剂盒PrimeScript RT MasterMix(Perfect Real-time)均购自Invitrogen公司;RNase-free DNase Set试剂盒购自Qiagen公司;RNA Nano6000检测试剂盒购自安捷伦公司;NEBNext®UltraTMRNA Library Prep Kit和荧光定量染料试剂盒Power SYBR®Green Master Mix购自Applied Biosystems公司。
Nanohotometer®分光光度计购自IMPLEN公司;NanoDrop分光光度计购自Thermo Fisher Scientific公司。
1.2 方法
1.2.1 RNA提取与质量检测 从回肠组织中提取总RNA,使用TRIzol®Plus RNA纯化试剂盒进行纯化,并按照说明书使用RNase-free DNase Set对总RNA进行处理。使用Nanohotometer®分光光度计检测RNA的纯度,使用NanoDrop分光光度计检测RNA的浓度,使用1.0%琼脂糖凝胶电泳检测RNA样品的完整性和污染程度,使用安捷伦生物分析仪2100系统的RNA Nano 6000检测试剂盒评估RNA样本的完整性。
1.2.2 文库构建与质检 建库起始RNA为总RNA,总量≥1 μg。使用NEBNext®UltraTMRNA Library Prep Kit建库试剂盒建立文库。通过Oligo(dT)磁珠富集带有Poly A尾的mRNA,随后在NEB Fragmentation Buffer中用二价阳离子将得到的mRNA随机打断。以片段化的mRNA为模板,随机寡核苷酸为引物,在M-MuLV逆转录酶体系中合成cDNA第一条链,随后用RNaseH降解RNA链,并在DNA Polymerase Ⅰ体系下,以dNTPs为原料合成cDNA第二条链。纯化后的双链cDNA经过末端修复、加A尾并连接测序接头,用AMPure XP beads筛选250~300 bp的cDNA,进行PCR扩增并再次使用AMPure XP beads纯化PCR产物,最终获得文库。
文库构建完成后,先使用Qubit2.0 Fluorometer进行初步定量,稀释文库至1.5 ng/μL,随后使用Agilent 2100 bioanalyzer对文库的insert size进行检测,insert size符合预期后,采用实时荧光定量PCR对文库有效浓度进行准确定量,文库有效浓度的最低阈值为2 nmol/L,以保证文库质量。
1.2.3 上机测序 库检合格后,把不同文库按照有效浓度及目标下机数据量的需求筛选后,在Illumina平台上测序产生150 bp配对末端读数。测序的基本原理是边合成边测序(sequencing by synthesis)。在测序的流通通道中加入4种荧光标记的dNTP、DNA聚合酶以及接头引物进行扩增,在每一个测序簇延伸互补链时,每加入一个被荧光标记的dNTP就能释放出相对应的荧光,测序仪通过捕获荧光信号,并通过计算机软件将光信号转化为测序峰,从而获得待测片段的序列信息。
1.3 数据处理和生物信息学分析
1.3.1 参考序列比对 对过滤后的数据进行Trinity拼接处理,将Trinity拼接得到的转录组作为参考序列(Ref),采用RSEM软件将每个样品的高质量数据(clean reads)与Ref进行比对,分别过滤掉比对质量值低于10的reads,非成对比对上的reads,比对到基因组多个区域的reads。
1.3.2 差异基因表达分析 使用DESeq R包(1.10.1)进行H和L组的差异表达分析。DESeq提供统计程序,使用基于负二项分布的模型确定数字基因表达数据中的差异表达。使用Benjamini和Hochberg方法调整得到的P值以控制错误发现率。由DESeq筛选出P<0.05的基因被指定为差异表达基因。
1.3.3 GO和KEGG富集分析 采用GOseq(1.46.0)和KOBAS(KOBAS 3.0)软件对差异基因集进行GO(gene ontology)功能富集分析和KEGG(kyoto encyclopedia of genes and genomes)通路富集分析。
1.3.4 实时荧光定量PCR分析 利用实时荧光定量PCR验证番鸭回肠基因文库中RNA-Seq基因表达数据的准确性和重复性。分别提取高、低腹脂率组番鸭回肠的总RNA,利用反转录试剂盒和实时荧光定量试剂盒进行cDNA合成和基因定量检测,引物采用NCBI网站上的Primer-BLAST程序(https:∥www.ncbi.nlm.nih.gov/tools/primer-blast/index.cgi?LINK_LOC=BlastHome)进行设计,由生工生物工程(上海)股份有限公司合成,序列见表1。实时荧光定量PCR反应体系20 μL:Power SYBR®Green Master Mix 10 μL,cDNA模板1 μL,上、下游引物(10 μmol/L)各0.5 μL,ddH2O 8 μL。反应程序:95 ℃ 1 min;95 ℃ 15 s,63 ℃ 25 s,共40个循环;55~95 ℃绘制熔解曲线。每个样品重复3次,各个基因的相对表达水平以2-ΔΔCt法进行计算。

表1 引物信息
1.4 数据分析
采用SPPS 20.0中的非配对t检验进行差异显著性分析,采用GraphPad Prism 8.0软件作图,结果以平均值±标准差表示,以P<0.05和P<0.01作为差异显著性判断标准。采用R软件包进行皮尔森相关性分析。
2 结 果
2.1 腹脂与腹脂率
由表2可知,H与L组的体重差异均不显著(P>0.05),H组的腹脂重和腹脂率均极显著高于L组(P<0.01)。

表2 各组体重、腹脂重与腹脂率
2.2 RNA测序结果
10个样本中原始数据的平均值为52 472 860 bp,对原始数据进行质控处理后平均每个样本有51 479 286个高质量数据,占原始数据的98.11%。Q30平均值为91.30%。将Trinity拼接得到的转录组作为Ref,高质量数据映射到Ref的平均映射率为80.11%(表3)。

表3 番鸭回肠转录组的序列质量和比对信息
2.3 差异表达基因分析
两组样品中共检测到31 517个基因,以P<0.05和log2|FoldChange|>1为阈值,共鉴定出602个差异表达基因,其中包括285个上调基因和317个下调基因(图1)。根据每个样本差异表达基因表达水平的log2(倍数变化)值进行聚类分析,两组样品都显示出了良好的重复性(图2)。

①竖线是差异阈值的2倍,横线是P=0.05阈值。②A象限的点表示下调基因,B象限的点表示上调基因,其余的点表示非差异表达基因①The vertical line is twice the difference threshold,and the horizontal line is the P=0.05 threshold. ②The dots in quadrant A represent down-regulated genes,those in quadrant B represent up-regulated genes,and the remaining dots represent non-differentially expressed genes图1 H和L组差异表达基因火山图Fig.1 Volcano plot of differentially expressed genes in H and L groups

①色标表示RPKM归一化的log2转换计数。②单杠代表基因;垂直栏代表样品。③颜色较深表示较高表达的基因,颜色较浅表示较低表达的基因①The color scale represents the log2 conversion count normalized by RPKM. ②The horizontal bar represents genes;The vertical bars represent samples. ③Darker colors indicate genes with higher expression,and lighter colors indicate genes with lower expression图2 差异表达基因的聚类热图分析Fig.2 Clustering heat map analysis of differentially expressed genes
2.4 差异表达基因的功能富集分析
对602个差异表达基因进行了GO功能富集分析,包括生物过程、细胞成分和分子功能三大类,共有73个通路显著富集(P<0.05),筛选出5个与脂质代谢相关的条目:脂质应答(response to lipid)、类固醇激素介导的信号通路(steroid hormone mediated signaling pathway)、类固醇激素应答(response to steroid hormone)、类固醇激素刺激的细胞应答(cellular response to steroid hormone stimulus)和脂质的细胞应答(cellular response to lipid),而且富集到这5个条目的差异表达基因是相同的(表4)。

表4 与脂质代谢相关的GO条目
进一步对602个差异表达基因进行KEGG通路分析,共有16个通路显著表达(P<0.05,图3)。其中PPAR信号通路(PPAR signaling pathway)、初级胆汁酸的生物合成(primary bile acid biosynthesis)、胆汁分泌(bile secretion)和花生四烯酸代谢通路(arachidonic acid metabolism)与脂肪代谢沉积相关(表5)。

Renin-angiotensin system,肾素-血管紧张素系统;Drug metabolism-other enzymes,药物代谢-其他酶;Pentose and glucuronate interconversions,戊糖和葡萄糖酸酯的相互转化;Histidine metabolism,组氨酸代谢;PPAR signaling pathway,PPAR信号通路;Glutathione metabolism,谷胱甘肽代谢;Chemical carcinogenesis,化学致癌作用;Primary bile acid biosynthesis,初级胆汁酸的生物合成;Pyrimidine metabolism,嘧啶代谢;Taurine and hypotaurine metabolism,牛磺酸和低牛磺酸代谢;Type Ⅱ diabetes mellitus,Ⅱ型糖尿病;Arachidonic acid metabolism,花生四烯酸代谢;Protein digestion and absorption,蛋白质消化吸收;Bile secretion,胆汁分泌;Ascorbate and aldarate metabolism,抗坏血酸和醛酸代谢;Cytokine-cytokine receptor interaction,细胞因子细胞因子受体相互作用图3 差异表达基因KEGG富集分析Fig.3 KEGG enrichment analysis of differentially expressed genes
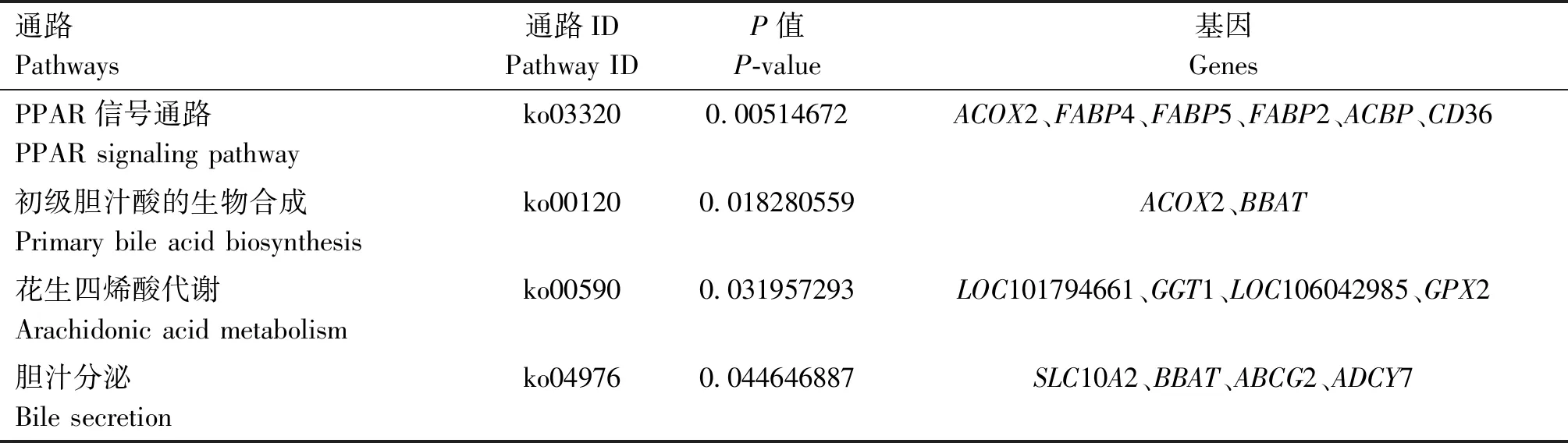
表5 与脂肪代谢相关的KEGG通路
2.5 实时荧光定量PCR 验证
为了验证RNA-Seq结果的准确性,共挑选了5个差异表达基因(ACBP、FABP2、FABP4、FABP5和ACOX2基因)进行实时荧光定量PCR分析。如图4所示,H组这5个基因相对表达量均高于L组,但差异均不显著(P>0.05),与RNA-Seq结果一致。
2.6 相关性分析
通过对番鸭腹脂重和腹脂率与筛选出的基因进行皮尔森相关性分析发现,腹脂重和腹脂率与NR5A2、ACBP、ACOX2、FABP2、FABP4和FABP5基因均呈正相关(r=0.47~0.87)。其中,ACOX2和FABP5基因与腹脂重和腹脂率呈显著正相关(P<0.05),ACBP基因与腹脂重和腹脂率呈极显著正相关(P<0.01)(图5)。

图4 实时荧光定量PCR验证Fig.4 Validation of Real-time PCR

*,显著相关(P<0.05);**,极显著相关(P<0.01)*,Significant correlation (P<0.05);**,Extremely significant correlation (P<0.01)图5 腹脂重和腹脂率与差异表达基因的相关性分析Fig.5 The correlation analysis of abdominal fat weight and abdominal fat rate with differentially expressed genes
3 讨 论
家禽胴体脂肪含量过高会对其生产性能造成一定的影响,特别是肉用型家禽[16]。与肌内脂肪不同,腹脂不能增加肉的风味和口感,过高的腹脂含量只会造成饲料的浪费[17],因此如何调控番鸭的腹脂含量成为了一个亟待解决的问题。腹脂的形成与脂肪的代谢和沉积有很大的关系,可以通过调控脂肪的代谢和沉积来影响腹脂的生成。脂肪是体内重要的能量来源和能量贮备形式,在体内的能量动态平衡中发挥着重要作用[18]。家禽脂肪细胞的自身合成能力有限,其脂肪来源主要是肝脏的合成以及肠道外源脂肪的吸收[19]。动物采食后,其中的脂类分子会被肠道吸收,经门脉系统被运送至血液,储存于脂肪组织中(如腹脂)[20]。因此,本试验对番鸭的回肠组织进行转录组分析,共筛选出602个差异表达基因。通过GO功能和KEGG富集分析,筛选出与脂肪代谢相关的5个GO通路、4个KEGG通路和6个基因(NR5A2、ACBP、ACOX2、FABP2、FABP4和FABP5基因)。
GO富集分析发现,脂质应答、类固醇激素介导的信号通路、类固醇激素应答、类固醇激素刺激的细胞应答和脂质的细胞应答5个条目与脂肪代谢相关,其中5个条目富集的基因是相同的,只有NR5A2基因参与了脂肪的代谢,其他基因与脂肪代谢之间的关系尚不明确。核受体通过调节参与胆汁酸合成、胆固醇稳态和甘油三酯合成的基因的表达,是一个关键的代谢传感器[21]。与氧化甾醇受体NR1H3/LXR-alpha和NR1H2/LXR-beta一起,是脂质代谢的重要转录调节因子[22]。
KEGG富集分析发现,有4个与脂肪代谢相关的通路:PPAR信号通路、初级胆汁酸的生物合成、花生四烯酸代谢和胆汁分泌。PPAR信号通路主要参与调控脂肪酸代谢、糖代谢、细胞增殖与分化等[23]。FABP2、FABP4、FABP5、ACBP和ACOX2等与脂肪代谢相关的基因在此通路富集表达。FABP4是脂肪酸结合蛋白(fatty acid-binding proteins,FABPs)家族中的一员,对机体的脂质代谢有重要的调控作用,一方面它能促进血液内的脂肪酸向细胞膜流动;另一方面又能通过结合脂肪酸来调控脂类代谢相关基因的表达[24]。该基因在H组得到上调,说明其在H组中得到较高的表达。同属于脂肪酸结合蛋白家族的FABP2和FABP5也在H组上调表达。其中FABP2主要存在于小肠细胞中,但在回肠中表达最高,参与了从肠腔中吸收脂肪酸的过程,负责长链脂肪酸的吸收、运输与代谢[25-26]。研究表明,FABP5通过影响脂质代谢从而发挥信号转导作用,广泛参与摄取和运输长链脂肪酸,基因调控、细胞生长和分化[27]。研究表明,ACBP可以刺激食欲,与人的肥胖也息息相关[28]。此外,研究结果表明,胞质外的ACBP在调节摄食行为以及葡萄糖和脂质代谢过程中起关键作用[29]。在本试验中发现该基因在H组的表达高于L组,与上述研究结果一致。ACOX2基因在H组上调表达,其参与脂肪酸代谢、脂蛋白代谢和胆汁酸合成以及过氧化物脂类代谢,在脂肪酸代谢途径中高表达,是脂肪酸代谢过程中重要的调控基因[30]。ACOX2基因又被称作过氧化物酶支链酰辅酶A氧化酶2,在过氧化物酶体中,该基因作为一种限速酶,在支链脂肪酸β氧化过程中催化不饱和脂肪酸氧化,将长链脂肪酸氧化生成短链脂肪酸[31]。此外,该基因还会催化胆固醇形成胆汁酸,而后者可以通过影响葡萄糖代谢以及脂质和能量的消耗,从而调控组织内脂肪酸的含量[32]。
4 结 论
本试验通过对高、低腹脂率番鸭回肠进行转录组测序分析,发现共有602个差异表达基因,其中包括285个上调基因和317个下调基因。差异基因主要富集于5个与脂肪代谢相关的GO通路(脂质应答、类固醇激素介导的信号通路、类固醇激素应答、类固醇激素刺激的细胞应答和脂质的细胞应答)和4个KEGG通路(PPAR信号通路、初级胆汁酸的生物合成、花生四烯酸代谢和胆汁分泌)。此外,筛选出了6个与脂肪代谢显著相关的基因:NR5A2、ACBP、ACOX2、FABP2、FABP4和FABP5,其中,ACOX2和FABP5与腹脂重和腹脂率呈显著正相关,ACBP与腹脂重和腹脂率呈极显著正相关。此研究填补了番鸭回肠组织中调控脂肪代谢沉积基因的空白,为后续腹脂沉积的调控提供了理论基础。
