间歇性θ节律经颅磁刺激改善大鼠工作记忆的海马与前额叶跨脑区神经网络效应研究*
2022-08-20张天恒郭苗苗徐桂芝吉利辉王中豪
张天恒 郭苗苗 徐桂芝 吉利辉 王中豪
(河北工业大学电气工程学院,省部共建电工装备可靠性与智能化国家重点实验室,天津市生物电工与智能健康重点实验室,天津 300130)
经颅磁刺激(transcranial magnetic stimulation,TMS)是一种非侵入、非药物治疗的神经调控技术,在探索大脑认知功能和治疗脑疾病方面得到广泛的应用,具有重要的临床价值[1-3]。θ节律刺激(theta burst stimulation,TBS)是一种模拟神经节律放电特性的新型TMS刺激模式,与传统的重复TMS相比,刺激强度降低,刺激时长缩短,被认为是更有效的刺激模式,并随后被用于进行标准化的频率刺激[4-5]。TBS按照间隔时间不同分为间歇性TBS(intermittent TBS,iTBS)和连续性TBS(continuous TBS,cTBS)。研究表明,iTBS可以对大脑皮层产生兴奋效果[6],改善大脑认知功能,但其电生理机制仍不清楚。Blumberger等[7]研究了iTBS和rTMS两种刺激模式在治疗抑郁症方面的应用,结果表明iTBS的治疗效果与10 Hz rTMS的治疗效果相当,但iTBS的治疗周期更短。
工作记忆(working memory,WM)是中枢神经系统重要的高级功能,在许多复杂任务的高级认知活动中发挥着重要作用,如理解、阅读及学习等[8]。WM障碍是神经系统正常衰老和一些神经退行性疾病,如轻度认知障碍、阿尔茨海默病(Alzheimer’s disease,AD)等的主要临床表现之一,因此研究WM的神经机制对WM障碍等神经系统疾病的治疗具有重要的科学价值[9]。工作记忆是较高认知能力的基础,腹侧海马(ventral hippocampus,vHPC)和内侧前额叶皮层(medial prefrontal cortex,mPFC)脑区是工作记忆的主要责任脑区[10]。近年来已有研究证明,TMS作用于大脑多靶区可以有效改善AD患者的认知功能和记忆[11],特别是轻度障碍患者和AD早期。王欣[12]通过对AD转基因鼠的实验研究发现,TMS可以实现对AD早期异常神经活动的调控,并实现了对AD认知障碍的延缓。同时,iTBS的刺激方案也被广泛用于研究AD患者记忆障碍皮质可塑性的调节机制[13],iTBS可以通过诱导突触可塑性来影响脑内的神经活动,同时神经元的突触可塑性与学习记忆密切相关[14],这种新的刺激模式也为探索大脑认知功能的神经调控提供新的有效干预方法和手段。
大脑是一个非常复杂的系统,多个神经元、神经元集群或者多个脑区相互连接成庞杂的结构网络,并通过相互作用完成脑的各种功能[15]。在体多通道记录技术能够长期稳定记录动物清醒状态下大脑多个脑区的神经元电活动信号,为研究大脑特定目标脑区的神经活动提供有效技术手段[16]。大脑结构和功能的神经网络连接与脑认知机制之间存在紧密联系[17],学习记忆等认知功能的实现需要大脑多个脑区的神经同步活动,神经网络研究已经成为脑功能与脑疾病研究领域的热点[18]。有研究表明,在空间工作记忆任务期间,vHPC-mPFC网络中双向信息流增加,认为信息流的增加预测了记忆的准确性[19]。同时有研究发现,记忆和认知能力下降与大脑不同脑区之间联系的改变有关,这可能是由相应脑区之间结构和功能上的连接障碍所致[20-21]。工作记忆障碍被认为是大脑认知功能衰退的核心特征,它来源于局部脑回路与远程脑回路间的失联[22],Dai等[23]基于磁共振成像和图论网络分析,发现AD患者的临床表现与大脑功能连接网络或结构连接的异常有关。因此,从前额叶-海马神经网络协同性的角度探索经颅磁刺激对工作记忆的调控机制研究具有十分重要的意义和价值[24]。
因此,本文应用工作记忆行为学和电生理学,通过在体多通道微电极记录技术,记录并获取工作记忆过程中大鼠mPFC和vHPC的局部场电位(local field potentials,LFPs)信号,基于时域格兰杰因果方法,深入分析了mPFC-vHPC双脑区的神经网络连接,从神经网络协同性的角度,研究了iTBS磁刺激模式对工作记忆过程中跨脑区神经网络的调控作用,为临床TMS模式优化提供科学依据与理论支撑,具有重要的学术价值和应用前景。
1 材料与方法
1.1 磁刺激大鼠模型制备
本文以12只成年雄性Wistar大鼠为实验对象,随机分为刺激模型组和对照组,每组6只,大鼠周龄为8~10周,体重约300 g,本实验均已由河北工业大学生物医学伦理委员会审查通过(审查编号:HEBUTaCUC201905)。
TMS设备采用重复脉冲磁刺激系统(英国Magstim公司生产,型号:Rapid2),刺激线圈采用8字型线圈。刺激时,将大鼠的头部固定,同时避免其挣扎减少应激反应。刺激线圈放置在与大鼠的顶骨平行的头皮表面上方约2~3 mm处。刺激组采用iTBS经颅磁刺激模式对大鼠头部进行刺激,刺激强度为100%运动阈值,刺激频率为5 Hz,每天刺激600个脉冲,刺激总时长为14 d,对照组大鼠不进行任何刺激操作。经颅磁刺激的TBS模式中每个脉冲又包含3个50 Hz的高频脉冲,iTBS刺激模式为刺激2 s,间隔8 s(图1)。

Fig.1 Schematic diagram of iTBS magnetic stimulation paradigm
1.2 电极植入与数据采集
磁刺激完成后,利用在体多通道技术对大鼠进行电极植入手术,分别在大鼠vHPC和mPFC脑区各植入8通道微电极阵列(1~8通道为mPFC,9~16通道为vHPC),电极位置排布均为2×4(图2b)。
首先,在大鼠腹腔注射水合氯醛(40 mg/kg)进行麻醉,皮下注射阿托品以减少呼吸道分泌。待其麻醉后,剔除大鼠头部毛发并将其头部固定在立体定位仪上。切开头部皮肤,暴露颅骨,找到冠状缝与矢状缝的交点,为前囟点位置。参考大鼠脑定位图谱[25],以前囟点为参考原点,定位mPFC脑区(向前2.5~4.5 mm,旁开0.2~1.0 mm,深度2.5~3.0 mm)和vHPC脑区(向后4.0~5.3 mm,旁开4.0~5.0 mm,深度7.5~8.5 mm)(图2)。

Fig.2 The location of electrode placement in brain regions of rats and the LFPs signals
使用颅钻在颅骨上述定位区域各开一个矩形窗,暴露并移除硬脑膜,将双脑区微电极阵列缓慢垂直推进至目标脑区深度,同时,对记录到的神经元信号进行持续监控,最后用牙科水泥和不锈钢螺钉进行固定。手术完成后,将大鼠放回饲养笼进行单独饲养,恢复期2 d后,进行T迷宫工作记忆行为学任务,并同步采集记录行为学过程中的大鼠vHPC和mPFC脑区的LFPs信号。
本文应用在体多通道神经电生理信号采集系统(美国Plexon公司生产,OmniPlex128)对大鼠vHPC和mPFC脑区的神经电信号进行记录,采样频率为1 kHz,通过500 Hz低通滤波从神经电信号中提取原始LFPs信号,再经过去除50 Hz工频干扰和基线漂移,以及0.5~100 Hz滤波等预处理,然后用于后续的数据处理与分析。
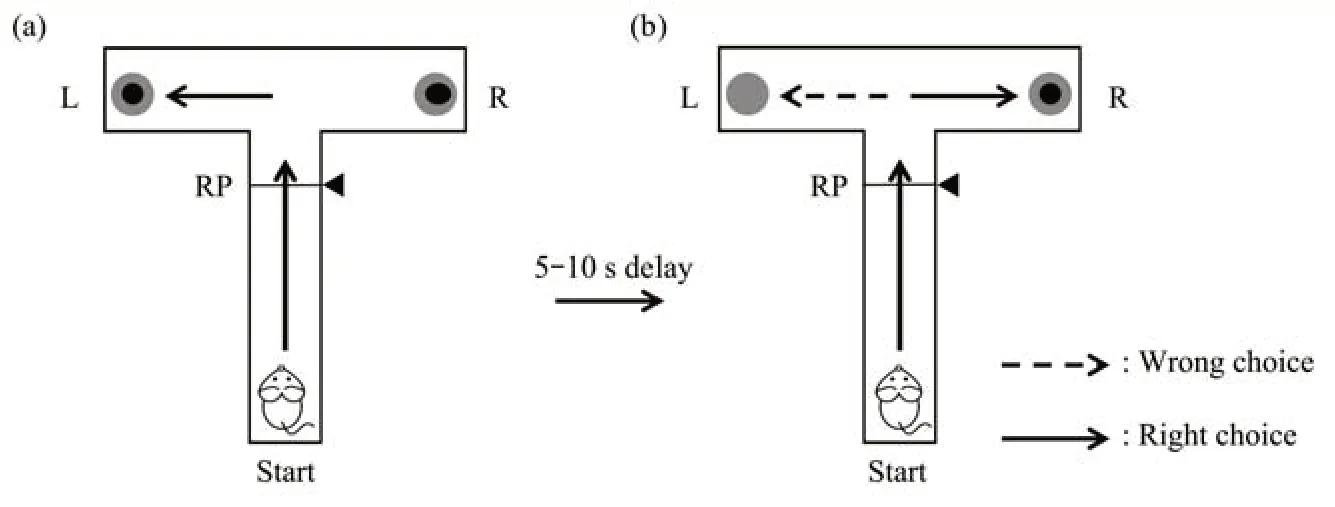
Fig.3 T Maze WMbehavioral task
1.3 T迷宫行为学实验
本文应用T迷宫[26-27]行为学实验来评价大鼠工作记忆能力。大鼠恢复期过后,开始对大鼠进行2 d适应性训练,即让大鼠在迷宫中自由探索,2次/d,每次30 min,随后开始正式训练任务。每天对每只大鼠进行两组T迷宫WM任务训练,每组训练包含20次实验。每次实验任务均包括自由选择和延时交替选择两个阶段(图3)。任务开始前,在T迷宫左右两目标臂端放置食物奖励;任务开始后,大鼠从起始端出发,首先进行自由选择阶段,此时大鼠可以任意进入一个目标臂端,并获得食物奖励。随后大鼠回到起始端经过5~10 s延迟,重新出发,开始延时交替选择阶段。此时当大鼠选择与自由选择阶段不同目标臂端时,大鼠可以获得食物奖励,并视为正确执行WM任务;而当大鼠选择与自由选择阶段同一目标臂端时,则大鼠不能获得食物奖励,视为错误执行WM任务。一次实验任务完成以后,大鼠返回到起始区域,准备重新开始下一次WM任务训练。在大鼠执行T迷宫WM任务行为学过程中同步采集记录vHPC和mPFC脑区的LFPs信号。
记录每天大鼠行为学数据结果,以正确执行WM任务次数占总任务次数的百分比表示大鼠每天任务的正确率。当大鼠执行T迷宫WM任务的正确率达到并保持在80%以上连续2 d,认为其“学会”此任务。在大鼠进行T迷宫工作记忆行为学任务的同时,同步采集记录行为学过程中的大鼠vHPC和mPFC脑区的LFPs信号。将大鼠正确执行工作记忆任务中延时交替选择阶段的LFPs数据用于后续的网络计算和分析。
1.4 数据处理与分析
本文基于格兰杰因果[28-29](Granger causality,GC)网络连接模型,构建复杂因果网络,对比分析不同组大鼠在工作记忆任务过程中vHPC-mPFC跨脑区网络的因果关联特性,研究iTBS磁刺激对大鼠工作记忆的跨脑区神经网络调控机制。
格兰杰因果关系GC值的大小反映了不同电极通道之间神经信号因果连接的强度大小。计算神经信号每两个通道之间的因果连接GC值,同时采用Bonferronni校正计算有显著性(P<0.05)的格兰杰因果GC值,去除无效连接,这样可以得到一个因果连接矩阵网络GCd:

其中,GCij表示通道j对通道i的因果连接。分别计算vHPC与mPFC脑区内部以及vHPC-mPFC神经信号每两个通道之间的因果连接GC值,可以分别构建得到vHPC与mPFC脑区自身的因果网络,以及vHPC-mPFC两个脑区的跨脑区因果网络。
然后在矩阵网络GCd的基础上,对网络进行阈值化处理剔除网络伪连接,得到二值矩阵网络GCb:
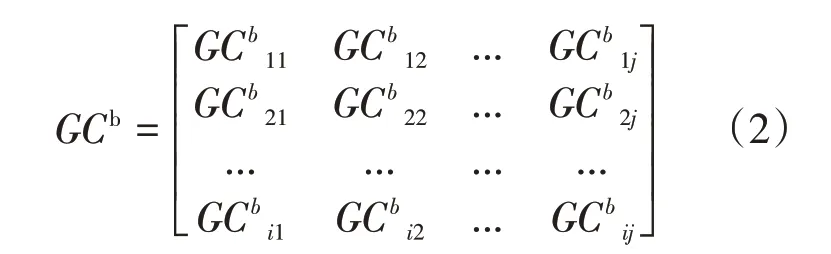
其中,GCbij的值只为0或者1,当GCbij大于阈值为1,小于阈值为0。当计算每个脑区内因果连接时,对角线为节点自连接,设置为0。
在二值矩阵网络的基础上,进一步计算网络连接强度、连接密度、全局效率以及因果流向性等网络特性进行对比分析。
网络连接密度可以表征网络通道节点之间连接数目的多少,用CD表示:
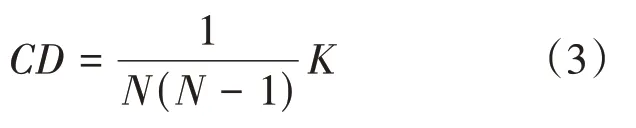
其中,N表示通道数量,K表示矩阵网络中所有非零元素的数量。
全局效率指的是网络中各节点之间最短路径长度的调和平均数的倒数,最短路径长度表示网络中从一个节点到另一个节点的最短连接边的长度。全局效率是衡量网络对信息的总体传输能力的全局指标,用Eg表示:

其中,dij表示从节点j到节点i的最短路径,auv表示从节点j到节点i的连接边,Lij表示从节点j到节点i的最短路径长度。
节点的度定义为网络中与节点之间相连的其他节点数目,一个节点的度越大意味着这个节点在网络中越重要。在有向网络中,节点的度包括出度和入度,节点总的度大小为该节点出度与入度的和:
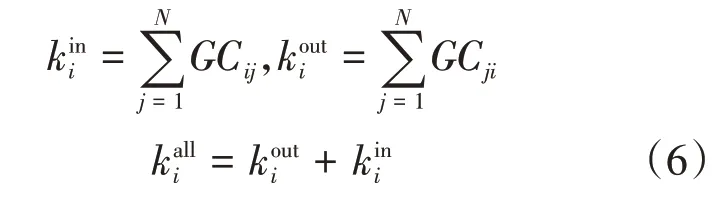
其中,kiin为入度,表示其他通道对通道i有因果连接的通道数;kiout为出度,表示通道i对其他通道有因果连接的通道数。
因果流向性是复杂网络连接中的一个重要性质。网络节点的因果流向定义为其出度和入度之间的差值,用CFi表示:
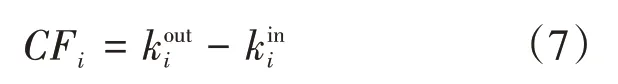
其中,CFi值大于0,表示该节点对整个网络产生强烈的因果影响,为因果源,反之CFi值小于0,表示该节点为因果汇。
1.5 组织学检验
在实验结束后,对大鼠脑组织进行切片,验证微电极阵列在大鼠脑区的植入位置。将大鼠全脑完整地取出,放入多聚甲醛溶液中进行浸泡固定24 h,再将全脑放入30%的蔗糖溶液中进行脱水,等待大脑沉降24 h后取出,分别进行前额叶和海马脑区的修块保留,然后使用振动切片机,沿电极植入方向进行大脑冠状切片,脑片厚度为150μm,将切好的脑片按照顺序贴在载玻片上,并用中性树胶进行封片,最后应用光学显微镜拍照记录实际电极植入位置,并与大鼠脑图谱进行比对,验证植入位置是否在海马脑区和前额叶皮层。
1.6 统计学分析
本实验所得数据采用Wilcoxon符号秩和检验法进行统计学分析,以P值大小为参考对不同组间数据差异是否有统计学意义进行判断,认为当P<0.05时数据具有显著性差异并记为“*”,同时P<0.01时记为“**”,P<0.001时记为“***”。
2 结 果
2.1 组织学验证结果
对大鼠脑组织进行切片检测,记录大鼠前额叶皮层和海马脑区的切片图,并与大鼠脑图谱进行对照,结果如图4所示。
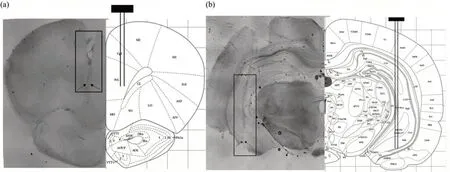
Fig.4 The histological representations of the electrode positions
图4右侧部分为标准脑图谱[25],图中PrL和CA1分别为前囟点前4.20 mm前额叶皮层和前囟点后5.04 mm海马脑区位置;左侧为实际大脑切片图,由于微电极阵列位置排布均为2×4,在脑组织切片上可以观察到两列电极孔道,图中黑色框和黑色箭头表示为两列电极孔道位置和电极末端位点。
2.2 行为学统计分析
统计每组大鼠“学会”工作记忆任务所需时长天数,作为评价大鼠工作记忆能力的行为学指标,并对刺激组和对照组大鼠平均时长天数进行对比分析(图5)。
对照组大鼠“学会”工作记忆任务的平均时长天数为(7.67±1.966)d,刺激组大鼠“学会”工作记忆任务的平均时长天数为(5.00±0.894)d,刺激组大鼠“学会”工作记忆任务所需时长天数明显低于对照组,完成工作记忆任务所需时长减少了(2.67±1.63)d,经Wilcoxon符号秩和检验,两组时长天数之间存在统计学差异(df=5,Z=-2.207b,P=0.027)(图5)。结果表明,iTBS刺激增强了大鼠的学习记忆能力,使其完成工作记忆任务所需时长减少。
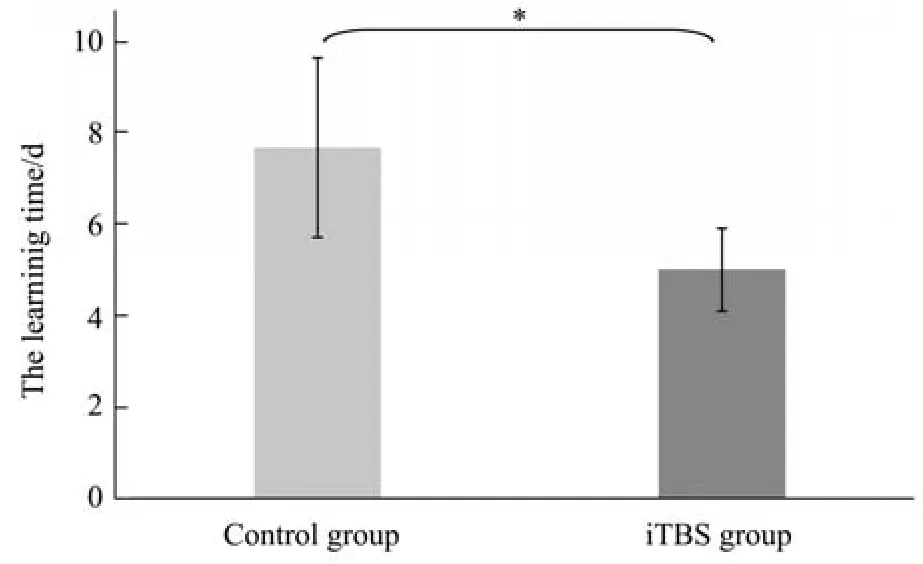
Fig.5 The behavioral results of rats in T maze(n=6,*P<0.05)
2.3 磁刺激对前额叶脑区因果连接的影响
选取大鼠在正确执行工作记忆任务中延时交替选择阶段的LFP数据,以T迷宫行为学参考点为0点,前后各取了2 000 ms,总共4 000 ms的数据长度进行计算,利用格兰杰因果网络构建方法,构建分析了对照组和刺激组大鼠在工作记忆过程中mPFC脑区的脑因果网络以及网络特性值(图6)。

Fig.6 The comparison of mPFC causal network between iTBS group and control group
如图6a1所示,网络矩阵的横坐标与纵坐标均表示mPFC植入的电极通道,矩阵中的每个元素对应从横轴对应通道到纵轴对应通道的格兰杰因果连接强度,色条表示因果连接强度的大小,颜色越接近红色表示对应两个通道之间的因果连接越大;反之,颜色越接近蓝色表示对应的因果连接越小,对角线表示通道的自因果,均设置为0。同时,在mPFC脑区的有向网络连接中(图6a2),单向连接用黑色线表示,双向连接用紫色线表示。可以看出,在大鼠的mPFC脑区因果网络中,iTBS组大鼠mPFC脑区因果网络连接强度明显强于对照组,网络双向连接数量多于对照组,表明iTBS刺激增强了工作记忆过程中mPFC脑区内的因果连接性。
进一步对网络连接强度、连接密度以及全局效率等网络特性进行对比分析(图6b),可以看出,iTBS组大鼠mPFC脑区因果网络的连接强度、连接密度以及全局效率值都明显高于对照组大鼠。经Wilcoxon符号秩和检验,两组之间均存在统计学差异(连接强度:df=119,Z=-4.460b,P=8.00×10-6;连接密度:df=119,Z=-4.621b,P=4.00×10-6;全局效率:df=119,Z=-3.150b,P=0.002),3种网络特性值变化趋势基本一致,表明iTBS刺激增强了工作记忆过程中mPFC脑区内的因果连接强度,使连接数量增多,连接密度增大,并进一步提高了信息传递效率。
2.4 磁刺激对海马脑区因果连接的影响
按照同样的方法,构建分析了对照组和刺激组大鼠在工作记忆过程中vHPC脑区的脑因果网络以及网络特征参量(图7)。
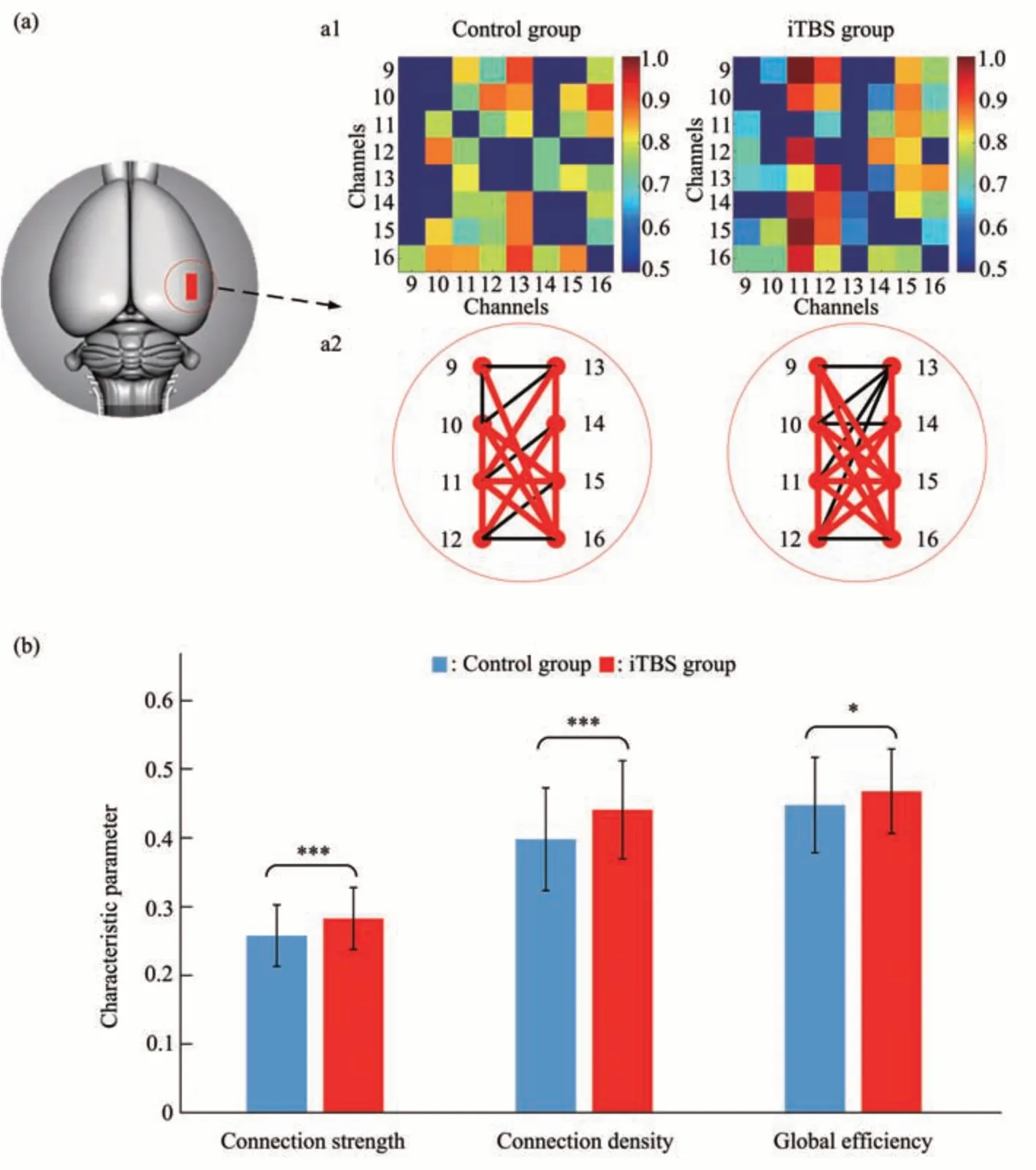
Fig.7 The comparison of vHPC causal network between iTBS group and control group
如图7a1所示,矩阵的横坐标与纵坐标均表示vHPC植入的电极通道,矩阵中的每个元素对应从横轴对应通道到纵轴对应通道的格兰杰因果连接强度。同时,在vHPC脑区的有向网络连接中(图7a2),单向连接用黑色线表示,双向连接用红色线表示。可以看出,在大鼠的vHPC脑区因果网络中,iTBS组大鼠vHPC脑区因果网络连接强度明显强于对照组,网络双向连接数量多于对照组,表明iTBS刺激增强了工作记忆过程中vHPC脑区内的因果连接性。
同样进一步对vHPC脑区网络连接强度、连接密度以及全局效率等网络特性进行对比分析(图7b),可以看出,iTBS组大鼠vHPC脑区因果网络的连接强度、连接密度以及全局效率值都明显高于对照组大鼠。经Wilcoxon符号秩和检验,两组之间均存在统计学差异(连接强度:df=119,Z=-3.740b,P=1.84×10-4;连 接 密 度:df=119,Z=-4.139b,P=3.50×10-5;全 局 效 率:df=119,Z=-2.006b,P=0.045),3种网络特性值变化趋势基本一致,表明iTBS刺激增强了工作记忆过程中mPFC脑区内的因果连接强度,使连接数量增多,连接密度增大,并进一步提高了vHPC脑区内的信息传递效率,此结果与上述mPFC脑区因果网络结果变化较为一致。
2.5 磁刺激对前额叶与海马跨脑区因果网络连接的影响
按照同样的方法,进一步构建分析了对照组和刺激组大鼠在工作记忆过程中vHPC与mPFC脑区的双向的跨脑区因果网络(图8)。
如图8a1和8b1所示,网络矩阵的横坐标与纵坐标分别表示mPFC和vHPC植入的电极通道,矩阵中的每个元素对应从横轴对应通道到纵轴对应通道的格兰杰因果连接强度。同时,在mPFC与vHPC的有向网络连接中(图8a2,b2),单向连接分别用蓝色线和红色线表示。可以看出,在大鼠vHPC与mPFC的跨脑区网络中,vHPC-mPFC网络矩阵的因果连接数量更多,连接强度更大,同时与对照组相比,iTBS组的vHPC-mPFC和mPFCvHPC网络矩阵的因果连接数量和连接强度都有所增大。经Wilcoxon符号秩和检验,刺激组与对照组之间均存在统计学差异,vHPC-mPFC网络(连接强度:df=119,Z=-2.079b,P=0.038;连接密度:df=119,Z=-2.263b,P=0.024);mPFC-vHPC网络(连接强度:df=119,Z=-3.198b,P=0.001;连接密度:df=119,Z=-3.312b,P=0.001)。结果表明,在工作记忆过程中,从vHPC到mPFC有更多的信息传递,并且iTBS刺激增强了工作记忆过程中vHPC-mPFC的因果连接性,从vHPC到mPFC的因果连接更加明显,提高了vHPC-mPFC两个脑区的信息交互。
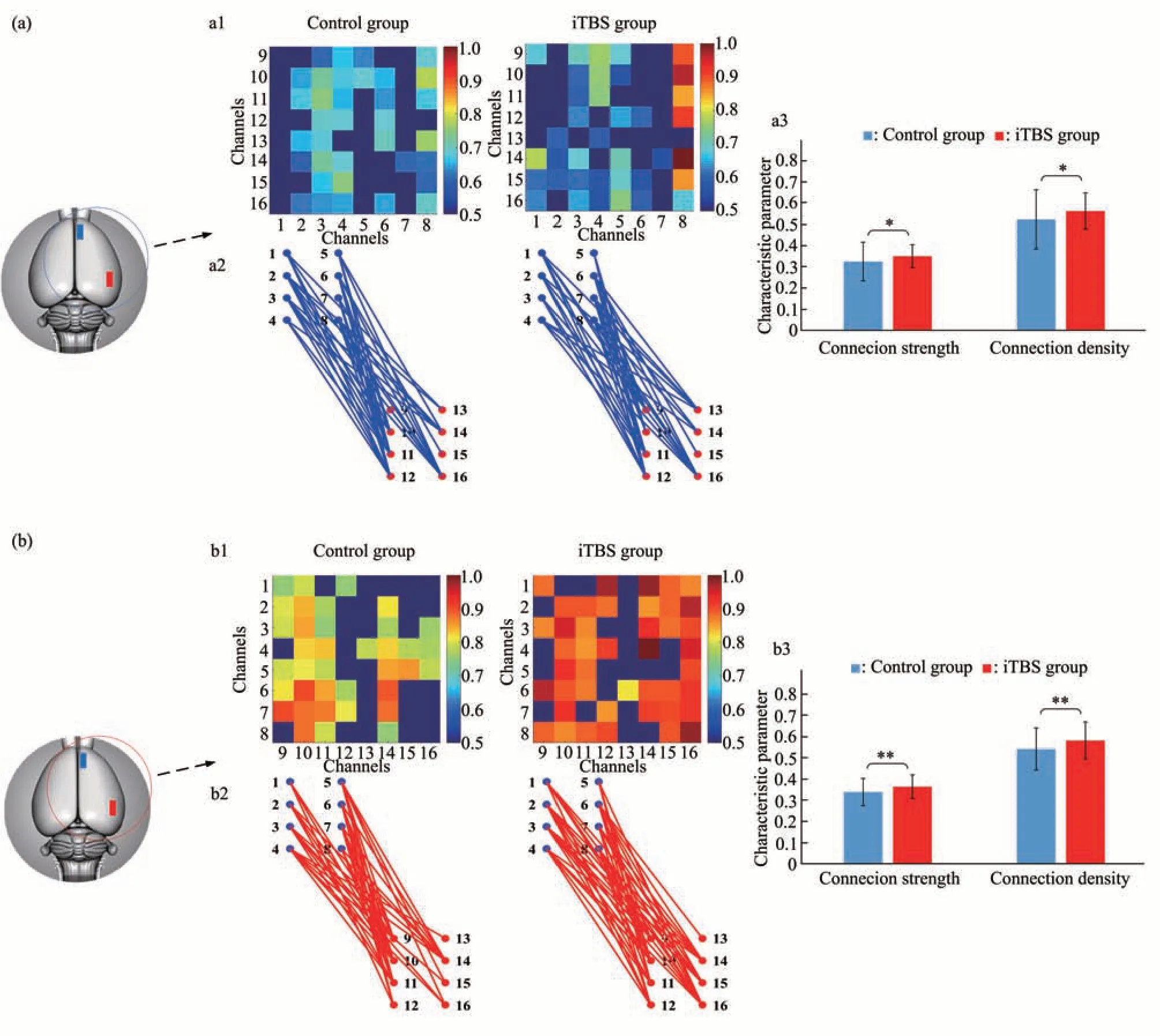
Fig.8 The comparison of vHPC and mPFC cross-brain network between iTBS group and control group
2.6 磁刺激对前额叶与海马跨脑区因果网络连接特性的影响
在vHPC与mPFC的跨脑区因果网络基础上,进一步分析了对照组和刺激组大鼠在工作记忆过程中vHPC与mPFC脑区的度连接分布和信息流向(图9)。
图9a为大鼠vHPC与mPFC的跨脑区网络度分布,色条表示度连接的大小,颜色越接近红色表示对应通道的度连接越大;反之,颜色越接近蓝色表示对应通道的度连接越小。可以看出,vHPC脑区比mPFC脑区表现出更大的度连接,最大度连接的节点通道位于vHPC脑区,说明在vHPC与mPFC的跨脑区网络中,vHPC脑区里有更重要的网络连接节点,vHPC脑区为主要责任脑区,在工作记忆过程中负责更多的信息整合。同时,iTBS组大鼠mPFC与vHPC脑区的整体度连接分布呈现增加趋势(图9b),并且平均度连接值显著增大(df=119,Z=-2.113b,P=0.035),说明iTBS增强了vHPC与mPFC的跨脑区网络连接,提高了vHPC与mPFC两个脑区的信息交互。

Fig.9 The comparison of vHPC and mPFC network node degrees between iTBS group and control group
图10a为大鼠vHPC与mPFC的跨脑区网络因果流向,色条表示因果流向的大小,颜色越接近红色表示对应通道的因果流向越大,正值呈现因果流出;反之,颜色越接近蓝色表示对应通道的因果流向越小,负值为因果流入。从图10a,b可以看出,vHPC主要表现为因果流出,mPFC主要表现为因果流入,iTBS组大鼠的mPFC与vHPC脑区的整体因果流向明显增加,并且平均因果流向值显著增大(df=119,Z=-3.923b,P=8.8×10-5),说明大鼠在工作记忆过程中从vHPC到mPFC有更多的信息传递,并且iTBS增强了从vHPC到mPFC的因果流向性,提高了vHPC-mPFC两个脑区的信息传递和交流。

Fig.10 The comparison of vHPC and mPFC network causal flow between iTBS group and control group
3 讨 论
本文基于格兰杰因果关系算法,对大鼠执行WM任务中的LFPs信号因果网络进行分析,并引入网络连接强度、网络连接密度、全局效率以及因果流向等网络特性参数,通过对比分析对照组和刺激组在T迷宫内执行WM任务过程中的行为学表现及LFPs信号因果网络特性的差异,讨论分析了iTBS对大鼠WM相关行为及神经网络关联特性的影响。
行为学结果表明,经过iTBS刺激的正常成年大鼠比无磁刺激的正常成年大鼠达到“学会”记忆任务的平均时长天数显著缩短(P<0.05),经颅磁刺激对大鼠的认知行为产生了相应的影响,提高了大鼠工作记忆能力。行为表现是大脑神经活动的最终表达,也是反映磁刺激效应的宏观表现。已有研究表明,iTBS能够提高健康人类对照组的认知能力[30],在本文结果中,iTBS使大鼠在更短的时间内完成了工作记忆任务,提高了大鼠认知记忆能力,与以往的对iTBS刺激积极影响的研究结果较为一致,并进一步表明了iTBS在提高工作记忆等认知功能方面的有效性。
前额叶和海马脑区是工作记忆的主要责任脑区,其中vHPC在记忆的形成和巩固中发挥重要作用,mPFC也与记忆的巩固和提取尤其相关,工作记忆的功能实现跟mPFC与vHPC脑区各自发挥作用都息息相关,磁刺激对mPFC与vHPC脑区神经网络的影响也会因此影响到工作记忆过程中。越来越多的研究表明,mPFC与vHPC脑区的神经同步活动是工作记忆的重要机制[31-32]。本文利用格兰杰因果方法,研究了iTBS对mPFC和vHPC脑区的神经网络影响,结果表明,iTBS增强了mPFC和vHPC脑区内神经网络连接,提高了网络连接密度和传递效率,提高了mPFC与vHPC脑区的神经同步活动,并且mPFC和vHPC脑区的增强效果具有一致性。同时mPFC和vHPC在生理结构上存在直接通路的神经投射连接,主要起自vHPC脑区到mPFC脑区[33],vHPC-mPFC回路是海马和前额叶之间存在功能联系的解剖学基础,磁刺激对mPFC及vHPC的增强一致性,也证实了两个脑区结构和功能上的网络连接信息交互。
由于记忆的复杂性,其功能无法由单独脑区独立完成,工作记忆的形成依赖于大脑不同脑区结构或功能上的网络连接,并且前额叶皮层与海马脑区之间的网络连接是工作记忆功能执行的关键[34],前额叶和海马通过分工合作共同完成认知活动任务[35-36]。以往的研究表明,与工作记忆相关的神经活动主要从海马转移到前额叶,工作记忆相关的大脑处理信息随神经活动在脑区之间发生传递[19]。本文的研究结果进一步证实了以往的研究,在本文中,从vHPC到mPFC的因果连接网络更强,连接数量更多,并且网络因果流向也表现为vHPC流出、mPFC流入。本文iTBS的作用结果表明,iTBS增强了从vHPC到mPFC的网络连接、节点度和因果流向,使更多的大脑信息从vHPC流向mPFC,增强了vHPC与mPFC之间的协同交互。而且vHPC与mPFC脑区相关神经元的信息功能影响了大鼠工作记忆的行为表现,从而导致了大鼠工作记忆能力的提升,本文中iTBS影响神经网络的变化与行为学结果呈现一致性。因此,iTBS可以通过提高vHPC与mPFC之间神经信息的网络协同作用来提高大鼠的工作记忆行为表现。
此外有研究表明,vHPC与mPFC的theta和gamma振荡同步是工作记忆中信息传递的关键[31],同时记忆的形成、巩固与检索过程都依赖于特征神经振荡在PFC和HPC脑区间的同步作用,本文从时域网络研究中发现从vHPC到mPFC的脑网络连接可能是工作记忆的潜在编码机制之一,因此theta和gamma频段的网络同步也是今后研究iTBS调控大脑认知功能关注的重点,iTBS对于vHPC与mPFC的网络同步效应可能影响到了记忆形成或检索的不同阶段,iTBS对于工作记忆发展过程中theta和gamma频段网络同步的影响有待进一步的详细剖析,以期望更深层的挖掘出iTBS的神经调控机制。
4 结 论
本文结果表明,iTBS模式经颅磁刺激对大脑神经活动及行为表现均有显著的积极影响,iTBS磁刺激可以促进大鼠在工作记忆任务的行为表现,并且能够增强大鼠vHPC-mPFC脑区的网络连接,提高信息交互和传递效率,改善大鼠工作记忆能力。iTBS的神经调控机制可能是通过增强vHPC与mPFC之间的网络连接来提高工作记忆能力。本文有助于对iTBS刺激神经调控机制的理解,在iTBS刺激对工作记忆障碍表征的神经疾病治疗和临床应用方面具有重要意义。
