低钾胁迫对谷子幼苗叶片光合作用的影响
2022-08-19李艳芬郑君岗尹美强原向阳温银元
李艳芬,郑君岗,尹美强,原向阳,温银元
(山西农业大学 农学院,山西太谷 030801)
谷子(SetariaitalicaL.)生育期短,基因组小而简单,具有高质量的参考基因组和丰富的标记,有C4光合作用、抗旱、耐贫瘠等特征,是C4光合作用的模式作物。钾在作物生长发育过程中,既可作为营养物质,也可作为一种信号物质,调节作物的多种代谢过程[1-2],在渗透调节、叶绿素合成、优化光合性能、酶的活化等方面发挥着极其重要的作用[3-4]。中国75%的土壤缺乏有效钾[5],缺钾可导致谷子减产8.39%~33.43%[6]。
光合作用是植物生长发育和产量形成的生理基础,钾素是影响叶片光合作用的重要因子。缺钾会引起作物叶缘失绿、叶绿素含量降低[7]、气孔开度降低[8]、PSⅡ反应中心破坏[9]、碳同化关键酶活性下降[10],导致光合速率下降。彭海欢等[11]研究表明缺钾会导致水稻叶片PSⅡ反应中心受损,电子传递速率降低,发生光抑制。进一步研究低钾胁迫PSⅡ损伤位点,发现净光合速率降低主要是PSⅡ反应中心数目减少、热耗散增加,供体侧放氧复合体(OEC)遭到破坏,产生光抑制[12]。这表明缺钾导致作物光合作用降低的原因可能发生在光反应阶段(原初反应、电子传递与光合磷酸化),也可能发生在暗反应(碳同化)阶段,表现为气孔限制和非气孔限制。低钾胁迫引起谷子光合速率降低的限制因子及相关机理目前仍不清楚。
光合色素、叶绿素a荧光参数、P700氧化还原状态、气体交换参数、碳同化酶活性等指标可综合反映植物光合代谢活力及其对环境的响应特征。本研究以不同钾敏感型谷子品种为研究对象,系统分析低钾条件下谷子幼苗的钾含量、光合色素、叶绿素荧光参数、P700氧化还原状态、气体交换参数、碳同化酶活性等的变化,探讨低钾胁迫降低谷子光合作用的限制因子及相关机制,为谷子钾高效品种的选育及钾肥的合理应用提供理论支持。
1 材料和方法
1.1 试验材料
参试谷子(SetariaitalicaL.)品种2个,即钾敏感型品种‘晋谷21号’(JG21)和钾非敏感型品种‘龙谷25号’(LG25)[13]。
1.2 试验设计

1.3 测定项目与方法
1.3.1 农艺性状每个处理选取长势一致的谷子幼苗3株,测定株高、茎粗、干物重和倒二叶叶长、叶宽,并计算叶面积。
1.3.2 钾含量样品采用H2SO4-H2O2法进行消煮,原子吸收仪测定谷子幼苗叶片钾含量,数值为3次测定的平均值。
1.3.3 光合色素含量取新鲜叶片剪碎混匀,称取0.2 g,采用80%丙酮浸提法获得浸提液,于663、646和470 nm处测定浸提液吸光值。同时称取1 g叶片烘干后称重,每个处理重复3次,计算光合色素含量,单位为mg·g-1。
1.3.4 气体交换参数每个处理选择长势一致的谷子幼苗3株,利用便携式光合测定系统Li-6800测定植株完全展开叶(倒二叶)的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci)。测定前用1 100 mol·m-2·s-1饱和光强对叶片进行15 min光诱导,设定CO2浓度为400 mol·mol-1,叶室温度为25 ℃,气体流速500 mol·s-1。光合参数测定结束后标记各叶片,继续用于叶绿素荧光参数测定。
1.3.5 叶绿素荧光及P700氧化还原状态测定用便携式光纤型双通道DUAL-PAM-100荧光测量系统同步测定光系统Ⅱ(PSⅡ)叶绿素荧光参数和光系统Ⅰ(PSⅠ)P700氧化还原状态。将上述标记的幼苗叶片暗适应30 min后,打开检测光先测定初始荧光产量(F0)、最大荧光产量(Fm)和最大P700信号(Pm),随后打开光化光(AL),光强度为425 mol·m-2·s-1,待荧光信号达稳态后(4~5 min)打开饱和脉冲光(SP),信号稳定后,进行P700氧化还原态的测定,然后照射饱和脉冲光测定光适应状态下的最大荧光产量(Fm′)和PSⅠ最大荧光产量(Pm′)。PSⅡ最大光化学量子效率(Fv/Fm)、PSⅡ实际光化学量子效率(ΦPSⅡ)、PSⅡ相对电子传递速率(rETRⅡ)、光化学猝灭(qP)、非光化学猝灭(NPQ);PSⅠ光化学量子产量[Y(Ⅰ)]、PSⅠ相对电子传递速率(rETRⅠ)、PS Ⅰ供体侧限制[Y(ND)]、PSⅠ受体侧限制[Y(NA)]均由仪器计算给出,每个参数数值均为3次测定的平均值。
1.3.6 碳同化关键酶活性取新鲜的谷子倒二叶叶片0.1 g,采用二磷酸核酮糖羧化酶/加氧酶(Rubisco)和磷酸烯醇式丙酮酸化酶(PEPC)活性检测试剂盒(北京Solarbio公司)进行碳同化关键酶活性测定,每个处理重复3次。
1.4 数据分析
采用Excel 2010进行数据整理和图表制作,利用SPSS 25.0进行显著性分析,Canoco 5进行冗余(RDA)分析。
2 结果与分析
2.1 低钾胁迫对谷子幼苗生长的影响
钾在植物生长发育过程中起着重要作用,低钾胁迫(K0、K0.01、K0.1和K1)抑制了谷子幼苗的生长,幼苗株高、茎粗、叶面积、干物重、钾含量降低(表1,图1);缺钾导致谷子幼苗叶片叶面积减小、失绿枯萎,在叶尖和老叶中表现得更明显(图1)。其中,在正常钾处理(K5)下,‘晋谷21号’和‘龙谷25号’地上部干重和地下部干重均达到最大,并与其余处理差异显著,此时两品种幼苗株高、茎粗、叶面积、根冠比也达到最大或者达到较高水平。与K5处理相比,无钾(K0)处理下‘晋谷21号’地上部干重、地下部干重、叶面积和根冠比分别显著降低了35.96%、44.94%、31.35%和15.20%,K0处理下‘龙谷25号’地上部干重、地下部干重、叶面积则分别显著降低了33.33%、30.40%和16.29%,其根冠比在各处理间则未达到差异显著。同时,‘晋谷21号’和‘龙谷25号’叶片钾含量在K5处理下也达到最大,并随钾浓度降低而呈不断降低趋势,且当钾浓度低于0.1 mmol·L-1时均与K5处理差异显著;钾处理浓度低于0.01 mmol·L-1时与K0处理差异不显著;完全缺钾时‘晋谷21号’和‘龙谷25号’叶片钾含量比K5处理分别显著低降低了48.14%和37.85%。
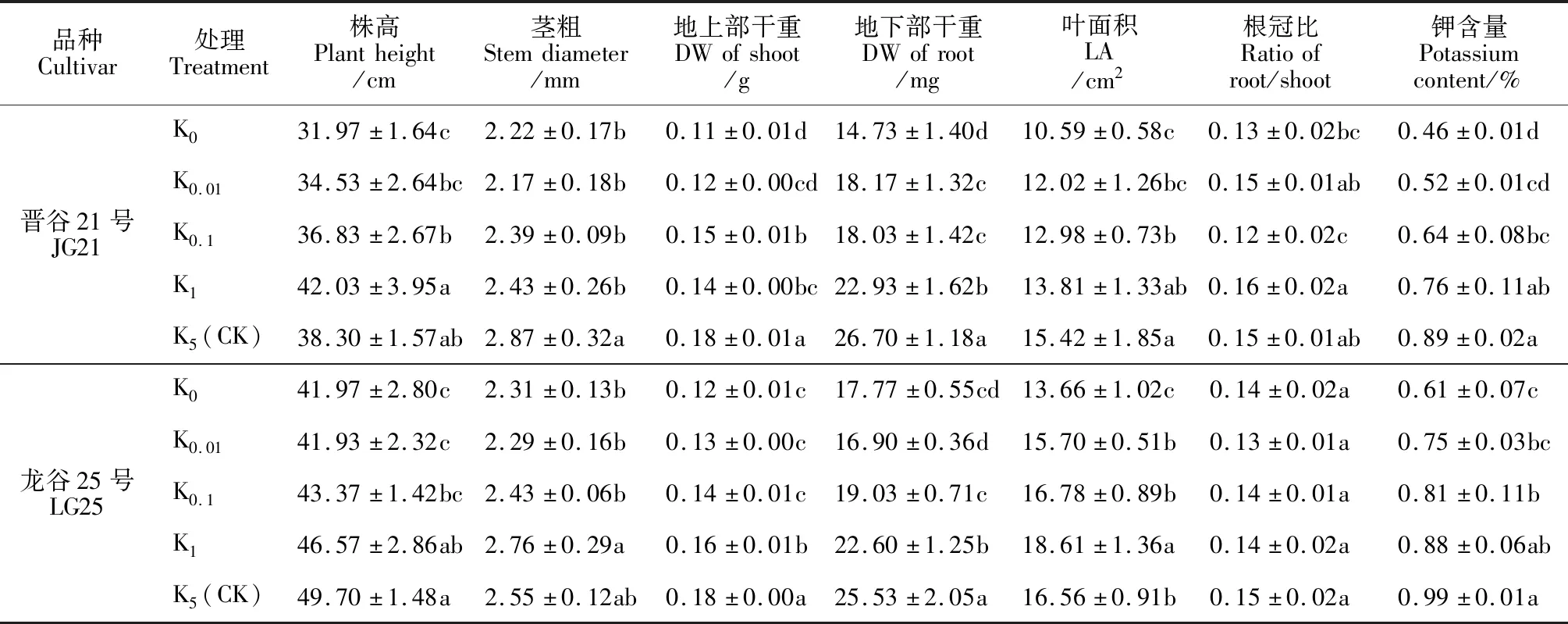
表1 不同钾浓度下谷子幼苗生长及钾含量的变化Table 1 The growth and potassium content of foxtail millet seedlings under different potassium concentrations
2.2 低钾胁迫对谷子幼苗叶片光合色素含量的影响
‘晋谷21号’和‘龙谷25号’叶片的光合色素含量与其钾含量的响应表现出相似的趋势(表2)。其中,在正常钾素处理(K5)下,‘晋谷21号’和‘龙谷25号’叶片的光合色素含量均达到最大值,总叶绿素含量分别为12.11和12.68 mg·g-1,类胡萝卜素含量分别为1.59和1.44 mg·g-1。谷子幼苗光合色素含量在缺钾处理下均比K5处理不同程度降低,但K1和K0.1处理的降低幅度均未达到显著水平;无钾处理(K0)谷子幼苗光合色素各组分含量除‘龙谷25号’叶绿素b含量外,均比K5处理显著下降,其中‘晋谷21号’以类胡萝卜素含量降幅最大(31.89%),‘龙谷25号’以叶绿素a含量降幅最大(16.44%)。

表2 低钾胁迫下谷子幼苗叶片光合色素含量的变化Table 2 The photosynthetic pigments in leaves of foxtail millet seedlings under low potassium stress
2.3 低钾胁迫对谷子幼苗气体交换参数的影响
由表3可知,低钾胁迫显著影响叶片光合气体交换参数;随着钾浓度的增加,‘晋谷21号’和‘龙谷25号’叶片净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、气孔限制值(Ls)均呈现逐渐增加趋势,而胞间二氧化碳浓度(Ci)却逐渐降低。其中,与正常钾浓度处理(K5)相比,K1处理的两品种谷子幼苗叶片各光合气体交换参数变化均不显著,K0.1处理叶片的Ci和Ls仍变化不显著,而叶片其余参数均显著降低;K0、K0.01处理两个品种谷子幼苗各光合气体交换参数与K5相比均差异显著,但K0与K0.01处理间均无显著差异。其中,与K5处理相比,K0处理下‘晋谷21号’和‘龙谷25号’叶片Pn显著降低了32.94%和21.22%,叶片Gs分别显著降低了20.06%和22.32%,其Ci却分别显著升高15.08%和27.41%。说明缺钾严重影响植物对外界CO2的吸收与同化,导致两品种叶片Pn、Gs、Tr、Ls显著降低,却增加了Ci,K0、K0.01和K0.1处理下光合速率降低主要是由非气孔限制引起的。
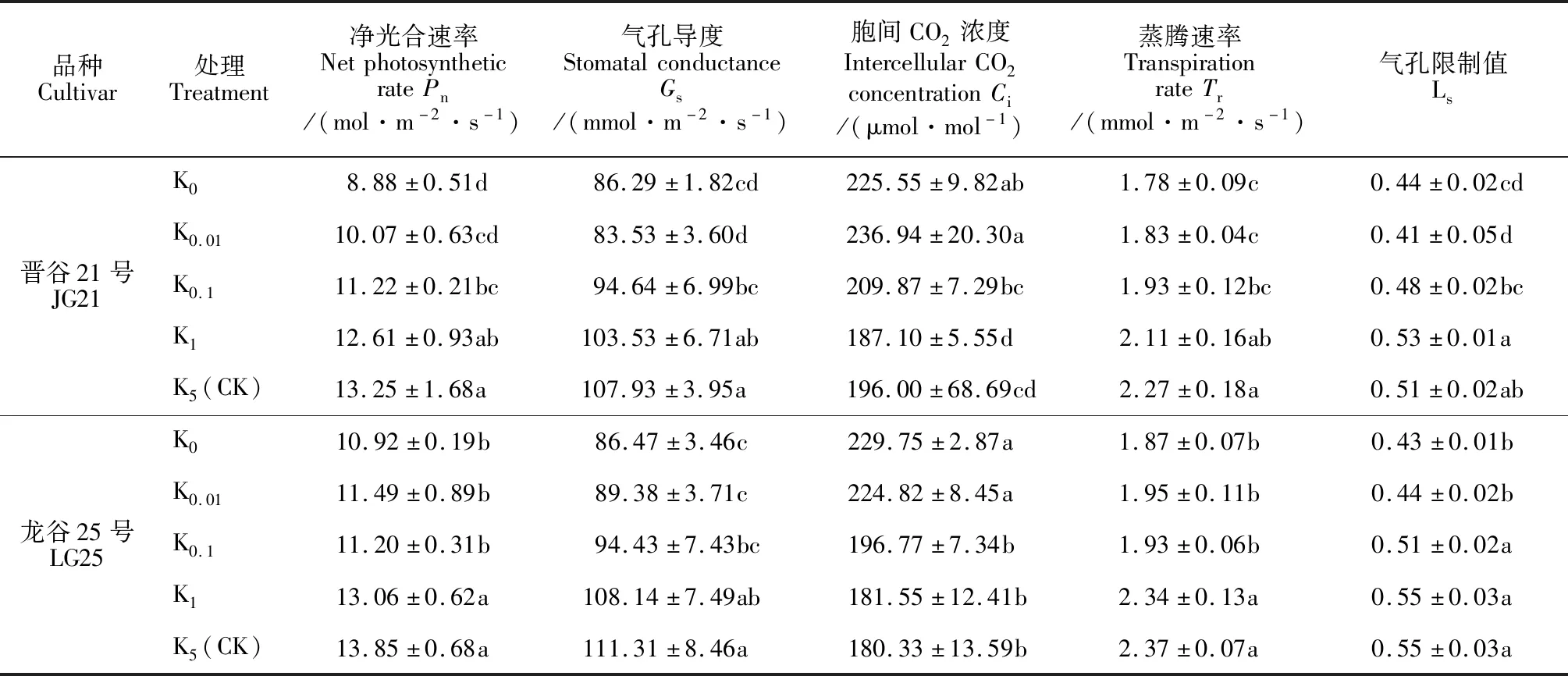
表3 低钾胁迫下谷子幼苗叶片气体交换参数的变化Table 3 The gas exchange parameters in leaves of foxtail millet seedlings under low potassium stress
2.4 低钾胁迫对谷子幼苗叶绿素荧光参数的影响
前期试验发现谷子幼苗生长状况、光合色素含量和气体交换参数在K0处理和K0.01处理下差异均不显著,故在后期试验中去掉了K0.01处理。‘晋谷21号’和‘龙谷25号’谷子幼苗叶片Fv/Fm、ΦPSⅡ和rETRⅡ在低钾胁迫下均发生了不同程度的降低,qP也基本呈降低趋势,而NPQ随钾素浓度降低基本呈升高趋势(表4)。其中,‘晋谷21号’叶片Fv/Fm、ΦPSⅡ和rETRⅡ在K1处理下就均比K5处理显著降低,‘龙谷25号’叶片Fv/Fm在K1处理、ΦPSⅡ和rETRⅡ在K0.1处理下也比K5处理显著降低,且钾浓度越降低幅度越大与受到胁迫程度呈正相关;在K0处理下,‘晋谷21号’叶片Fv/Fm、ΦPSⅡ和rETRⅡ较K5处理分别显著降低了5.19%、20.15%和20.12%,‘龙谷25号’相应显著降低了6.43%、17.15%和17.26%。同时,‘晋谷21号’叶片的qP和NPQ在各处理间差异不显著,‘龙谷25号’叶片的NPQ仅在K0处理下较K5处理显著升高了16.36%,而其相应的qP却显著降低了14.04%。以上结果说明低钾胁迫下谷子幼苗PSⅡ反应中心可能部分关闭,光合电子传递受阻,光能利用能力下降,光抑制加剧,叶片通过增加热耗散来减轻光抑制造成的损伤,且‘晋谷21号’受到的影响更大。

表4 低钾胁迫下谷子幼苗叶片叶绿素荧光参数的变化Table 4 The chlorophyll fluorescence parameters in leaves of foxtail millet seedlings under low potassium stress
2.5 低钾胁迫对谷子幼苗叶片P700参数的影响
由图2可知,随钾素处理浓度降低,‘晋谷21号’和‘龙谷25号’叶片Y(Ⅰ)、rETRⅠ基本呈逐渐降低的趋势,并在K0、K0.1处理下较K5处理降幅达到显著水平,在K1处理下变化不显著;‘晋谷21号’叶片Y(Ⅰ)、rETRI在K0处理下较K5处理显著降低了21.28%和21.31%,‘龙谷25号’则相应显著降低了18.71%和18.69%。同时,随钾素处理浓度降低,两品种叶片Y(ND)呈逐渐降低的趋势,Y(NA)则逐渐增加,并均在K0、K0.1处理下与K5处理差异显著。‘晋谷21号’和‘龙谷25号’ Y(ND)在K0处理下较K5处理分别显著降低了38.57%和44.46%,相应的Y(NA)则分别显著增高了36.19%和54.94%。以上结果说明低钾胁迫下Y(Ⅰ)的降低主要与PSⅠ受体侧非光化学量子产量Y(NA)的增加有关。
2.6 低钾胁迫对谷子幼苗碳同化关键酶活性的影响
图3显示,两品种谷子幼苗叶片碳同化酶Rubisco和PEPC活性均在K5处理下最高,并基本随钾素浓度降低呈逐渐降低趋势,两碳同化酶活性分别在K1和K0.1处理下降幅达到显著水平,且‘龙谷25号’碳同化酶活性降幅小于‘晋谷21号’降幅。其中,‘晋谷21号’和‘龙谷25号’在K0处理下的Rubisco活性分别较K5处理显著降低了42.86%和26.85%,它们相应的PEPC活性则分别较K5处理显著降低42.71%和42.77%,而两酶活性在K0与K0.1处理无显著差异。可见,低钾胁迫显著降低了谷子幼苗碳同化关键酶活性,且‘晋谷21号’降幅更大,Rubisco活性对低钾反应更敏感。
2.7 谷子幼苗叶片钾含量与其余生长生理指标的关系
利用冗余分析(RDA)研究在不同低钾胁迫下叶绿素含量、光合参数、叶绿素荧光参数、碳同化关键酶活性和幼苗生长相关指标(叶面积、植株干物质积累)之间相关性。结果(图4)表明,RDA的前两个轴分别共解释了两个品种总方差的85.0%(图4,A)和79.6%(图4,B),说明RDA的结果可靠。冗余分析能直观地显示谷子幼苗钾处理(空心箭头)与生理指标(实心箭头)间的关系;箭头连线的长度代表指标影响作用的大小,生理指标与钾处理间的夹角越小表示二者相关性越大[14]。其中,‘晋谷21号’叶片的LA、Pn、Rubisco、PSⅡ相关指标、叶绿素含量和幼苗生长指标与K5处理高度相关,PEPC和PSⅠ相关指标与K1处理有一定相关,这表明K5处理主要促进了光能的捕获和PSⅡ反应中心活性进而显著影响光合能力来促进植株生长发育(图4,A)。‘龙谷25号’除光能耗散指标外,绝大部分指标与K1处理和K5处理有良好的相关性,表明1.0 mmol·L-1K+(K1)即可满足‘龙谷25号’对钾素的需求(图4,B)。两个谷子品种都能通过改变叶片Y(NPQ)、Y(NO)、Y(NA)以适应低钾(K0.1)和无钾(K0)胁迫对光合作用的影响,‘龙谷25号’在K0处理下能维持较高的Rubisco活性,使其Pn明显高于‘晋谷21号’。
3 讨 论
植物大部分的干物质来自于绿色器官的光合作用[15],矿质元素与植物光合作用密切相关。钾素缺乏不仅影响叶绿体结构和功能,使光合色素含量降低,而且影响电子传递和碳同化关键酶活性,导致光合能力下降,抑制植物生长和发育[16]。
3.1 谷子幼苗叶片气体交换参数对低钾胁迫的响应
低钾胁迫下钾非敏感型大豆品种对胁迫的耐性较钾敏感型品种更强,叶片具有较高的光合色素含量、气孔导度和较低的胞间CO2浓度,能更好地捕获光能和与外界进行气体交换[17]。夏颖等[18]认为低钾胁迫抑制叶片气孔开放,引起光合速率降低,最终导致棉花生物量降低。杜琪等[12]则认为低钾胁迫下光合速率下降主要由PSⅡ光抑制等非气孔限制引起。耐低钾玉米自交系叶绿素含量稳定,Ci和Ls较低,比低钾敏感自交系的Pn高[19]。本研究结果显示‘晋谷21号’和‘龙谷25号’在K1处理下净光合速率并未显著降低。两品种K0、K0.01、K0.1处理叶片Pn、Gs、Tr、Ls均较K5处理显著降低,它们的Ci却显著增加,说明无钾和低钾胁迫下(K0、K0.01、K0.1)谷子幼苗光合速率降低主要是由非气孔限制引起的。同时,低钾胁迫下‘龙谷25号’和‘晋谷21号’叶片Pn较正常钾浓度处理分别降低了5.73%~21.20%和4.83%~32.94%,钾非敏感型品种‘龙谷25号’的降幅小于钾敏感型品种‘晋谷21号’。
3.2 谷子幼苗叶片PSⅠ和PSⅡ的电子传递能力和能量分配对低钾胁迫的响应
PSⅠ和PSⅡ协同进行光合电子传递并偶联磷酸化形成同化力(ATP和NADPH),推动光合碳同化进行。叶绿素荧光参数(PSⅡ)和P700氧化还原状态(PSⅠ)是反映植物对光能吸收、转化、传递、分配情况的灵敏探针,当发生胁迫时,反应中心会造成不同程度损伤,使光合电子传递和光合作用活力受到抑制。逆境下PSⅡ反应更为敏感,限制光合电子向PSⅠ传递,当植物体内活性氧过量时PSⅠ活性会受到明显抑制[20-21]。一般认为,Fv/Fm、ΦPSⅡ和qP下降表明PSⅡ反应中心活性降低,光能利用能力降低,PSⅡ受体侧电子传递受阻,PSⅡ激发压升高[22-23]。缺钾破坏茶树幼苗PSⅡ供体侧到PSⅠ的整个电子传递链,降低光合电子传递和光能转化效率,使PSⅡ过度还原,过剩激发能增加,活性降低[24]。油菜在缺钾胁迫下Fv/Fm、ΦPSⅡ、ETR和qP降低,NPQ显著上升[25]。在本研究中,低钾胁迫下谷子幼苗叶片PSⅡ和PSⅠ都发生了不同程度的抑制,PSⅡ受影响程度大于PSⅠ。低钾胁迫下谷子幼苗叶片Fv/Fm、ΦPSⅡ、qP和rETRⅡ显著降低,说明PSⅡ反应中心开放程度降低,使植物产生光抑制。NPQ反映植物耗散过剩光能和光保护能力,逆境下NPQ的增加表明叶片通过增加热耗散以减弱PSⅡ的光抑制,低钾胁迫使‘龙谷25号’谷子幼苗叶片NPQ显著升高,而对‘晋谷21号’无明显影响,说明耐低钾品种在低钾胁迫下可通过热耗散减轻光合机构遭受的光破坏。
当PSⅠ功能受损时Y(Ⅰ)会降低,同时Y(ND)和Y(NA)也会发生相应的变化。Y(ND)较高,表明植物接收的能量过剩,但可通过提高热耗散来维持PSⅠ活性。Y(NA)是指受体侧限制引起的PSⅠ非光化学能量耗散的量子产量。植物在逆境下具有较高的Y(ND)和较低的Y(NA),从而对PSⅠ起保护作用[26]。强闪光处理引起棉花PSⅠ的Y(Ⅰ)活性降低与受体侧受限制引起的非光化学量子产量Y(NA)增加有关[27]。在本研究中,随缺钾程度的增加,谷子幼苗叶片Y(ND)逐渐减小,Y(NA)逐渐增大,说明在缺钾条件下谷子幼苗叶片PSⅠ反应中心P700可有效接收PSⅡ传来的电子,并完全被还原,但是光合关键酶活性降低,同化力需求减少,受体侧电子传递受阻,PSⅠ反应中心P700不能被有效地氧化,PSⅠ活性Y(Ⅰ)降低。
3.3 谷子幼苗叶片碳同化关键酶活性对低钾胁迫的响应
Rubisco和PEPC是谷子等C4植物光合作用中决定碳同化速率的关键酶[28],叶肉细胞中的PEPC固定外界CO2转移到维管束鞘细胞中用于Rubisco催化的碳同化[29],二者的含量、活性与C4光合速率呈正相关。钾素缺乏环境下水稻Rubisco[30]和PEPC酶[10]活性下降。王晓磊等[19]发现低钾胁迫下玉米PEPC和NADK等光合关键酶活性降低,且耐低钾玉米自交系较低钾敏感玉米自交系下降幅度小。Wang等[31]研究发现耐缺钾型大豆品种‘铁丰40’在缺钾条件下叶绿素荧光参数和Rubisco含量不受影响,能够维持较高的光合效率。在本试验中,低钾胁迫显著降低了‘晋谷21号’和‘龙谷25号’叶片Rubisco和PEPC活性,钾非敏感型品种‘龙谷25号’低钾胁迫下Rubisco、PEPC活性高于钾敏感型品种‘晋谷21号’,从而维持较高的光合速率。
将‘晋谷21号’和‘龙谷25号’幼苗的叶片钾含量与生长生理指标分别进行冗余分析,借以进一步明晰两品种在不同低钾胁迫程度下的差异。结果表明‘晋谷21号’在K5处理下主要通过促进Rubisco活性和PSⅡ反应中心活性,进而显著影响光合能力来促进植株生长发育;‘龙谷25号’耐低钾能力更强,在K0处理下仍能维持较高的Rubisco酶活性和净光合速率(Pn),有利于植株生长和叶片的扩展。
综上所述,低钾胁迫下两个谷子品种幼苗叶片净光合速率均显著降低,抑制了谷子幼苗的生长。‘晋谷21号’正常钾浓度处理(K5)主要促进叶片Rubisco活性和PSⅡ反应中心活性,进而显著影响光合能力来促进植株生长发育。K1处理下两品种净光合速率并未明显降低,K1处理对PSⅡ的影响大于PSⅠ,PSⅠ通过较高的Y(Ⅰ)和Y(ND)维持光合系统稳定,通过增加热耗散保护光合机构。随缺钾程度加深,两品种K0.1和K0处理叶片净光合速率降低主要与气孔导度降低、光系统的电子传递与能量转换抑制,以及碳同化关键酶Rubisco、PEPC活性下降等有关。低钾胁迫下谷子叶片Gs下降,PSⅡ反应中心开放程度降低,电子传递受阻,碳同化关键酶活性下降,从而影响其光合作用的正常进行。钾非敏感品种‘龙谷25号’对钾素吸收利用能力较强,低钾胁迫下的叶片钾含量高于‘晋谷21号’,能通过较高的叶绿素含量、PS Ⅱ开放比例、Rubisco活性和热耗散能力来维持较高的光合活性(Pn)。
