DNA感受器AIM2在心血管疾病中的研究*
2022-08-18陆琳吴诚洁涂均楚李玉洁张瑜李杨欣
陆琳,吴诚洁,涂均楚,李玉洁,张瑜,李杨欣
苏州大学医学院附属第一医院 心血管病研究所,江苏 苏州 215006
心血管疾病(cardiovascular disease, CVD),如心肌梗死(myocardial infarction, MI)、心力衰竭(heart failure, HF)和动脉瘤,已成为损害我国居民健康的“头号杀手”。动脉粥样硬化是心肌梗死和心衰的诱导因素[1],也是动脉瘤的风险因素[2]。随着研究的逐渐深入,人们认识到动脉粥样硬化是一种脂质驱动的进行性炎症性疾病[3-5],其特征是脂质沉积于动脉内膜,同时伴有平滑肌细胞和纤维增生、血小板聚集,最终形成粥样硬化斑块,引起血管狭窄或闭塞,导致急性心血管疾病[4]。心肌缺血缺氧时间延长导致心肌细胞坏死,继而释放损伤相关分子模式(damageassociated molecular pattern, DAMP),如线粒体DNA(mitochondrial DNA, mtDNA),以激活模式识别受体(pattern recognition receptor, PRR),导致促炎细胞因子和趋化因子的持续产生,从而触发下游信号级联以招募中性粒细胞、单核细胞和淋巴细胞聚集到心肌损伤部位,出现左心室(LV)功能障碍和LV重塑,这也是HF的两个主要特征[6]。此外,动脉瘤是一种血管疾病,是血管平滑肌细胞(vascular smooth muscle cell, VSMC)凋亡、细胞外基质(extracellular matrix, ECM)降解和免疫炎症细胞浸润导致的主动脉弹性降低和永久性扩张[7]。在动脉瘤中,浸润性免疫炎症细胞,如中性粒细胞、巨噬细胞、T淋巴细胞和B淋巴细胞,会产生一系列促炎细胞因子、基质金属蛋白酶(MMPs)、活性氧自由基等。这些因素可加重主动脉炎症,诱导VSMC凋亡,并导致ECM降解,最终使主动脉内侧变薄[8]。总之,炎症反应对动脉粥样硬化、心肌梗死、心衰和动脉瘤的发生发展至关重要。
黑色素瘤缺乏因子(absent in melanoma 2,AIM2)是调控炎症反应的关键因子,也是胞内DNA模式识别受体,能够特异性识别病原体释放入胞浆中的双链DNA(dsDNA)。受到dsDNA的刺激后,AIM2与凋亡相关斑点样蛋白(apoptosisassociated speck-like protein containing a CARD,ASC)和半胱天冬酶-1(Caspase-1)组成AIM2炎症小体[9-10],将Caspase-1的前体pro-Caspase-1剪切成有活性的Caspase-1。活化的Caspase-1可切割GSDMD,生成GSDMD-C端片段和GSDMD-N端片段,而GSDMD-N端形成聚合物锚定至细胞膜,形成孔道结构,破坏细胞膜完整性,导致细胞焦亡。另外,活化的Caspase-1也能促进pro-IL-1β、pro-IL-18裂解为有活性的IL-1β、IL-18,这两种炎性因子能够从GSDMD-N端介导形成的孔道中释放,募集、激活其他免疫细胞及趋化因子、炎症因子、黏附因子等,最终形成“瀑布效应”,导致炎症反应被正反馈放大。AIM2炎症小体通路的持续激活最终导致组织损伤。更重要的是,有大量证据证明AIM2参与心血管疾病的多种病理过程,包括VSMC和心肌细胞焦亡[11-12]、VSMC迁移[13]、炎性细胞因子产生、巨噬细胞激活[14]等。
本文将概述AIM2的生物学特性,总结影响AIM2炎症小体通路的相关因素。此外,还将讨论AIM2对动脉粥样硬化、动脉瘤、心肌梗死以及心力衰竭的影响,旨在为心血管疾病的治疗找到新的方向。
1 AIM2炎症小体
1.1 AIM2的结构
AIM2最早是在人黑色素瘤细胞株中被发现缺失,且能逆转该肿瘤表型,因而得名“黑色素瘤缺乏因子2”。AIM2是人类染色体lq22上的AIM2基因(1 485 kb)编码的一种344个氨基酸组成的蛋白质(39 kDa),属于干扰素诱导基因家族。AIM2主要定位于细胞质中,有两个重要的结构域:一个是与dsDNA识别与结合有关的,含有2个相邻寡核苷酸/寡糖结合结构域(OB)的C端HIN结构域;另一个是由6个α-螺旋组成的N端间pyrin结构域(PYD)[15]。进化过程中,PYD虽然高度保守,但是在招募ASC时也发挥着类似OB的作用[16]。
1.2 AIM2炎症小体的组装
AIM2是一种模式识别受体(PRR),具有特异性识别抗原信号的功能,识别和结合来自细菌、病毒、疫苗或自身的细胞质dsDNA。这种dsDNA序列独立,在诱导AIM2组装、AIM2 PYD自繁殖和ASC聚合方面具有长度依赖性,通常长dsDNA比短dsDNA更有效,因为组装AIM2细丝螺旋基的概率增加[17]。当AIM2 C末端的HIN-200区识别dsDNA后,AIM2会募集衔接蛋白ASC。ASC作为重要的衔接分子,除了PYD结构域外,还包含一个与Caspase-1的CARD结构域直接相互作用的CARD结构域。衔接蛋白ASC将效应蛋白Caspase-1募集到炎症小体复合物上。AIM2炎症小体的形成使Caspase-1分子彼此靠近,从而产生自催化作用活性Caspase-1分子。随后,活化的Caspase-1将前体细胞因子IL-1β和IL-18切割成成熟的活性形式[18]。此外,Caspase-1通过裂解GSDMD诱导细胞发生焦亡,其特征为渗透失衡、细胞肿胀、膜完整性丧失,最终导致细胞破裂。
细菌、病毒或受损线粒体释放的dsDNA与AIM2结合,诱导由AIM2、ASC和pro-Caspase-1组成的AIM2炎症小体的组装。活化的Caspase-1切割pro-IL-1β、pro-IL-18和GSDMD,介导炎症反应和细胞焦亡。细菌DNA释放到细胞质中依赖于GTP酶介导的溶菌作用,其表达由cGASSTING轴激活,也由细胞质dsDNA激活(图1)。
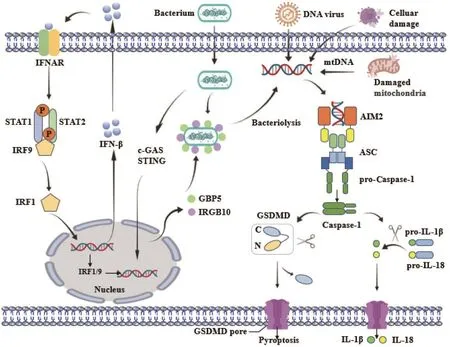
图1 AIM2炎症小体激活图
1.3 AIM2的表达及其作用
AIM2的表达直接影响其功能。研究表明,通过AIM2干扰素敲低AIM2的表达后,显著降低了人单核-巨噬(THP-1)细胞、小鼠巨噬细胞系(B6 MCLs或N3-KO-MCLs)和骨髓源性巨噬细胞中合成DNA poly(dA: dT)介导的IL-1β释放和Caspase-1激活[9-10,19],同时也减弱了293T-ASC-caspase-1细胞的死亡[9]。
早期研究发现,在人肠道、睾丸和脾脏组织以及外周血白细胞中能够微弱地检测到AIM2 mRNA。随着研究的深入,研究人员发现AIM2在正常颈动脉和主动脉的内膜内皮细胞(EC)和中膜平滑肌细胞中也表达[20]。此外,研究发现在感染、炎症或动脉粥样硬化的情况下,AIM2的表达在心血管系统的许多细胞类型中被诱导上调(表1)。例如,AIM2基因表达在THP-1 M1极化巨噬细胞中上调[21],导致炎症反应持续。在培养的原代人主动脉内皮细胞(HAoECs)和人主动脉平滑肌细胞(HAoSMC)中,暴露于促炎细胞因子(如肿瘤坏死因子-α(TNF-α)和干扰素-γ(IFN-γ))或dsDNA刺激后,AIM2表达迅速升高[20]。此外,dsDNA在晚期动脉粥样硬化病变中大量积累[21-22],表明dsDNA对动脉粥样硬化斑块中AIM2表达的增加起着关键作用。研究人员通过用高脂饮食饲养小鼠后发现其AIM2表达增加,其中氧化低密度脂蛋白(oxLDL)是一种致动脉粥样硬化风险的因子,可以通过转录因子核因子-κB(NF-κB)信号转导增加AIM2蛋白的表达[11],而miR-485通过直接结合AIM2的3'-UTR,在转录后抑制AIM2的表达[23]。H9C2心肌细胞中AIM2的蛋白表达在高糖刺激下显著增加,而在活性氧抑制剂的作用下显著降低[12]。另外,AIM2去甲基化能上调AIM2基因的表达[24]。

表1 AIM2表达的影响因素
1.4 AIM2炎症小体激活的机制
随着研究的深入,越来越多的证据表明线粒体功能障碍、高胆固醇含量、高密度脂蛋白(high-density lipoprotein, HDL)水平、干扰素、细菌与病毒的感染在调节AIM2炎症小体激活中发挥着重要作用。因此,下文重点讨论这些AIM2炎症小体激活的分子机制及相关作用。
1.4.1 线粒体功能障碍
大量研究证实,线粒体功能障碍通过调节AIM2炎症小体活性,在动脉粥样硬化、心肌梗死和心力衰竭等心血管疾病的发病机制中发挥作用。线粒体功能障碍通常会增加活性氧(reactive oxygen species, ROS)的产生[25],而线粒体来源的ROS(mtROS)诱导AIM2炎症小体的激活。研究表明,线粒体特异性ROS清除剂mitoTEMPO能抑制弗朗西斯菌感染的骨髓源性巨噬细胞中Caspase-1裂解和IL-1β分泌[26]。此外,mtROS的异常增加会导致线粒体结构和功能障碍,使mtDNA溢出到细胞质中。例如,细胞内25-羟基胆固醇水平不足导致胆固醇过度积累,线粒体呼吸和膜极化受损,mtDNA被释放到胞质中进一步激活AIM2炎症小体。
受损的线粒体能释放磷酸甘油酯变位酶家族成员5(PGAM5)[27],这是一种线粒体蛋白磷酸酶,是线粒体功能障碍的驱动因素,加速ASC聚合和活性氧生成,促进骨髓来源的巨噬细胞中AIM2炎症小体活化和IL-1β分泌[28]。PGAM5还与BCL2样1(BCL2L1)的BH3结构域结合,通过与BCL2L1相互作用,抑制含FUN14结构域的蛋白1(FUNDC1)去磷酸化,最终抑制线粒体自噬,这是线粒体失调的关键原因[29]。mtROS增加也与线粒体自噬功能缺陷有密切关联[30]。线粒体自噬缺陷增加了mtDNA的释放,导致糖尿病患者心肌细胞和心肌巨噬细胞中AIM2炎症小体的过度激活[31]。
以上结果证明线粒体功能障碍是AIM2炎症小体激活的驱动因素,而AIM2炎症小体的激活又增加线粒体活性氧的产生、线粒体的肿胀和管状线粒体网络断裂,这些都加剧线粒体的损伤[32]。因此,线粒体功能障碍导致AIM2炎症小体激活的机制是一个反馈回路,即线粒体功能障碍诱导mtROS产生以激活AIM2炎症小体,进而抑制线粒体自噬,诱导线粒体损伤。
1.4.2 胆固醇含量升高和高密度脂蛋白水平降低
研究表明,胆固醇含量升高是通过诱导线粒体功能障碍,从而激活AIM2炎症小体。在脂多糖(lipopolysaccharide, LPS)激活的小鼠骨髓源性巨噬细胞中,胆固醇导致线粒体/线粒体相关膜胆固醇含量增加,mtROS生成增加,mtDNA释放增加,从而触发AIM2依赖性IL-1β的产生,而AIM2缺失或mtDNA缺失会显著减弱因胆固醇增加而诱导IL-1β产生导致的炎症反应[33],表明胆固醇含量升高后通过诱导线粒体受损从而引发AIM2炎症小体激活。相反,胆固醇25羟化酶(Ch25h)是一种从胆固醇中合成氧化甾醇25羟基胆固醇(25-HC)的酶,能抑制AIM2炎症小体的激活。25-HC参与抑制甾醇调节元件结合蛋白2(SREBP2)和胆固醇生物合成的过程。Ch25h通过维持对SREBP2激活和胆固醇合成的抑制过程来避免线粒体功能障碍,从而起到抑制AIM2炎症小体激活的作用[33]。
与总胆固醇、低密度脂蛋白胆固醇相比,HDL被称为“良好的胆固醇”,具有预防动脉粥样硬化或其他炎性疾病的作用。HDL最重要的抗动脉粥样硬化作用是通过其作为特定的胆固醇受体,将多余的胆固醇从外周组织的细胞中运输出去,从而促进胆固醇的逆向运输[34]。最近,HDL也被证明可以挽救由胆固醇含量升高引起的线粒体呼吸减弱[33],为HDL调节AIM2炎症小体激活提供了一个可能的作用机制。此外,在人类免疫缺陷病毒1型(HIV-1)感染患者中,HDL水平显著降低,与AIM2、ASC、IL-1β和IL-18的mRNA水平呈负相关,表明低水平的HDL可能诱导AIM2炎症小体的激活[35]。然而,AIM2炎症小体和HDL之间的关系还需进一步研究。
1.4.3 干扰素(interferon, IFN)
研究报道,cGAS-STING信号通路与AIM2炎症小体激活密切相关,并在炎症反应和动脉粥样硬化中发挥重要作用[36-37]。I型干扰素,尤其是IFN-β,被描述为cGAS-STING通路的标志性输出信号。
干扰素在AIM2炎症小体信号转导中通过两种机制发挥关键作用。首先,在cGAS-STING信号中,I型干扰素通过结合并激活I型干扰素受体(IFNAR)发挥作用[37]。在金丝雀痘病毒载体ALVAC感染期间,cGAS通过STING I型IFN途径促进AIM2启动和AIM2炎症小体激活[38]。除了转导上游信号外,带有IFNAR的I型干扰素可以通过上调AIM2表达直接促进AIM2炎症小体的激活。其次,I型IFN信号转导已被证明可促进细菌感染期间的细菌裂解和配体可用性[39]。在细菌感染期间,STING会诱导鸟苷酸结合蛋白(GBP)的表达,如GBP2、GBP3。GBP2和GBP3是两种I型IFN相关基因,它的信号转导会引起细菌裂解,导致细菌DNA释放到胞质中并激活AIM2[40-41]。
1.4.4 细菌与病毒
感染在心血管疾病的发展中也起着重要作用[42],例如在动脉粥样硬化斑块中观察到50多种不同的细菌种类和细菌DNA[22,43]。多种病毒和细菌与AIM2炎症小体的激活有关。AIM2不是序列特异性DNA传感器,因此除了自身DNA,来自微生物的DNA同样能够激活AIM2炎症小体。研究显示,巨细胞病毒[44]诱导小鼠骨髓来源的树突状细胞和巨噬细胞中AIM2炎症小体的激活,能增加IL-1β或IL-18的释放,加剧动脉粥样硬化[39]。一项体内研究证明,巨细胞病毒足以在C57BL/6和AIM2+/+小鼠中诱导系统性IL-18产生,但在AIM2-/-小鼠中不足以诱导,这表明巨细胞病毒以AIM2依赖的方式诱导AIM2炎症小体活化[39]。牙龈卟啉单胞菌可加剧Apoe-/-小鼠动脉中的炎症反应[45],通过激活TLR2和TLR4信号来增加THP-1细胞中AIM2的表达和AIM2炎症小体的激活[46]。
2 AIM2与心血管疾病
心血管疾病是人类死亡的主要疾病之一,而炎症小体引发的炎症反应逐渐成为动脉粥样硬化、动脉瘤、心肌梗死和心力衰竭的标志之一。AIM2被认为在感应dsDNA方面起着关键作用,导致AIM2炎症小体激活和炎症反应,这在感染条件下是有益的。然而,其持续激活可导致慢性炎症以及组织损伤,从而诱导相关炎性疾病的发生[47]。在这里,我们重点讨论AIM2在动脉粥样硬化、动脉瘤、MI以及HF中的重要作用。
2.1 AIM2与动脉粥样硬化
大量研究表明,AIM2在动脉粥样硬化的发展过程中发挥着重要的作用。AIM2在人类颈动脉粥样硬化斑块中表达上调,定位于坏死核心区域附近,与平滑肌肌动蛋白(SMA)和CD68重叠[20]。巧合的是,AIM2在小鼠晚期动脉粥样硬化斑块中的表达也显著上调,并与dsDNA共定位[48]。这表明AIM2参与动脉粥样硬化的发展。另外,研究者在向Apoe-/-小鼠注射AIM2的慢病毒后,发现AIM2的过度表达扩大了主动脉病变面积。然而,在高脂饮食12周后,通过RNA干扰沉默AIM2可减少Apoe-/-小鼠的主动脉病变面积[11],表明AIM2能直接导致动脉粥样硬化。
尽管AIM2加速动脉粥样硬化斑块进展的确切机制尚不清楚,但研究表明有几种可能机制。首先,AIM2可以提高Apoe-/-小鼠的细胞间黏附分子1(ICAM-1)的水平[11]。ICAM-1通过结合淋巴细胞功能相关抗原-1(LFA-1)促进单核细胞黏附,因此LFA-1被认为是动脉粥样硬化形成的关键因子。小鼠ICAM-1的靶向性缺失已被证明可以减少主动脉的病变[49]。基于这些发现,ICAM-1水平升高可能是AIM2诱导动脉粥样硬化的一种机制。其次,AIM2能够触发VSMC迁移和焦亡[11,13]。VSMC是正常动脉壁中固有的细胞类型,也是动脉粥样硬化斑块的重要组成部分,在动脉粥样硬化的不同阶段发挥着不同的作用。早期动脉粥样硬化斑块中的VSMC被认为是通过迁移从局部内侧VSMC衍生而来,并已被证明能够增殖、转换表型、ECM脂质积累、形成泡沫细胞和趋化因子,所有这些都表明内膜VSMC是早期动脉粥样硬化的关键驱动因素[50]。值得注意的是,一项体外迁移实验表明,AIM2的过度表达增强了VSMC的迁移,这可能是通过上调MMP-2来实现的。MMP-2是VSMC迁移的关键调节因子,然而,AIM2沉默减弱了VSMC的迁移,证明AIM2参与动脉粥样硬化的形成[13]。另外,AIM2通过增加促炎因子的释放来强化动脉粥样硬化病变中的炎症反应,而这些促炎因子是动脉粥样硬化形成的特征[51]。最近的研究证明,Apoe-/-小鼠中AIM2的缺失可降低动脉粥样硬化病变中的IL-1β和IL-18水平[48]。总之,这些发现表明,AIM2导致动脉粥样硬化作用,部分依赖于IL-1β和IL-18的释放。值得注意的是,在Apoe-/-小鼠中,A151导致动脉粥样硬化病变中IL-1β和IL-18阳性细胞减少。A151是一种AIM2拮抗的合成寡核苷酸[48],提示开发AIM2抑制剂具有治疗潜力[52-53]。
因此,动脉粥样硬化病变中dsDNA和AIM2的存在表明,活化的AIM2炎症小体是产生IL-1β和IL-18的重要途径,是动脉粥样硬化病变中炎症反应的导火索。
2.2 AIM与动脉瘤
临床数据有力地证明了AIM2与动脉瘤的发展有关。研究显示,与对照组相比,动脉瘤患者中AIM2的表达更高,AIM2炎症小体信号通路被激活的程度更强。在动脉瘤患者中,AIM2在其外周血白细胞(包括单核细胞、B淋巴细胞和T淋巴细胞)中的表达显著增加。此外,动脉瘤患者外周血单个核细胞中的AIM2对poly(dA:dT)DNA更敏感,更容易激活。这一点可以从体外培养的动脉瘤患者外周血单个核细胞中IL-1β的产生量高于对照患者[54]中得到证明,这意味着AIM2在动脉瘤形成中可能发挥作用。通过活检对人类动脉瘤样本进行研究,结果表明,与对照组相比,AIM2在人类动脉瘤样本病变中的表达显著上调[38,55]。这进一步证实AIM2参与动脉瘤的发生过程。此外,与正常主动脉壁相比,动脉瘤主动脉壁中ASC、Caspase-1和IL-1β的基因表达显著增加,对ASC、Caspase-1、Caspase-5和AIM2的免疫染色显示,它们在动脉瘤样本的淋巴细胞以及位于外中膜和外膜的淋巴聚集物中表达,表明AIM2炎症小体在动脉瘤病变中被激活,而淋巴细胞可能是动脉瘤病变中AIM2炎症小体的来源[56]。
最近,科研人员对动脉瘤患者病变处分离得到的VSMC进行了研究。结果表明,IFN-γ启动后,AIM2、NLRP3、IFI16(干扰素诱导蛋白16)和Caspase-1的mRNA表达上调,而动脉瘤VSMC对来自相邻动脉瘤VSMC的坏死细胞碎片的反应增强,导致AIM2的表达进一步增加,相应NLRP3和Caspase-1蛋白表达也增加[57]。因此,T细胞产生的IFN-γ与坏死细胞碎片协同触发主动脉VSMC中AIM2炎症小体激活,在动脉瘤形成过程中起关键作用。此外,考虑到AIM2炎症小体在细胞焦亡中的作用,有观点认为AIM2炎症小体和坏死细胞碎片的激活会导致一个恶性循环。在这个恶性循环中,AIM2炎症小体诱导VSMC死亡,而VSMC死亡又反过来促进AIM2炎症小体的激活,从而协同加速动脉瘤的进展。
我们之前已提到,AIM2上调了Apoe-/-小鼠腹主动脉中MMP-2的表达[13],并诱导IL-1β和IL-18的产生[48]。MMP-2也称为明胶酶A,其作用是降解弹性蛋白[58],而弹性蛋白断裂是动脉瘤的标志之一[7]。与此一致的是,小鼠模型中MMP-2的敲低不仅延迟了由CaCl2诱导的主动脉瘤的发生,而且延迟了主动脉瘤的破裂[58]。IL-1β已被证明能增加VSMC中MMP-2的活性[59],并加速动脉瘤的发生[60-61]。此外,AIM2炎症小体的激活诱导VSMC发生焦亡[11],而VSMC的减少是动脉瘤发展的关键事件[7]。因此,我们推测AIM2可能通过增加MMP-2的表达、促进IL-1β和IL-18的释放,并导致VSMC死亡来加速动脉瘤的发展,但这一推测还需进一步证实。
上述研究已证明AIM2与动脉瘤的发生发展有密切联系,然而其中的因果关系仍未得到很好的阐述,因为现有的研究几乎都是基于人群的病例对照研究,缺乏在细胞、分子水平的深入研究。AIM2在动脉瘤中的动物模型和抑制剂的合成及应用将是未来研究的一个重要领域。
2.3 AIM2与心肌梗死、心力衰竭
研究表明,AIM2参与心肌梗死和心力衰竭[62-64]。心肌缺血/再灌注(I/R)损伤后小鼠心肌组织AIM2表达增加,而AIM2的过度表达又加剧I/R诱导小鼠心肌损伤[63]。IL-18是AIM2炎症小体下游效应器,也参与心肌梗死和I/R损伤。心肌梗死小鼠的心脏IL-18 mRNA表达水平显著上调[65-66],而降低IL-18的表达可减少I/R后的梗死面积[65],证明IL-18在心肌梗死中的病理作用。另外,AIM2通过诱导细胞焦亡,释放IL-1β、IL-18[23],会加重脑I/R损伤,也间接证明AIM2可能通过激活AIM2炎症小体和释放IL-18加剧I/R诱导的心肌损伤。
通过结扎左冠状动脉前降支,诱导2型糖尿病小鼠发生心肌梗死的研究显示,小鼠心脏左室射血分数降低、线粒体功能受损、mtDNA向细胞质释放增加、AIM2表达增加、Caspase-1过度激活、梗死周围或左室梗死区心肌细胞死亡,以及循环IL-18水平升高,与心肌梗死患者中的AIM2炎症小体持续激活一致[31]。AIM2炎症小体的持续激活,需要心肌细胞以线粒体功能缺陷依赖的方式从左室梗死周围区域释放mtDNA。
糖尿病心肌病是心力衰竭的重要原因之一。在链脲佐菌素诱导的糖尿病大鼠模型中,心脏组织中AIM2表达显著上调,表明AIM2在糖尿病心肌病中起重要作用,而沉默AIM2可改善心功能不全,降低I型胶原和III型胶原蛋白水平。而且,在高糖处理的H9C2心肌细胞中,AIM2的沉默降低了GSDMD-NT相关的细胞焦亡,表明AIM2通过诱导心肌细胞的焦亡和心室重塑,加剧糖尿病心肌病[12]。
目前研究显示,我们对AIM2在MI和HF中具体机制的了解仍不全面,因此AIM2在这些疾病中的精准机制及确切作用将是未来研究的重要领域。
3 靶向AIM2信号通路在疾病中的治疗潜力
尽管AIM2的病理作用尚未完全阐明,但几项针对AIM2信号通路的研究结果令人振奋。Yan等[63]揭示了miR-219a可介导心肌缺血/再灌注小鼠中AIM2和TLR4/MyD88通路的缺失,并用七氟醚治疗心肌缺血/再灌注小鼠,发现其显著减小梗死面积,抑制心肌细胞死亡。此外,Gao等[67]发现穿心莲内酯(一种从穿心莲中提取的活性成分)能够通过抑制AIM2感知dsDNA,靶向抑制AIM2炎症小体激活,显著减轻C57BL/6小鼠中辐射诱导的肺组织损伤、炎症反应和纤维化。另外,组蛋白脱乙酰酶3(HDAC3)的选择性抑制剂RGFP966对AIM2炎症小体的抑制被证明对缺血性中风有治疗作用[68]。Chung等[69]发现红葡萄藤叶提取物产品EFLA 945能够通过阻止DNA进入巨噬细胞,抑制AIM2炎症小体和Caspase-1激活,以及IL-1β和IL-18的释放,进而降低炎症反应和银屑病的严重程度。此外,卡那单抗对其下游效应因子IL-1β的抑制降低了心肌梗死患者心力衰竭的概率[70]。这些发现表明,靶向AIM2信号通路可能有助于预防动脉粥样硬化、动脉瘤、MI和HF。
4 问题与展望
AIM2被认为是dsDNA传感器,是AIM2炎症小体激活和宿主防御所必需的。如上所述,AIM2炎症小体的激活会导致促炎因子的产生和释放、细胞焦亡,以及ICAM-1和MMP-2表达增加,并参与动脉粥样硬化、动脉瘤、MI、HF等炎性疾病的发病过程。然而,AIM2在这些疾病中的确切作用和机制仍然是个谜团。因此,需要更多的研究来进一步阐明AIM2的确切作用,同时,建立细胞特异性敲低或过表达AIM2的动物模型将有助于我们深入了解AIM2在上述疾病的各个阶段所发挥的不同作用。此外,AIM2炎症小体与上述心血管疾病之间的因果关系仍未确定,AIM2是否是导致动脉粥样硬化、动脉瘤、MI和HF的直接因素,或者这些疾病的发生是否依赖于AIM2炎症小体的激活,亟需深入探讨。同时,寻找靶向抑制AIM2的药物也是未来研究的一个重要领域。综上,进一步了解AIM2的作用和机制将有助于开发新的治疗策略,以抑制动脉粥样硬化、动脉瘤、MI和HF。
作者贡献声明 陆琳,综述构思及撰写,图表绘制;吴诚洁, 综述构思及修订;涂均楚,李玉洁,张瑜,综述修订;李杨欣,综述构思,图表设计及修订。
