海洋风暴沉积的微体生物研究进展
2022-08-15戚洪帅沈林南张爱梅王承涛
陈 敏,戚洪帅,沈林南,张爱梅,王承涛,方 琦
(1.自然资源部第三海洋研究所,福建 厦门 361005; 2.自然资源部北部湾滨海湿地生态系统野外科学观测研究站,广西 北海 536015; 3.厦门市国土资源与房产管理局,福建 厦门 361013)
热带风暴是全球最主要的海洋自然灾害之一。全球每年发生80~100个热带风暴,而中国是受热带风暴影响最严重的国家之一[1-2]。在我国历史上,单次热带风暴造成的死亡人数超过10万人的有8次,其中在1696年长江三角洲发生的风暴潮曾导致十多万人死亡[3]。
人类通过仪器对热带风暴资料进行记录的年限较短,不超过150 a。强台风的危害极大,其形成后毁灭性极强,因此人们希望能够掌握其形成和活动规律。随之而来的就是古风暴学的发展,通过一些地质记录和历史文献来研究地质历史时期以来古风暴的活动规律[4]。
早在二十世纪五十至六十年代,地质学家就开始对现代风暴沉积进行研究。当时的研究表明,由于暴风浪的影响,沉积物的运移和搬运都会出现变化[5-7]。二十世纪七十至八十年代风暴沉积研究达到了顶峰,其中最著名的就是提出了陆架风暴沉积理论,该理论对风暴沉积学的影响重大,在沉积学的历史长河中具有里程碑式的意义[8-9]。古风暴学在二十世纪九十年代逐渐出现在人们的视野中,作为一门新兴学科,其主要是通过沉积物中的地质记录,揭示几百年甚至几千年前热带气旋活动的规律。地质记录里的古风暴一般具有作用时间短、形成突然、强度大等特点,同时也会保留一些标志性的特殊古生物化石[10-14]。风暴期间产生较强的风浪会越过较低的障壁沙咀/岛进而改造后方海湾和潟湖沉积物,其高强度的改造作用远强于正常天气条件[2]。在台风盛行期间,其产生的大浪及风暴潮对沉积物的搬运作用急剧增强。风暴期间的向岸流和冲越流会携带较多海相微体生物,使其沉积在海岸后方的湖沼环境中;可根据这一特征利用微体化石来标识沉积层的风暴成因[4]。因此微体化石作为一项重要的指标,可以用来指示风暴沉积,并很好的判断其特征[15-19]。
低能沉积环境中的沿海沉积物记录了古风暴的地质证据,这些事件的许多最佳重建方法都来自微体生物(硅藻、有孔虫和花粉)组合的变化[20]。本文对目前国内外风暴沉积的微体生物研究进展进行综述,以期为风暴沉积研究提供借鉴和参考。
1 现代风暴沉积的微体生物特征
在现代台风风暴过程中的微体生物研究目前还较为有限,可查询到的文献记录较少,且所获样品一般距离台风中心有一定距离,很难恰巧位于台风登陆点附近。
1.1 不同沉积环境中微体生物的沉积特征研究
前人在从后滨到内陆架的各种不同沉积环境中开展了风暴过程微体生物的沉积特征研究(表1)。
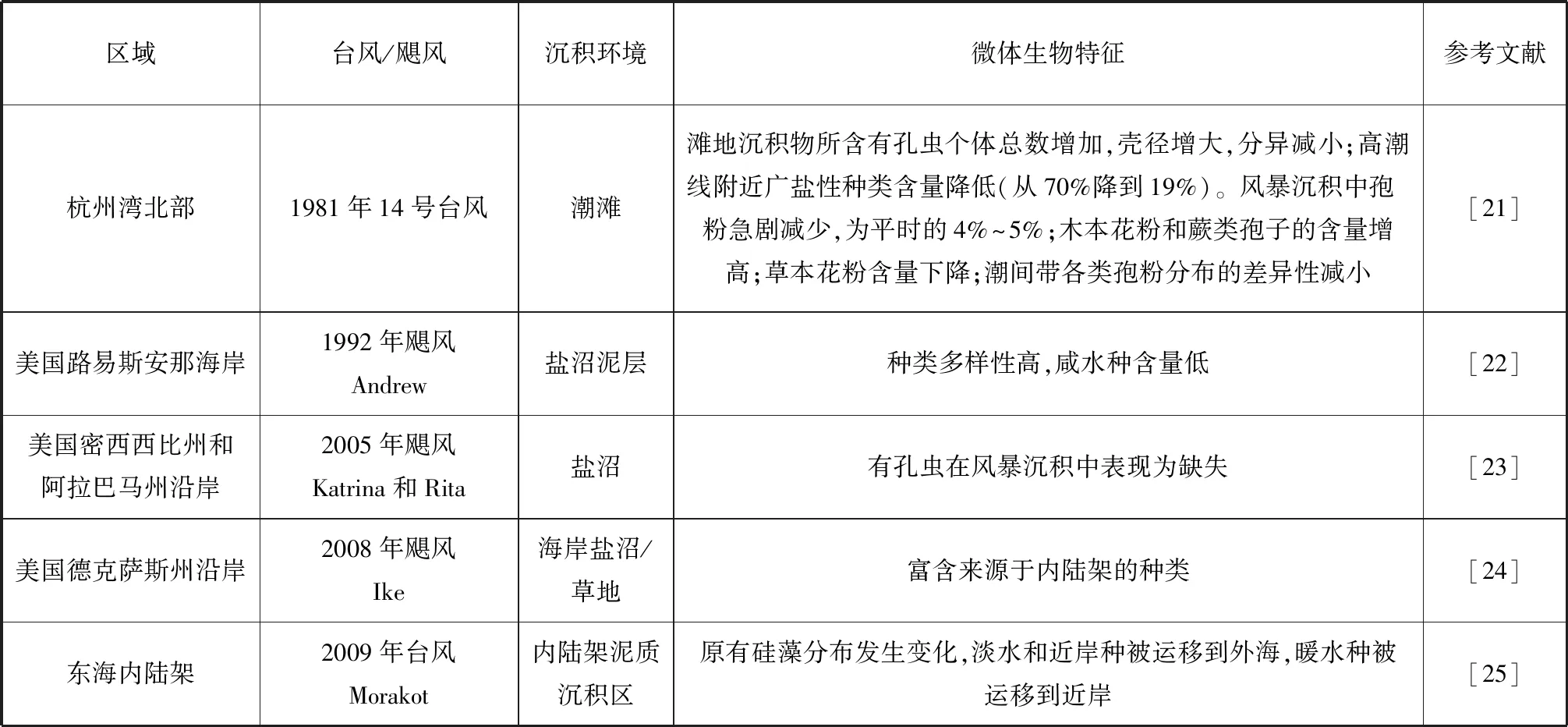
表1 现代风暴沉积的微体生物特征
首先,许多研究集中在海岸后方的盐沼中。盐沼中的沉积环境较为稳定,台风风暴盛行期间,会伴随强烈降水和巨大的风浪作用,沉积物大多被搬运和沉积在盐沼中,形成冲越沉积,这些沉积中的生物特征与正常天气下明显不同。Hawkes等(2012)比较了得克萨斯州加尔维斯顿和圣路易斯群岛的三个地点由飓风Ike(2008年9月13日登陆)造成的冲越沉积物的数量、粒度分布、有机质含量和有孔虫群落;该次风暴潮的高度在2.7~3.7 m,向陆淹没范围为113~330 m;在风暴过程中,只有10%~30%的风暴侵蚀沉积物以冲越的方式沉积下来,而其余70%~90%的风暴侵蚀沉积物则重新沉积在近岸环境中;风暴之前的草地/盐沼土中几乎不含有孔虫,而在风暴沉积的冲越砂中则含有墨西哥湾海湾和近岸环境特有的种类Ammoniaspp. 和Elphidiumspp.,以及常常出现在内陆架环境的Bolivinasubaenariensis、Quinqueloculinaseminulum和其他浮游性种类,表明这些冲越沉积来源于近岸和内陆架区域,由此估算出了风暴沉积砂层厚度约为2~28 cm;在该项研究中发现仅凭沉积学和地层学数据无法分辨风暴沉积层,因此有孔虫的应用是非常重要的[24]。这也许给了我们一些启示,在古风暴沉积记录中微体生物化石记录是否比一些沉积学记录更灵敏。另一个不同的例子来源于1992年的飓风Andrew。该飓风登陆美国路易斯安娜海岸之后,在盐沼中发现的风暴沉积层具有较高的硅藻种类多样性,较低的海水种含量和氮含量,同时风暴沉积物分选较差;这与之前的冲越砂研究有所不同,没有出现非常多的近岸和内陆架种,这是因为在风暴期间,各种不同来源的硅藻发生了迁移和混合,导致没有一种类群能占据主导地位,所以种类多样性高;而海水种减少和淡水种增多与飓风洪水和附近半咸水、淡水区域在飓风期间的物质搬运输入有关[22]。而来自于2005年飓风季节美国密西西比州和阿拉巴马州沿岸的盐沼沉积记录表明,在风暴沉积物中几乎没有有孔虫,而在盐沼沉积物中发现了大量代表盐沼环境的有孔虫类群;风暴沉积物中缺失有孔虫这一现象可能是由于其在海滩砂和沙丘砂中本来就很稀少,还有可能是因为沉积后变化造成的[23]。由此可见,沿岸盐沼的风暴沉积中微体化石以外来混合为特征,而具体情况根据它们不同的物源而有所差异。
其次,我国学者在潮滩上的研究表明,在1981年14号台风(中心最高风力达到12级以上,形成了巨大的风暴潮,潮位高达5.59 m,超过了此前当地历史上实测潮位的最高记录)经过杭州湾后,杭州湾北部滨岸潮滩上的风暴沉积中有孔虫和孢粉的基本特征出现了变化,风暴潮作用期间有孔虫个体数量明显增加,且其组合和分布也有明显变化,个体数达到平时的2~6倍,壳体直径变大,在高潮线附近广盐性种类数量明显下降,从台风前的70%以上降至19%;从有孔虫组合上看滨海沼泽型被强潮河口型(主要是浮游有孔虫)代替,从而导致差异性变小[21]。高潮坪有孔虫数量和浮游种类的增多说明台风将离岸水体中的有孔虫携带到潮坪上。从孢粉特征来看,台风前草本花粉含量由潮间带向海逐渐减少,而木本和蕨类则相反,台风过后虽然主要种属不变,但组合与分布发生一定变动;在特大风暴潮期间沉积物中孢粉数量急剧降低,仅为平时的4%~5%;蕨类和木本含量增高,高潮坪增加幅度超过了20%;草本花粉含量降低幅度较大,在高潮坪可达约50%;木本、草本和蕨类在潮间带沉积中含量相近,没有明显的分布差异[21]。在台风强烈水动力的扰动下,不难发现潮滩上的沉积物发生了垂直海岸方向的迁移和混合,因此其孢粉种类组成更均匀,差异性更小。
再次,Chen等(2019)在东海内陆架泥质区对2009年台风Morakot前后表层沉积硅藻的变化进行了研究,在台风发生前后,没有观察到优势种的显着变化;但硅藻组合特征存在变化,表明台风期间沉积物在大范围内发生了运移和混合;由于台风期间强大的水动力条件,原有的代表台湾暖流的硅藻组合与代表沿海水体的硅藻组合之间的界线消失了,淡水种甚至被运移到离岸100 km处,原有的季节性模式被颠覆(图1);这表明台风可以大大改变东海内陆架上表层沉积物的分布[25]。

图1 东海内陆架区域2009年夏季台风Morakot前后表层沉积硅藻的变化Fig. 1 Changes in surface sedimentary diatoms in the inner continental shelf of the East China Sea before and after Typhoon Morakot in summer 2009本图修改自文献[25]。
1.2 微体生物信号的留存情况研究
现代风暴发生后,这些当时的标志性微体生物信号多大程度上能留存下来以及它们可否保存在地层中以供人们识别是一个重要的问题。这方面的研究目前非常少,可以肯定的是在较高能的环境下风暴的标志性微体生物信号留存困难。在低能海岸潟湖或盐沼环境下的留存研究表明,登陆1 a后飓风Andrew对沉积物组成没有影响,沉积过程恢复到飓风前的特征;但飓风发生2 a后,硅藻种群仍然没有恢复到飓风前的状况,飓风通过减少沉积物中的硫化物含量使得硅藻种群向附生种为主发生了转变[22]。在Hawkes等的研究中,也观察到了上一个风暴事件(1983年的Alicia飓风)带来的异地有孔虫,同时伴随着低的有机质含量(约5%)和高的砂含量[24],显然这次风暴记录与之后2008年发生的飓风Ike风暴记录之间存在空白。因此,什么样的风暴?能在哪里?如何被保存下来?还是一个需要科学家继续挖掘研究的挑战。
2 古风暴沉积的微体化石特征
相对于现代风暴沉积的研究而言,海岸古风暴沉积的微体化石研究更为丰富,国内外有许多代表性研究(表2)。微体化石主要被用来帮助识别古风暴沉积层或古风暴事件,从而探讨历史时期特定区域的风暴历史和频次,进行古风暴重建。

表2 古风暴沉积的微体化石特征
2.1 古风暴层微体化石特征的研究
前人很多研究探讨了推断的古风暴层中微体化石的特征。许多学者认为风暴层常常具有高的有孔虫数量和种类多样性[17,19,31]。许世远等(1989)在长江三角洲的沼泽低地、贝壳沙堤、三角洲前缘和前三角洲等处都发现了风暴沉积;其中,在上海查山等地的剖面揭示出风暴沉积中陆相与海相动植物遗体共存,出现一定数量的广盐性有孔虫,以Ammoniabeccarii为主;在上海南库贝壳沙堤等地的风暴沉积中,富含Ammoniaanneetens等大个体有孔虫,直径为0.5~0.7 mm,壳体多破碎或磨蚀[26]。黄光庆等(1997、1998)通过对珠江口沉积物中有孔虫进行分析,发现正常海况下有孔虫种类较少,主要为广盐性有孔虫,壳体分异度低,优势度高,多见畸形壳体,无浮游种;而在台风风暴沉积中有孔虫的特点有明显变化,其壳体分异度高,优势度低,广盐性和窄盐性有孔虫混杂分布,且个体较小,壳径与沉积物粒径分布呈正相关关系,与现代河口原地埋藏群不同,能够识别出这些有孔虫为异地埋藏群(图2)[15-16]。
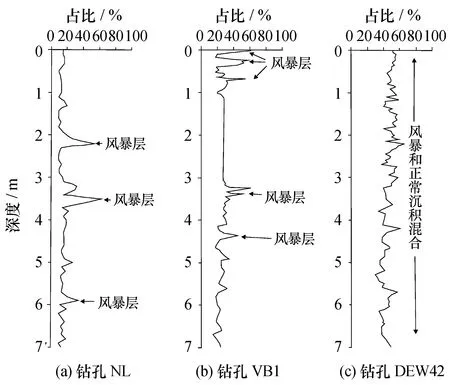
图2 珠江口钻孔中的异地有孔虫含量和风暴沉积层Fig.2 Content of heterochthonous foraminifera and storm deposits in cores in the Pearl River Estuary本图修改自文献[15],钻孔NL和钻孔VB1位于珠江口内,钻孔DEW42位于珠江口外。
为了找出风暴在不同登陆地点(风暴登陆中心和登陆点50 km外)沉积痕迹的差异性,Collins等(1999)对美国南卡罗莱纳州海岸沉积进行了研究,发现1989年9月飓风Hugo在登陆点附近的Prices Inlet地区形成了厚约8 cm的风暴砂层,其中含有许多来自于近海的有孔虫;在飓风登陆点50 km外的Sandpiper地区,可以看到夹在淡水有孔虫层之间的近海有孔虫层,但是X射线或沉积学研究都没有检测到该风暴事件的痕迹[17]。Hippensteel等(1999)在美国南卡罗来纳州Folly岛的盐沼沉积中发现了现代风暴沉积中混有近海和渐新世—中新世的有孔虫,这些有孔虫来源于近岸沉积的侵蚀[18]。Lane等(2011)通过对美国佛罗里达州沿海潟湖沉积的研究,发现了长达4 500 a的飓风引发的风暴潮记录,有些起源于距离海岸至少5 km的海洋有孔虫存在于沿海的古风暴层中,离岸有孔虫的存在和长期保存表明基于微体化石的古飓风重建是可行的[29]。这部分研究大多数来源于有孔虫,而关于硅藻,商志文等(2013)曾提到在渤海湾西岸的钻孔中有一层位可能受风暴事件影响,其中硅藻保存较差,属种也相对单一[32]。
2.2 利用微体化石进行的古风暴重建
利用有孔虫、硅藻、孢粉等进行的海岸古风暴重建研究主要是在沿岸沼泽和潟湖中进行。研究者们主要通过微体化石和地层学等证据相结合,识别全新世的古风暴层,并研究全新世风暴活动的时空变化、大小和频率,尤其是百年甚至千年尺度的气候变化与风暴的关系,从而可以评估长期的生态影响[20]。
在英国Morecambe湾滨岸湖沼相进行的孢粉和硅藻地层学研究中,Zong等(1999)识别出(6 030±70)~(5 740±60) a BP之间的9次强风暴沉积事件;在这些风暴事件中海洋硅藻从河口通过水道或越过沙坝被冲刷至后方潟湖环境,海洋硅藻的峰值被解释为风暴潮条件下堆积的结果;孢粉数据则给出了植被变化的一些信息[27]。而在美国墨西哥湾沿岸阿拉巴马州一个沼泽沉积层的花粉研究表明,阿拉巴马州海岸在过去1 200 a中,两次被4级或5级飓风直接袭击,每年登陆的概率为0.17%;飓风来袭后盐生植物(藜科)和灌木(杨梅科)的种群扩大了,这可能是由于盐水冲入沼泽地以及过度冲刷过程导致土壤盐渍化所致;每次强烈飓风和随之而来的大火之后,松树种群均显著减少,这表明反复发生的飓风与火灾相互作用导致树木死亡[28]。在亚洲地区,Sato等(2016)通过对日本中部沿海潟湖沉积进行硅藻化石分析重建了其全新世晚期的环境变化,根据硅藻的丰度确定了两个事件沉积层,在其中一层中海水种或半咸水种Glyphodesmiswilliamsonii占主导地位,该层被解释为海啸或风暴潮造成的短暂沉积[30]。
3 研究地点的选择与化石适用性的探讨
综上所述,大多数风暴研究需要选择在沿岸的低能环境下,尤其以湖泊[33]、沼泽[19,22-23,34]、沿海池塘[17,35]、潟湖/海湾[36]和沿海洼地[29]为佳。而较高沉积速率的河口也能保存有较好的风暴沉积层,例如在长江口风暴作用产生的沉积物会被后期常态沉积掩埋,从而避开之后水动力作用的改造;但在长江河口叉道地区水动力条件较强,风暴沉积不易保存,容易遭到破坏[26]。
在几种主要微体化石的选择上,大多数学者选择有孔虫和硅藻作为研究现代风暴和古风暴的良好类群。有孔虫具有坚硬的钙质外壳,其死亡后壳体易于保存,对海洋环境变化极为敏感[37-38],因此有孔虫可以作为沉积记录的重要载体,对探讨现代和古代风暴沉积均有着重要的作用[39-44]。而硅藻是一种较为常见的单细胞藻类,几乎地球上所有水体都能看见硅藻生长的踪迹[45-47],其主要以浮游和底栖的形式存在于各类水体中,对环境的温度、盐度和营养盐等反应敏感[44,48-49],鉴于其死亡后硅质壳体不易被溶解,易于保存在沉积物中,故而可以作为研究古环境变化[44-45,50-51]和古风暴特征的重要手段之一[32,52]。另一种重要的微体化石是孢粉,具有总体数量大和不易遭受破坏的特点[53],可以用来揭示地质历史时期陆地植被对气候变化的响应,还能通过孢粉研究古气候及古环境变化历史,作为识别古环境演变的重要载体[54-55];但是孢粉受到风暴作用的直接影响有限,且较难分辨地层中孢粉异常层位是否是风暴成因还是其他成因,故其应用有限。总的来说,将有孔虫和硅藻作为风暴研究的指标已经较为成熟。
4 结论和展望
通过对有关风暴沉积微体生物研究的梳理,我们得到以下几点结论:
(1)微体生物是否能记录风暴沉积事件取决于它们是否能很好的保存下来。古风暴记录研究地点的选择变得非常重要,最好的基于微体化石的古风暴重建应来自于有利的沿海环境,这些环境既是低能的,又具有足够的容纳空间,可以沉积和保存沉积物。从以往的研究来看沿海沼泽、潟湖、洼地或者水坑等低能沉积环境都是比较合适的地点,易于保存风暴冲越沉积物,而开阔的潮滩、浅海、河口外部则较难保存这些微体生物记录。
(2)在风暴沉积的识别过程中,一般需要结合地层学和微体化石证据对海岸沉积物序列进行分析,有时微体化石信号比其他的沉积学信号要灵敏。风暴的微体化石特征依赖于对正常原地组合类群中异质性类群的识别,这些异质性类群可以理解为“异地埋藏群”,尤其是在应用有孔虫或硅藻的异质性类群时。
(3)现代风暴微体生物研究虽然已有几十年历史,但是由于风暴过程的复杂性以及形成和保存等问题,仍有待进一步研究。了解现代风暴过程中不同沉积环境下微体生物的沉积特征、面貌以及它们的形成和保存机制是亟待解决的问题。
(4)总的来说,微体生物能灵敏的反映其生存环境,因此它们在搬运和埋藏过程中能很好的记录风暴沉积事件过程。微体生物在研究现代和古风暴沉积方面是一个很好和有效的指标,有时更优于其他沉积学指标。在未来的研究中,还需开展更多不同沿岸沉积环境下的现代风暴研究,并着重于提高古风暴重建的可靠性、准确性和精确性,以及探讨微体化石记录与风暴强度之间的相关关系。
