计算机辅助设计提高疏棉状嗜热丝孢菌脂肪酶热稳定性的研究
2022-08-11万思迪苗华彪韩楠玉
万思迪, 苗华彪, 韩楠玉,2,3
1. 云南师范大学生命科学学院, 云南昆明 650500; 2. 生物能源持续开发利用教育部工程研究中心, 云南昆明 650500; 3. 云南师范大学酶工程重点实验室, 云南昆明 650500
脂肪酶(Lipase,EC 3.1.1.3)广泛存在于微生物、动物和植物中,是一类能够催化酯键断裂和形成的酶[1]。脂肪酶在制革工业、食品加工、医药用品、日用化工等行业都具有良好的应用前景[2]。虽然脂肪酶广泛分布于自然界中,但直至1843年科学家才首次发现兔胰腺中的脂肪酶,随后在1854年和1864年又分别发现了胃脂肪酶和猪胰脂肪酶[3],微生物脂肪酶一直到1901年才被发现[4]。由于微生物具有种类繁多、易于操作、生长周期短、底物专一性强,并且具有较广的pH和温度适应性,因此来源于微生物的脂肪酶已经成为工业生产的重要来源[5]。疏棉状嗜热丝孢菌脂肪酶(Thermomyceslanuginosuslipase,TLL)是一种嗜碱性、底物专一性强且酶活高的脂肪酶。基于TLL的以上特点,诺维信公司开发了可溶性TLL(Lipozyme TL 100 L)和固定化TLL(Lipozyme TL IM),共两种TLL商品酶以供工业生产使用。基于此,TLL在生物柴油的生产、纯对映异构体和区域异构体的生产、聚合物的降解等方面得到了广泛应用[6]。然而,工业生产中往往历经高温条件,天然的TLL会丧失部分酶活,导致生产成本提高,致使TLL的应用受到限制。因此,对TLL耐热性进行改造以满足工业需求极其必要。
目前,可以通过化学修饰[7]、固定化[8]、蛋白质工程[9]、介质工程[10]等多种策略提高蛋白质的热稳定性,其中蛋白质工程主要通过理性设计、半理性设计和非理性设计等方法对蛋白质分子进行改造[11]。理性设计基于对酶的结构、功能和催化活性进行分析,在酶的特定位点上通过定点突变引入特异性的突变[12],通常会借助同源序列比对、晶体结构分析以及计算机辅助设计等手段对酶进行改造。魏子翔[13]等人通过对TLL的盖子(lid)和loop 结构域进行理性设计,获得了一款新型的高活性、耐高温的脂肪酶突变体,并实现了高效表达。姜占宝[14]等人通过定点诱变引入二硫键,致使突变体脂肪酶在60 ℃下耐受20 min后比野生型脂肪酶剩余酶活提高了17.6%。ZHANG[15]等也利用分子动力学模拟的方法提高解脂耶氏酵母脂肪酶的热稳定性,最终获得最适温度提高5 ℃的突变体V213P,且催化活性与野生型相当。温露文[16]等人采用预测软件PoPMuSiC和FoldX计算CALB潜在热稳定性突变位点,利用重叠延伸PCR技术在基因CALB中引入10个单点突变,最终获得比野生型热稳定性提高的3个单突变体A146G、A151P和L278M,对其组合突变得到比野生型最适反应温度均提高了5 ℃的突变体A146G-L278M和A146G-L278M-A151P,Tm值也分别提高了3.3 ℃和4.2 ℃。WANG[17]等也利用计算软件Fold X预测出4个能提高华根霉脂肪酶热稳定性的单突变位点S142A、D217 V、Q239F 和 S250Y,然后对其进行组合产生4倍突变体m31,m31最适温度提高5 ℃、在60 ℃下半衰期为野生型的41.7 倍,表现出增强的热稳定性。王睿[18]基于B-factor和RMSF值为筛选依据以提高脂肪酶分子的热稳定性,选取30个突变体进行验证后发现其中4个突变体的热稳定性得到显著提高。
本研究借助Rosetta旗下自由能计算模块ddg_monomer得出三个突变体(T114W、R175W、R195W)的自由能变化相对野生型TLL小于-5 kcal/mol,预测其为热稳定性可能提高的脂肪酶突变体,并利用定点突变技术成功表达该三个突变体。研究发现其中两个突变体(T114W和R195W)的热稳定性和生物柴油转化率较野生型TLL明显提高。迄今通过计算机辅助设计提高TLL热稳定性的研究较少,该突变体的成功构建为今后研究脂肪酶热稳定性及活性提高提供了参考,得到的TLL突变体T114W和R195W将能很好的应用于工业生产中。
1 材料与方法
1.1 材料
1.1.1菌种来源
TLL连接的pPIC9K载体菌株(pPIC9K-TLL)和大肠杆菌(DH5α)由本实验室保存,毕赤酵母GS115和表达载体pPIC9K均购自于Invitrogen公司。
1.1.2主要培养基
① LB培养基:0.5%酵母粉,1%蛋白胨,1% NaCl,pH自然(约为7),固体培养基在此基础上加2%(w/v)琼脂。② YPD培养基:1%酵母粉,2%蛋白胨,2%葡萄糖,固体培养基在此基础上加2%(w/v)琼脂。③ BMGY培养基:1%酵母粉,2%蛋白胨, 2%甘油,灭菌后超净工作台加入1%生物素,10%YNB溶液,10%磷酸储备液。④ BMMY培养基:1%酵母粉,2%蛋白胨,灭菌后超净工作台加入2%甲醇溶液,1%生物素,10%YNB溶液,10%磷酸储备液。
1.1.3试剂与仪器
棕榈酸对硝基苯酯(pNPP)购于上海麦克林生化科技有限公司,标准品棕榈酸甲酯购于Sigma公司,其余均为实验室常用试剂;基因扩增仪(ETC811,苏州东盛兴业科学仪器有限公司),电转化仪(MicroPulserTm,Bio-Rad),电子恒温水浴锅(DZKW-4,北京中兴伟业仪器有限公司),全波长酶标仪(Thermo Fisher Multiskan,美国),核酸电泳仪(DYC,北京六一),凝胶成像仪(UniversalHoodⅡ,Bio-Rad),蛋白纯化系统(GE Healthcare AKTA Purifier 10)。
1.1.4引物
引物由北京擎科生物科技有限公司合成(表1)。
表1 定点突变PCR产物
1.2 方法
1.2.1计算机辅助设计方法
TLL晶体结构(PDB ID:1DT3)[19]的单体(共269个氨基酸)用于自由能变化的计算,蛋白质稳定性预测工具Rosetta ddg_monomer[20]用于计算TLL所有单位点突变体(共计5 111个)与野生型之间的自由能变化差值(ΔΔG)。首先,对TLL单体的三维结构进行能量最小化计算;能量最小化完成后执行循环次数设置为50次Rosetta ddg_monomer程序,具体的命令如下(/path/to/ddg_monomer.linuxgccrelease-in:file:s TLL.pdb-ddg::mut_file each_mutant.mutfile-ddg:weight_file soft_rep_design-database /usr/local/rosetta/main/database/-fa_max_dis 9.0-ddg:minimization_scorefunction talaris2013-ddg::iterations 50-ddg::dump_pdbs true-ignore_unrecognized_res-ddg::local_opt_only false-ddg::min_cst true-constraints::cst_file input.cst-ddg::suppress_checkpointing true-in::file::fullatom-ddg::mean false-ddg::min true-ddg::sc_min_only false-ddg::ramp_repulsive true-unmute core.optimization.LineMinimizer-ddg::output_silent)。结果统计发现,只有三个突变体的ΔΔG<-5 kcal/mol(T114W、R175W、R195W),预测其为热稳定性可能提高的脂肪酶突变体。
1.2.2定点突变和基因克隆
在定点突变实验中,利用pPIC9K-TLL作为模板构建了三种脂肪酶单突变体T114W、R175W、R195W(图1A),定点突变PCR方法均参照定点突变试剂盒说明书进行。然后将5 μL PCR产物转至50 μL的DH5α感受态细胞中,冰浴半小时后于42 ℃水浴锅热激1 min,在超净工作台内加入提前灭好菌的200 μL LB培养基,放入摇床进行孵育,最后将培养获得的菌体涂于固体LB培养基上过夜培养。次日,挑取长势良好的单菌落进行阳性克隆验证后(图1B),再将验证成功的单菌落送至测序公司测序验证。
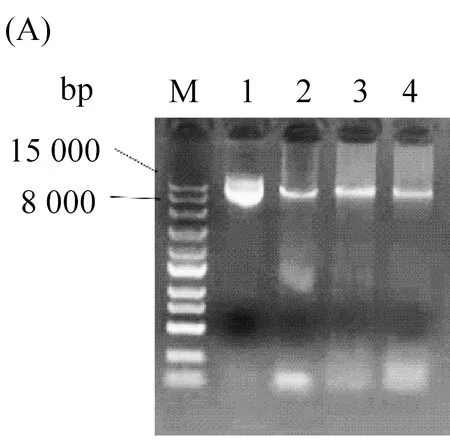
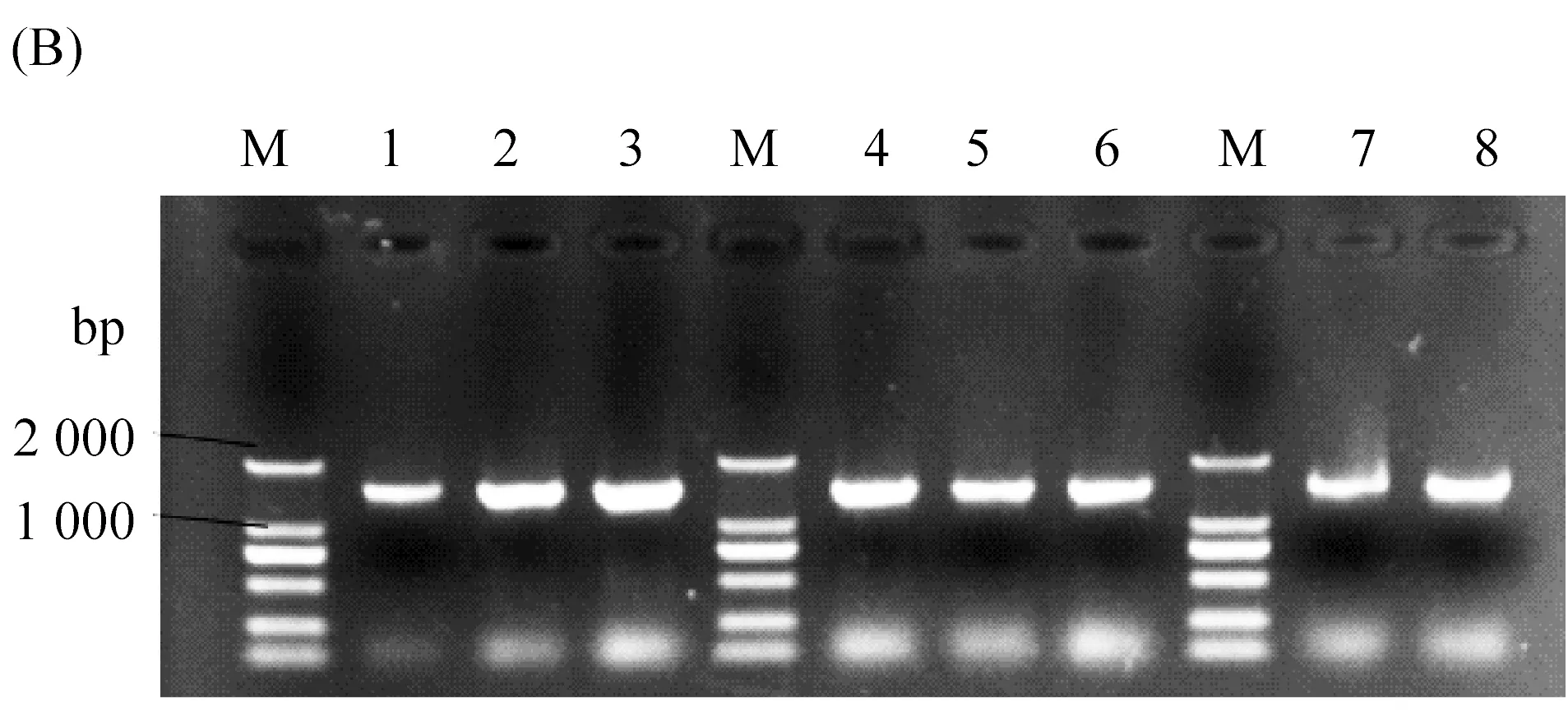
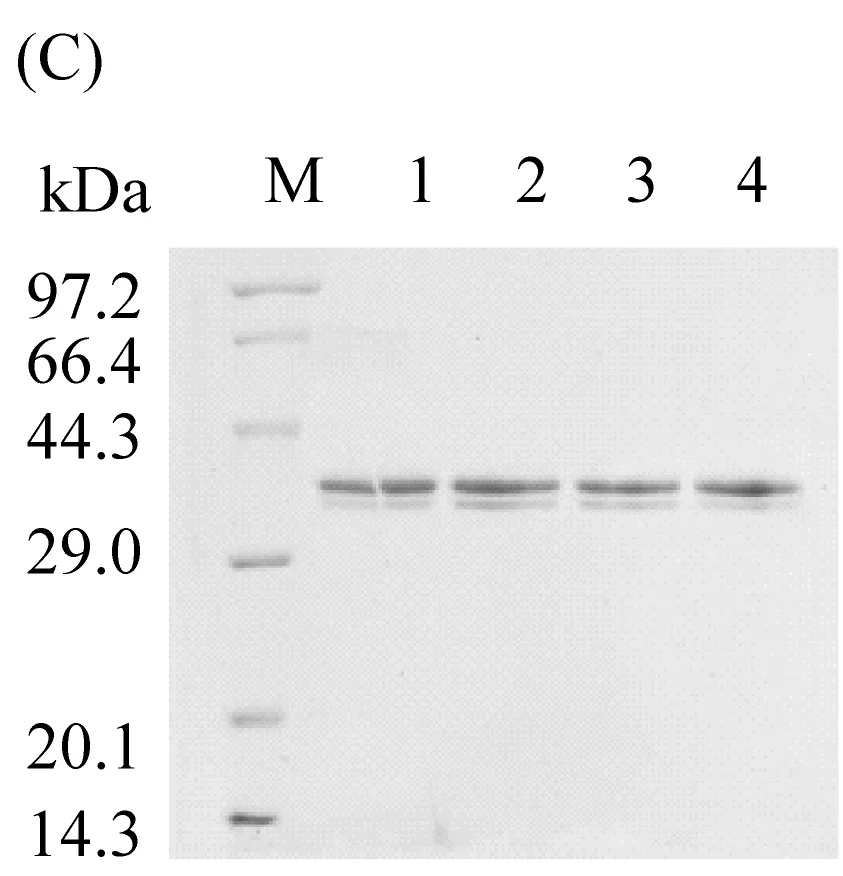
图1 电泳图(A)为野生型TLL及突变体T114W、R175W、R195W的突变条带,M为15 000 bp的Marker,泳道1:野生型TLL,泳道2:T114W,泳道3:R175W,泳道4:R195W。(B)为突变体T114W、R175W、R195W阳性克隆验证成功的条带,M为2000 bp的Marker,泳道1、2、3为T114W阳性克隆验证成功的条带,泳道4、5、6为R175W阳性克隆验证成功的条带,泳道7、8为R195W阳性克隆验证成功的条带。(C)为野生型TLL及突变体T114W、R175W、R195W的蛋白胶图,M为97.2 kDa的Marker,泳道1:野生型TLL,泳道2:T114W,泳道3:R175W,泳道4:R195W。
1.2.3蛋白表达与纯化
① 电转化:用限制性核酸内切酶SalI 线性化野生型pPIC9K-TLL和三种脂肪酶单突变体T114W、R175W、R195W,之后电转化入毕赤酵母GS115菌株中;② 菌株培养:将菌液涂至含有山梨醇的YPD固体培养基上,在30 ℃恒温培养箱培养48 h,之后用含有遗传霉素的YPD固体培养基筛选出至少30个克隆子,再转至YPD固体培养基上30 ℃培养48 h;③ 发酵:各挑取YPD固体培养基上的菌株于装有2 mL已灭菌BMGY培养基的试管中,30 ℃、200 r/min 振荡培养48 h,到超净工作台取出菌液,8 000 r/min离心10 min收集菌体,再将菌体转至装有2 mL已灭菌BMMY培养基的试管中,30 ℃、200 r/min 振荡培养48 h获得发酵液;④ 纯化:将获得的上清液加入Ni2+-NTA亲和层析柱,用含有不同浓度的咪唑洗脱液进行洗脱,最终用SDS-PAGE确认纯化结果(图1C)。
1.2.4酶活力测定
取适度稀释的酶液50 μL加至已经在37 ℃电子恒温水浴锅中预热2 min的450 μL底物溶液中,反应5 min,然后加入50 μL 10%的SDS终止反应,对照则在其它反应条件皆相同的情况下先加入SDS再加入酶液,最后皆加入500 μL 0.1 mmol/L的碳酸钠溶液。各吸取200 μL反应液和对照液在405 nm下测定吸光值。根据pNP标准曲线y=4.051x+0.011(R2=0.999 8)算出pNP的浓度,带入脂肪酶酶活力计算公式算出其酶活力。棕榈酸对硝基苯酯(pNPP)测定酶活的原理:为每分钟水解底物pNPP生成1umol对硝基苯酚所需要的酶量定义为一个酶活单位(1 U)[21]。
1.2.5最适pH、温度和 pH、温度稳定性测定
对野生型脂肪酶pPIC9K-TLL和三种脂肪酶单突变体T114W、R175W、R195W进行酶学性质的分析,最适pH测定:取适当稀释过的酶液在37 ℃、不同pH(4~12)下进行酶促反应测定最适pH;最适温度测定:取适当稀释过的酶液在最适pH时分别在30 ℃、40 ℃、45 ℃、50 ℃、55 ℃、60 ℃、70 ℃和80 ℃进行酶促反应测定最适温度;pH稳定性测定:用不同pH(4~12)的缓冲液适当稀释后的酶液在37 ℃下耐受1 h,之后在最适pH 、37 ℃下进行酶促反应,以未处理的稀释酶液作为 100%;温度稳定性测定:取适当稀释过的酶液在最适pH时分别在70 ℃下耐受1 min、3 min、5 min、7 min、10 min、15 min、20 min、25 min、30 min、45 min、60 min、75 min和90 min进行酶促反应测定温度稳定性;取适当稀释过的酶液在最适pH时分别在80 ℃下耐受1 min、3 min、5 min、7 min、10 min、15 min、20 min、25 min、30 min、45 min、60 min、75 min和90 min进行酶促反应测定温度稳定性。
1.2.6动力学测定
在37 ℃、pH为9.0时,以0.006 mmol/L 、0.012 mmol/L、0.030 mmol/L、0.045 mmol/L、0.060 mmol/L、0.120 mmol/L 、0.180 mmol/L、0.240 mmol/L、0.360 mmol/L和0.480 mmol/L浓度的pNPP进行酶促反应,利用GraphPad Prism 5的酶动力学(Enzyme kinetics)中的米氏方程(Michaelis-Menten)计算动力学参数Km值、Vmax值、kcat值。
1.2.7生物柴油转酯率测定及统计分析
先制备野生型脂肪酶及突变体催化酯交换反应制备生物柴油的反应体系,称取50 g已经除过杂质的地沟油置于100 mL锥形瓶内,加入6 000 U酶活单位的脂肪酶(野生型和突变体),再加入7.5 mL甲醇、2.5 mL正己烷、35 μL Span60和5 mL的水。配置完成之后将锥形瓶置于37 ℃、200 r/min 振荡培养箱中反应8 h,反应结束后置于80 ℃电热鼓风干燥箱灭活30 min,12 000 r/min离心10 min取上层油酯相测定酯交换反应所得脂肪酸甲酯的含量。然后再吸取1 mL地沟油于50 mL锥形瓶内,加入10 mL 10%盐酸-甲醇溶液、5 mL甲苯、1 mL饱和NaCl溶液,于50 ℃水浴反应8 h结束后,加入5 mL正己烷混匀,5 000 r/min离心3 min取上层有机相测定地沟油完全甲酯化后脂肪酸甲酯的含量。
对TLL酶比活力与脂肪酶突变体酶比活力的差异、TLL转酯率与脂肪酶突变体转酯率的差异,分别用独立样本t检验(independent-Sample T Tests)进行了比较,所有数据用SPSS 25.0 统计软件包进行比较分析。
2 结果与分析
2.1 序列分析
TLL序列全长825 bp,共编码275(含6个His标签)个氨基酸残基,蛋白分子量为29.9 kDa。通过纯化后的SDS-PAGE分析证明(图1C),TLL及突变体T114W、R175W、R195W分子量一致, 活性分别为311 U/mg 、191 U/mg、3 U/mg、262 U/mg。
2.2 TLL及突变体T114W、R175W和R195W的最适温度及温度稳定性
野生型脂肪酶TLL的最适温度为40 ℃;突变体T114W和突变体R195W的最适温度为40 ℃与野生型相同;然而,突变体R175W的最适温度为35 ℃,较野生型降低了5 ℃(图2A)。将野生型脂肪酶TLL及突变体T114W、R175W、R195W置于70 ℃和80 ℃下进行耐受处理(图2B、2C),具体表现为:在70 ℃下,野生型TLL处理90 min相对剩余酶活下降到44.49%,T114W、R175W和R195W在相同条件下相对剩余酶活下降到57.95%、39.63%和67.37%。温度耐受结果表明相比野生型TLL,突变体T114W和R195W在70 ℃下耐受90 min后相对剩余酶活分别提高了13.46%和22.88%,耐热性相比TLL有所提高。在80 ℃下,野生型TLL耐受30 min后相对剩余酶活降到47.03%,T114W、R175W和R195W在相同条件下相对剩余酶活下降到59.26%、11.88%和21.72%。温度耐受结果显示突变体T114W在80 ℃下的热稳定性相比TLL有所提高,相对剩余酶活提高了17.26%。
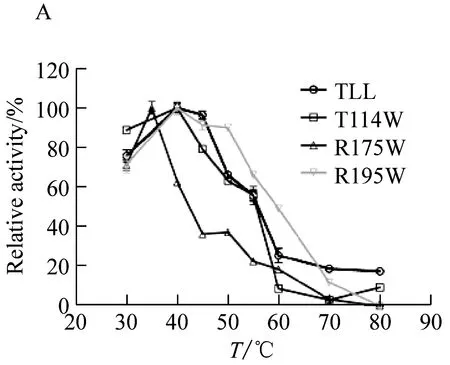
图A为最适温度
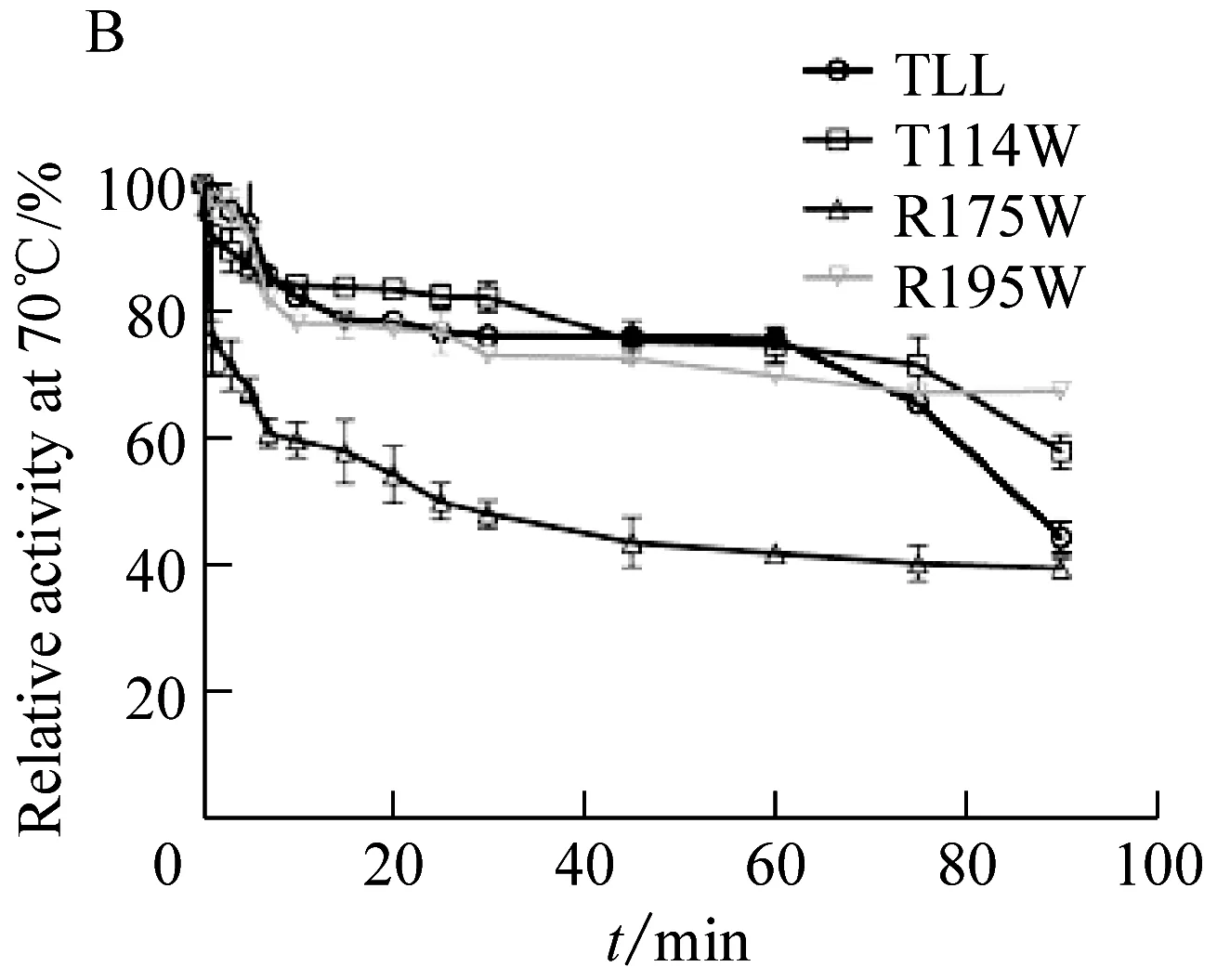
图B为在70 ℃下温度稳定性
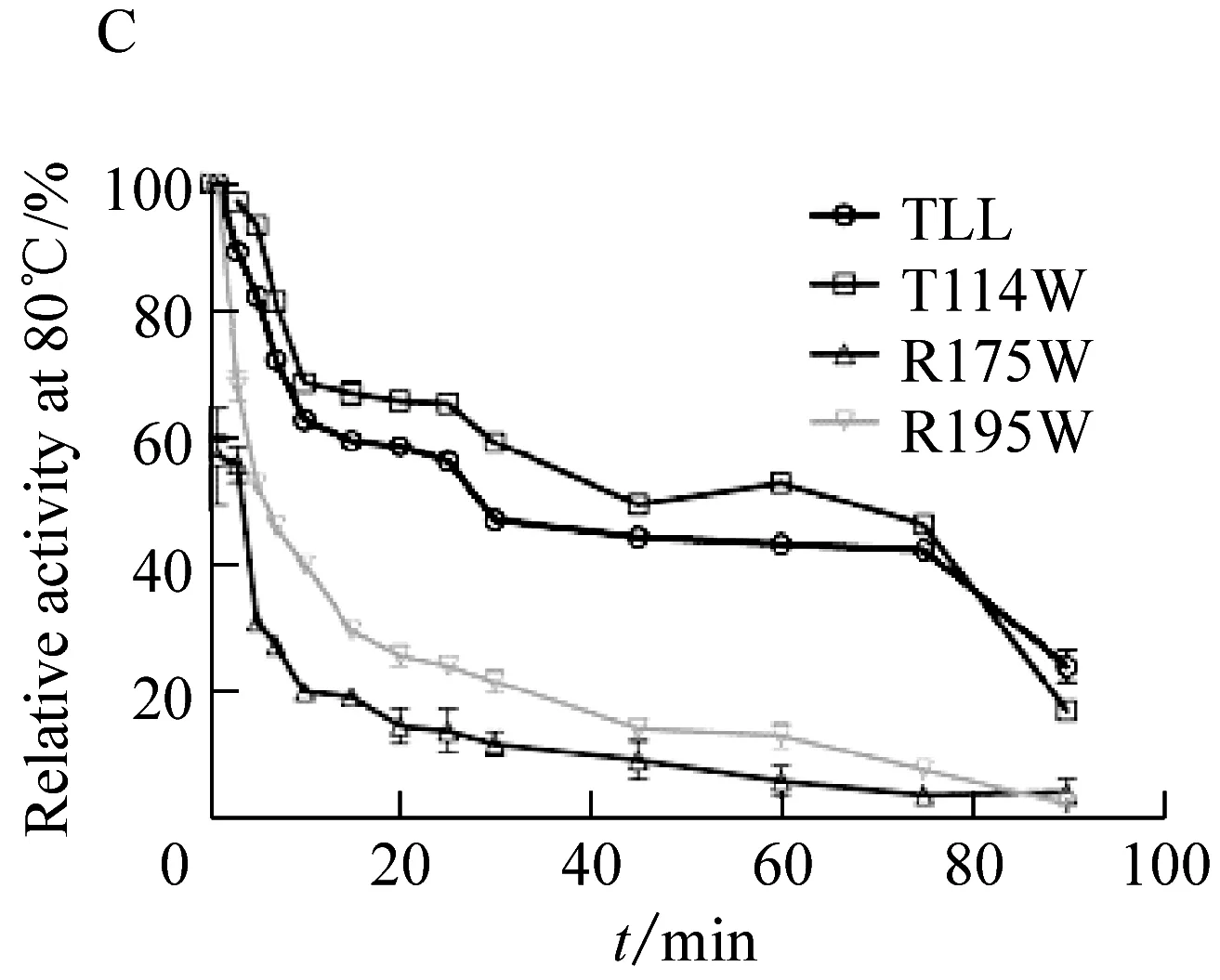
图C为在80 ℃下温度稳定性图2 TLL及突变体T114W、R175W、R195W温度图
2.3 TLL及突变体T114W、R175W、R195W的最适pH及pH稳定性
脂肪酶TLL与三个突变体的最适pH均为9.0,偏碱性(图3A)。对野生型和突变体在37 ℃、pH 4.0~12.0下处理1 h,发现耐热性提高的突变体T114W和R195W在pH 4.0~12.0的相对剩余酶活也均高于野生型,提高了10%~15%。而突变体R175W相比于野生型TLL在pH 3.0~10.0的pH耐受性无明显变化(图3B)。表明突变体T114W和R195W比野生型TLL的pH稳定性有所提高。
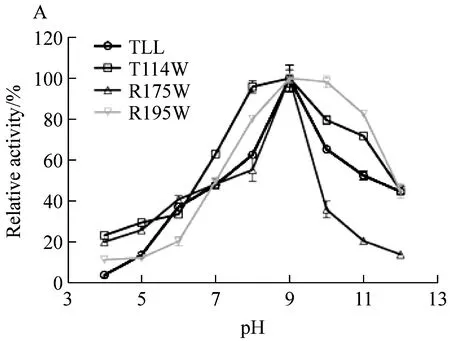
图A为最适pH
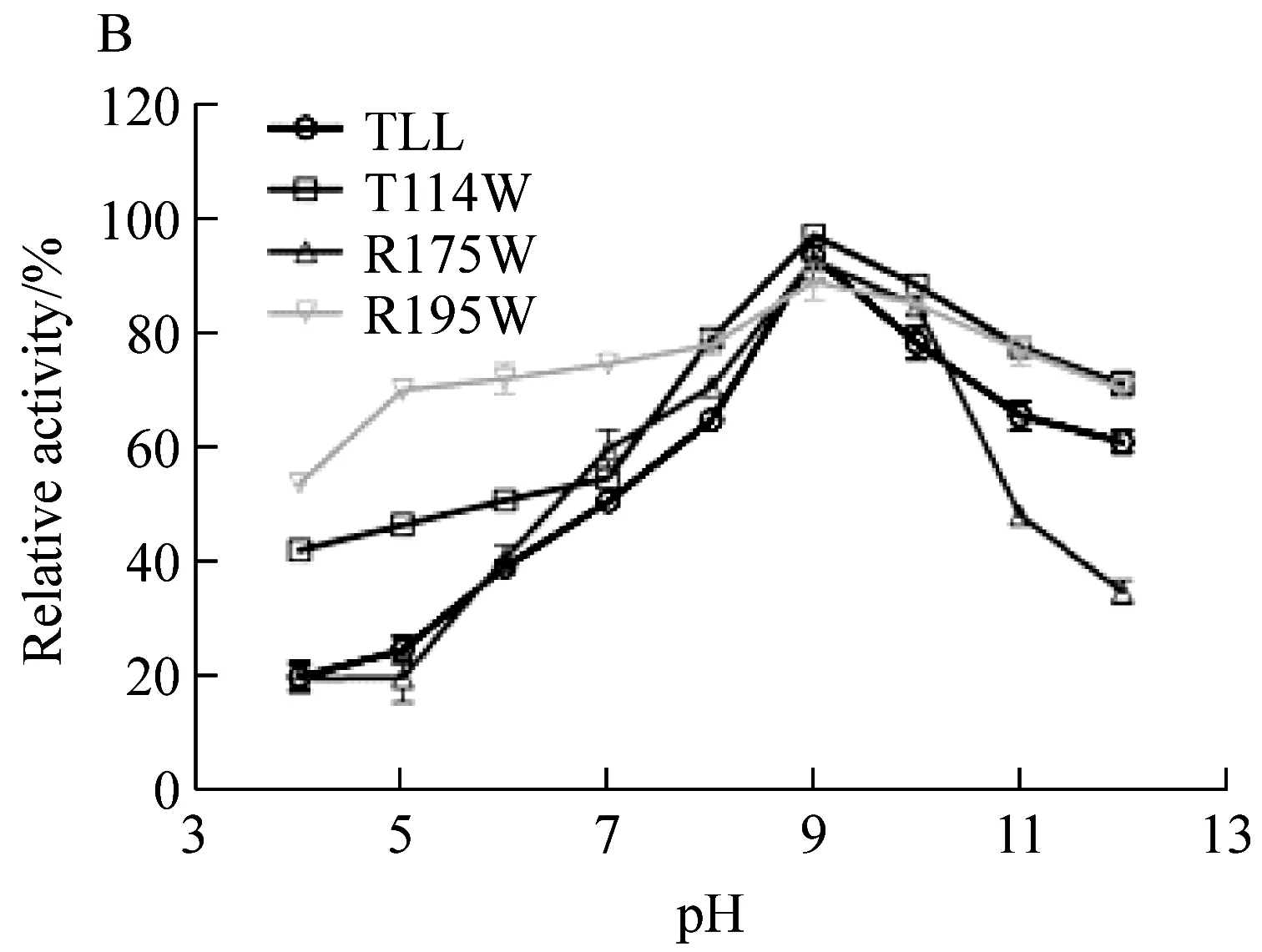
图B为pH稳定性图3 TLL及突变体T114W、R175W、R195W的pH图
2.4 TLL及突变体T114W、R175W、R195W的动力学分析
TLL及突变体T114W、R175W、R195W的酶促反应动力学参数见表2,T114W和R195W的Km与野生型相比有所下降,Km下降表明这两个突变体对底物的亲和力变大。而R175W的Km比野生型大则表明该突变体对底物亲和力相对较弱,联系到R175W的酶活性仅有3 U/mg,也间接表明了该突变体在酯水解反应中的催化作用极差。R195W的kcat/Km值比TLL的有所提高,则表明该突变体在催化速率方面有所升高,然而其余两个突变体在催化速率方面并未变化。综上表明,突变体R195W有更高的动力学和催化效率。
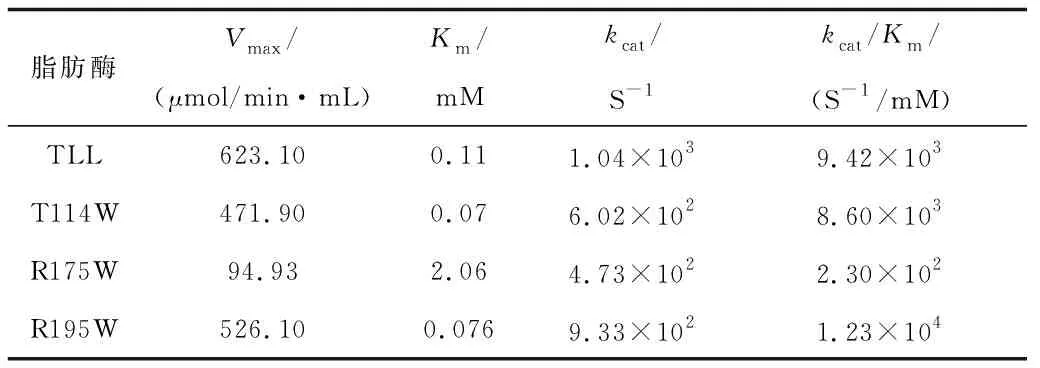
表2 TLL、T114W、R175W、R195W动力学参数
2.5 TLL及突变体T114W、R175W、R195W的生物柴油转酯率分析
经测定,野生型脂肪酶TLL及突变体T114W、R175W、R195W的转酯率分别为49.70%、64.82%、25.13%和58.20%,突变体T114W和R195W比野生型分别提高了15.12%和8.50%。对TLL及突变体的酶比活力和生物柴油转酯率进行了相关性分析,图4表明脂肪酶的酶比活力和生物柴油转酯率呈显著正相关(r=0.793,P=0.002),随着脂肪酶的酶比活力增高其转酯率也相应提高。
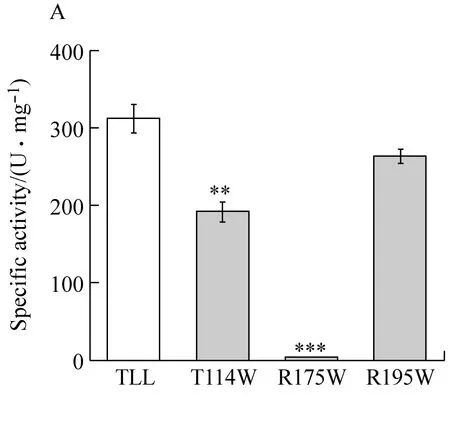
图A为酶比活力
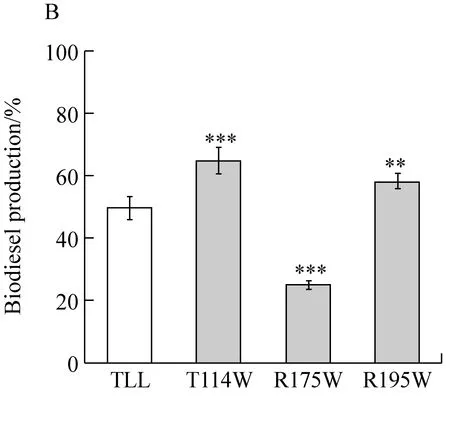
图B为转酯率
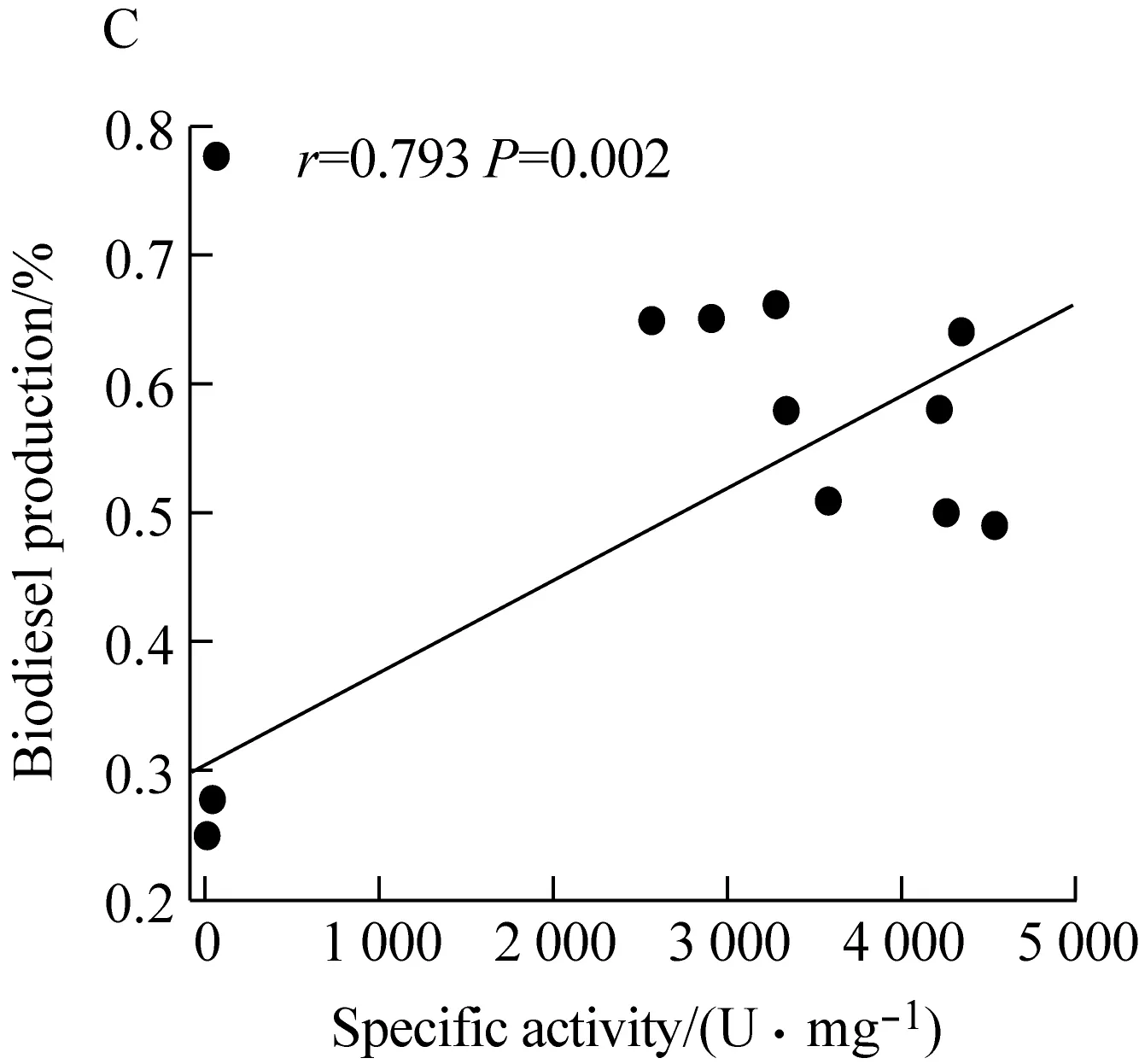
图C为二者相关性分析
3 讨论
本研究借助Rosetta旗下自由能计算模块ddg_monomer预测出三个突变体(T114W、R175W、R195W)的热稳定性相比于野生型TLL可能提高,并利用定点突变技术成功表达该三个突变体。实验发现其中两个突变体(T114W和R195W)的热稳定性和转酯率较野生型TLL明显提高,而R175W由于突变位置导致其酶活几乎丧失。Rosetta ddg_monomer预测的三个突变体均为将原有序列突变为色氨酸,对原始结构进行分析发现,突变位点T114和R195虽然均为极性氨基酸,但其并未和任何周围氨基酸形成氢键或者盐桥类的相互作用,当将这两个位点突变为色氨酸(Trp,W)时,位于α螺旋上的W114很有可能和相距0.3 nm的loop结构形成cation-π的相互作用;而当将195位的精氨酸(Arg,R)突变位色氨酸时,位于β折叠上的W195可以与H215的环状侧链形成稳定的π-π相互作用,这两种新增范德华力的形成,导致了TLL突变体结构更加稳定,且并不影响其活性,基于位点114和195的色氨酸突变体最终使得TLL的热稳定性、生物柴油转化率得以大幅度提高(图5)。这也说明了芳香类氨基酸可以通过cation-π或者π-π相互作用使TLL脂肪酶的三维结构更加稳定,后续对TLL的进一步改造提供了指导意义。
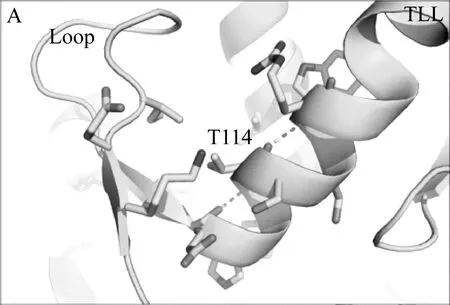
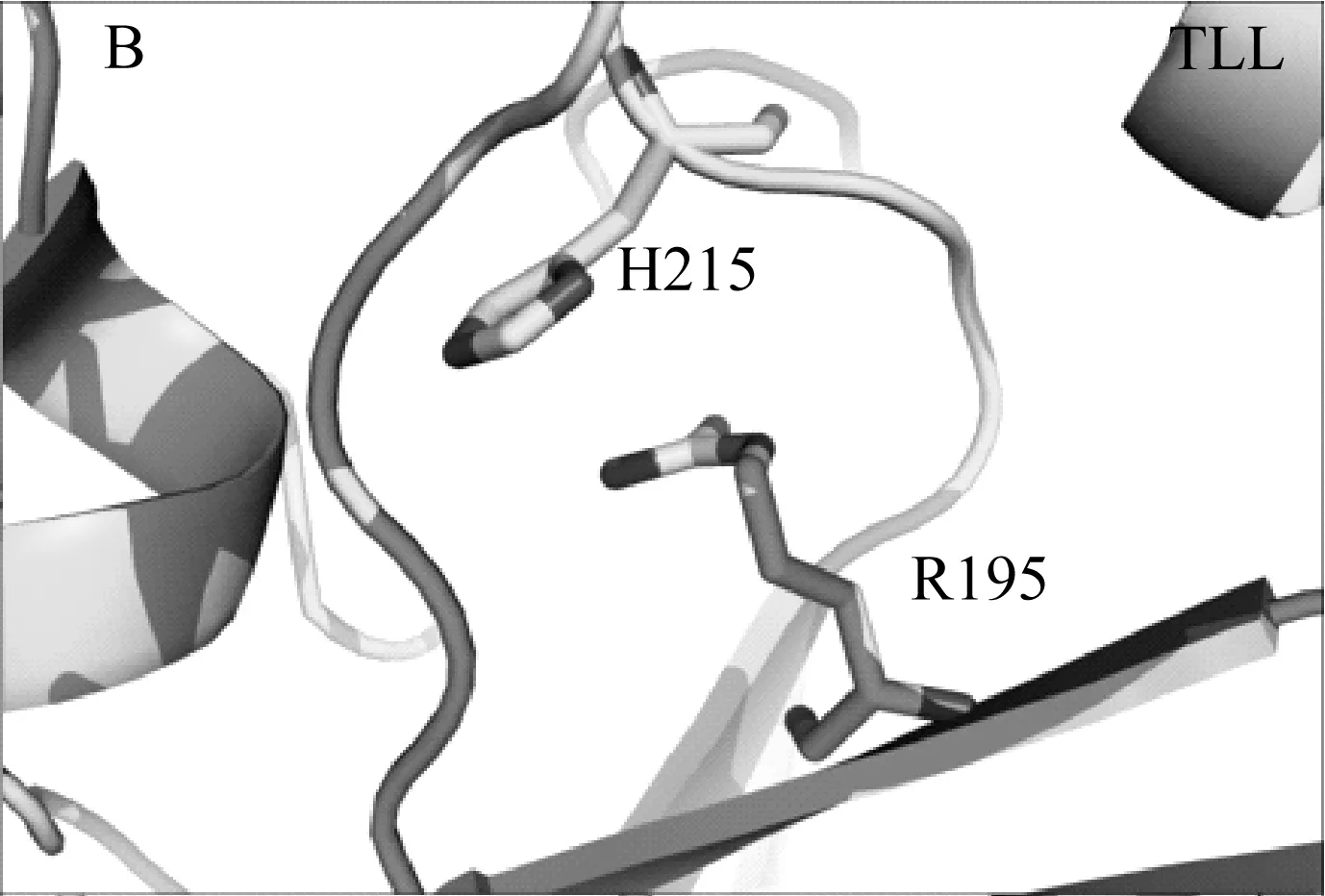
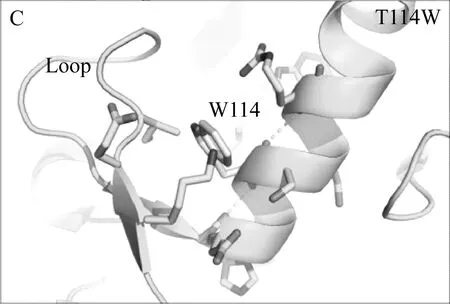
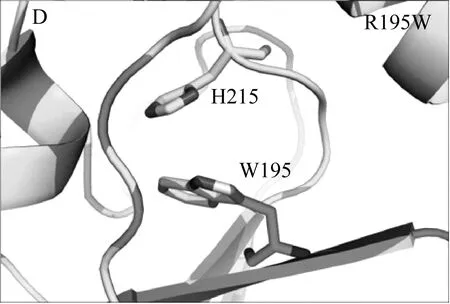
图5 TLL、T114W和R195W的3D结构比较
目前在优化TLL耐热性的研究中有很多是对其三维结构进行评估,替换不稳定的“盖子”或 loop区域氨基酸,得到耐热性提高的突变体酶[13]。除此之外,利用化学法或者物理法将脂肪酶制备成不溶于水但仍具备催化活性的固定化复合酶,来提高酶的热稳定性。但是该方法存在一些限制,工业条件下需要考虑到酶的稳定性、活性、纯度和对底物的选择性、特异性等问题,以及如何从反应介质中分离出高活性的酶,增加了工艺成本[22]。还有通过化学修饰,如增加糖基化位点等得到耐热性提高的突变体酶[23]。文献调研发现利用理性设计手段优化 TLL 耐热性的研究十分稀少,针对 TLL 耐热性的改造研究全部集中在固定化的技术革新上,本研究利用 Rosetta 软件对TLL及其突变体进行ΔΔG计算,通过对计算结果进行分析和筛选得到热稳定性提高的两个突变体酶,与以往提高TLL热稳定性的方法相比更加精准和有效。
此外,本研究还发现脂肪酶的水解活性与其生物柴油转化率呈显著正相关。由于脂肪酶的活性测定是测定其水解脂肪的效率属于水解反应,地沟油或其它脂质转化为生物柴油虽然为酯交换反应,但其催化途径类似,均是通过脂肪酶的催化作用将水或者醇类物质的羟基与氢离子或碳链部分通过亲核取代反应分离,最终达到生成甘油和脂肪酸或生物柴油的目的。因此,当对脂肪酶进行改造时,如果发现其催化活性有所提高,也间接证明其生物柴油转化率可能会有所升高。
综上,本研究证明利用计算机辅助设计软件提供潜在耐热TLL突变体的研究策略是有效而且可靠的,研究产生的耐热突变体T114W和R195W也能够用于极端条件下的工业生产中,且该研究为提高工业用酶耐热性提供了参考依据。
