水稻长链非编码RNA的研究进展
2022-08-11于雪然薛欣月杜怀东李培富马天利
于雪然,薛欣月,田 颖,杜怀东,李培富,马天利
(宁夏优势特色作物现代分子育种重点实验室/宁夏大学农学院,宁夏银川 750021)
1992年首次在人类中发现了长链非编码RNA(lncRNAs)。它最初被认为是一种不具备功能的“转录噪音”,是一段不具备编码功能的RNA序列。随着研究的不断深入,人们逐渐意识到lncRNAs在基因的表达调节中起着重要的作用。1999年,古井宏等在对水稻和转基因大豆的表达分析中也发现了lncRNA,这是一个与水稻根瘤菌形成有关的lncRNA-。通过表达分析表明,水稻与豆科植物具有共同的特性,并且可能在茎维管束的分化过程中起作用。
当前,水稻作为主要粮食作物之一,是世界上大部分人口的主食。已有研究表明,水稻中lncRNAs不仅参与水稻多个生长发育过程,而且在水稻参与非生物逆境反应及病害防御等过程中也发挥重要功能。虽然近年来植物lncRNAs的研究取得了一些进展,但是总体而言,水稻lncRNAs的研究还不够深入。本文就近年来水稻中已发现的lncRNAs的种类、参与的生物学过程、发挥的作用机制及与水稻相关的lncRNAs生物信息数据库进行综述及展望,期望能为更深入地了解水稻中的lncRNAs提供借鉴。
1 水稻lncRNAs的相关数据库
人们在研究核糖核酸的过程中发现了许多形状大小各不相同的非编码RNAs。非编码RNAs基因的进化与蛋白质编码基因的进化有所不同,在大多数情况下,蛋白质编码基因都是通过部分复制和后续序列的差异过程产生的。lncRNAs的进化主要是由于从影响染色质调节、增强子功能或其他过程的各个启动子中选择转录而来,且随时间流逝可以获得调控因子的结合位点。lncRNAs的形成大致通过以下5种方式完成:(1)一个蛋白质编码基因框架被破坏,从而突变转化成为一个功能性非编码RNA,同时整合了转座因子序列。(2)随着染色体重排后,2个未转录的序列区域和先前分离良好的序列并置,产生1个多外显子非编码RNA。(3)通过反向转座复制非编码基因,产生功能性非编码反向基因或非功能性非编码反向假基因。(4)非编码RNA中的相邻重复是由2个串联重复结构产生的。(5)功能性非编码RNA来源于转座因子的插入或分离。
随着植物中的lncRNAs不断被发现和深入研究,与植物lncRNAs相关的生物信息数据库也相继出现,如LncRNAdb、PLncdb、PlantNATsDB等。已知lncRNAs在水稻生长过程中对部分基因的表达起调节作用,但具体的作用机制和调控途径研究有限,特别是在生物技术、农业和基础研究等方面具有重要作用的lncRNAs仍缺乏全面的鉴定和功能注释。下边列举主要和水稻相关的数据库以期为后续的研究提供有效依据(表1)。

表1 水稻lncRNAs相关数据库
CANTATAdb数据库由最初的10种模式物种扩展到现在的39个物种,收录了大部分植物的lncRNAs信息,数量高达239 631种lncRNAs,是植物最大的lncRNAs数据库。其中包括已知的水稻lncRNAs,共38 529种,转录数达450 170个。
GreeNC数据库收录了49个物种的lncRNAs,其中包括37种植物和6种藻类,总lncRNAs超过120 000个,具有不少于20万条带注释的转录本。其中与水稻有关的lncRNAs有 4 495 个。
PLNlncRbase涵盖了43种植物的lncRNAs,共 1 187 个。具有可以使用植物种类名称或lncRNAs标识符搜索,以及从界面中检索植物lncRNAs条目等优点。
EVLncRNA是经过低通量实验验证的lncRNAs数据库,包括lncRNAdb、LncRANDisease、Lnc2Cancer和PLNIncRBase,以及其他先前未涵盖的功能和特定信息所建立的lncRNAs,共1 543个。其中有428个lncRNAs是来自44个植物物种的,这之中也包括部分水稻的lncRNAs。
PNRD是植物的微小核糖核酸(miRNA)数据库,满足了对植物miRNA研究的需求。充分利用大量公共资源的方法,为植物非编码RNA(ncRNA)的基础研究提供了便利。目前该数据库已从150种植物中收集了11类不同类型的ncRNA,共计25 739种。包括 lncRNAs、tRNA、rRNA、tasiRNA、snRNA 和 snoRNA 等。其中已经验证的水稻lncRNAs有38个,未预测具体功能的水稻lncRNAs有1个。
PLncPRO是一个利用转录组数据,使用随机森林算法对编码和长非编码数据进行分类,从而预测植物中非编码基因的数据库。与其他现有工具相比,PLncPRO具有更好的预测精度。在干旱或盐胁迫条件下,利用PLncPRO在水稻中发现了3 714个高置信度lncRNAs。
IC4R-2.0包含3 215个lncRNA位点和6 259个转录本,但没有相关的多组特征。该数据库,在水稻基因组中系统地表征了长链非编码 RNA(lncRNA)和环状 RNA(circRNA)。性能评估结果表明,与之前的注释系统相比,IC4R-2.0 实现了更高的完整性和质量,这主要归功于应用了大量的基因组注释RNA-Seq 数据。
PLncDB V2.0根据13 834个RNA-Seq数据集整理得到了80种植物的1 246 372个lncRNAs,这其中包含从98个RNA-Seq文库中鉴定的 11 565 个水稻lncRNAs信息。PLncDB V2.0整合来自其他4个资源(包括EVLncRNA、RNAcentral等)的lncRNAs信息,界面友好,具有很多特点,例如可以使用多种工具可视化lncRNA表达模式和表观遗传信号,具有高达5个内置搜索引擎用于lncRNAs的研究,还提供了lncRNAs的可能靶标和调控网络。
RiceLncPedia是一个精选的水稻 lncRNAs 数据库,包括2 312个公开可用的RNA-Seq文库,6 965个水稻 lncRNAs。并按以下方式分为5类:(1)lncRNAs在不同组织、发育阶段和胁迫处理中的表达谱;(2)lncRNAs与基因组变异的关联;(3)lncRNAs与表型的联系;(4)lncRNAs和转座子元件的重叠信息;(5)预测为miRNAs靶标或miRNA前体的lncRNAs。
2 lncRNAs在水稻生长发育中的作用
随着全基因组芯片及高通量测序数据的不断积累,研究发现在现有的物种中,大部分基因组是可以转录的,但只有大约2%的基因可以编码基因组。近年来,对基因的特异性和高通量研究发现,水稻中的一些lncRNAs参与多种生物学功能的调控过程,可以与蛋白、DNA和RNA相互作用,以RNA的形式在多种层面上参与调节水稻的分子生物学功能,包括水稻的生殖生长、抗逆及胁迫过程等。研究证明,lncRNAs作为一种生物调节因子参与了水稻的生命调节过程,在水稻不同的生长发育过程中具有特异性,在水稻的生长发育过程中起到了非常重要的作用。
2.1 非生物胁迫
非生物胁迫是水稻生长发育中常见的抑制因素。水稻需要优化其生长和代谢过程,来适应环境的变化。研究发现,水稻中的非生物胁迫,如低温、高温、干旱、盐碱、重金属等均与lncRNAs有关(图1)。许多表达下调多聚腺苷酸化(DPA)的lncRNAs通过参与激活多组应激反应基因的方式,影响水稻脱落酸的生物合成、转运和代谢过程。当水稻所处环境温度过高或过低时,一些DPA lncRNAs通过负调控相关酶活性的方式参与水稻的抗逆过程。Shin等发现1种lncRNA()在冷处理条件下,参与了幼苗的应激反应。Leng等在鉴定水稻孕穗期的冷敏感lncRNAs时,共得到1 485个lncRNAs,其中有566个被发现有差异表达。Luo等研究发现,暴露于高温胁迫下的水稻受精10 d后,在收获的穗中鉴定得到了578个lncRNAs,其中包括14个显著上调表达基因及45个显著下调表达基因。在这59个差异表达的基因中,有32个被预测到参与了淀粉的代谢和分解过程,表明这些基因参与胚乳淀粉的形成,从而进一步影响垩白的形成。Qi等研究发现,在干旱胁迫下,鉴定得到 1 092 个与lncRNAs有关的差异表达基因,该结果为水稻品种抗旱性的遗传改良提供了候选调控因子。Li等在水稻干旱反应研究过程中发现,有 6 885 个转录本和238个lncRNAs参与其中。作为的前体,调节关键基因,通过参与脱落酸信号传导的方式,调节水稻短期干旱记忆的过程。此外,Yuan等发现,一些与干旱有关的DPA lncRNAs参与了细胞壁的加厚和脱落酸的转运过程。Singh等在对水稻盐胁迫的研究中发现,水稻中有3 714个高置信度的lncRNAs参与该过程,其中有27.2%的lncRNAs至少在1种胁迫中差异表达。
目前已有转录组学相关研究表明,水稻中存在大量的lncRNAs,且此类lncRNAs在水稻细胞对氮磷的反应、磷饥饿和非生物胁迫等相关通路中均有注释。Shin等发现,在氮磷缺乏的情况下,的表达发生改变。这表明,可能参与了氮磷平衡的调控。在氮磷饥饿条件下,在水稻根中表达水平增加。在缺氮根中,表现出对氮饥饿反应的诱导表达模式。
许多lncRNAs在镉、砷和铁等金属胁迫下发挥作用。镉、砷等金属是植物生长发育过程中的非必需元素,日益加剧的镉、砷污染与水稻生产有直接关系,其在水稻中的可食用部分大量增加,严重影响了人类的健康。Chen等利用基因组测序技术发现,lncRNAs可以通过选择性剪接及标靶模拟物的方式参与控制水稻的镉代谢过程,使与半胱氨酸和甲硫氨酸代谢途经相关的基因发生显著变化。通过这些研究可知,在镉胁迫下,lncRNAs通过顺式调节富含半胱氨酸的代谢基因,反式调节光合作用和次生代谢物来减轻镉的毒性,从而激活各种生理生化反应。Tang等在研究水稻对砷胁迫的lncRNAs的响应中发现了56个lncRNAs,表明lncRNAs参与了砷胁迫的信号传递。
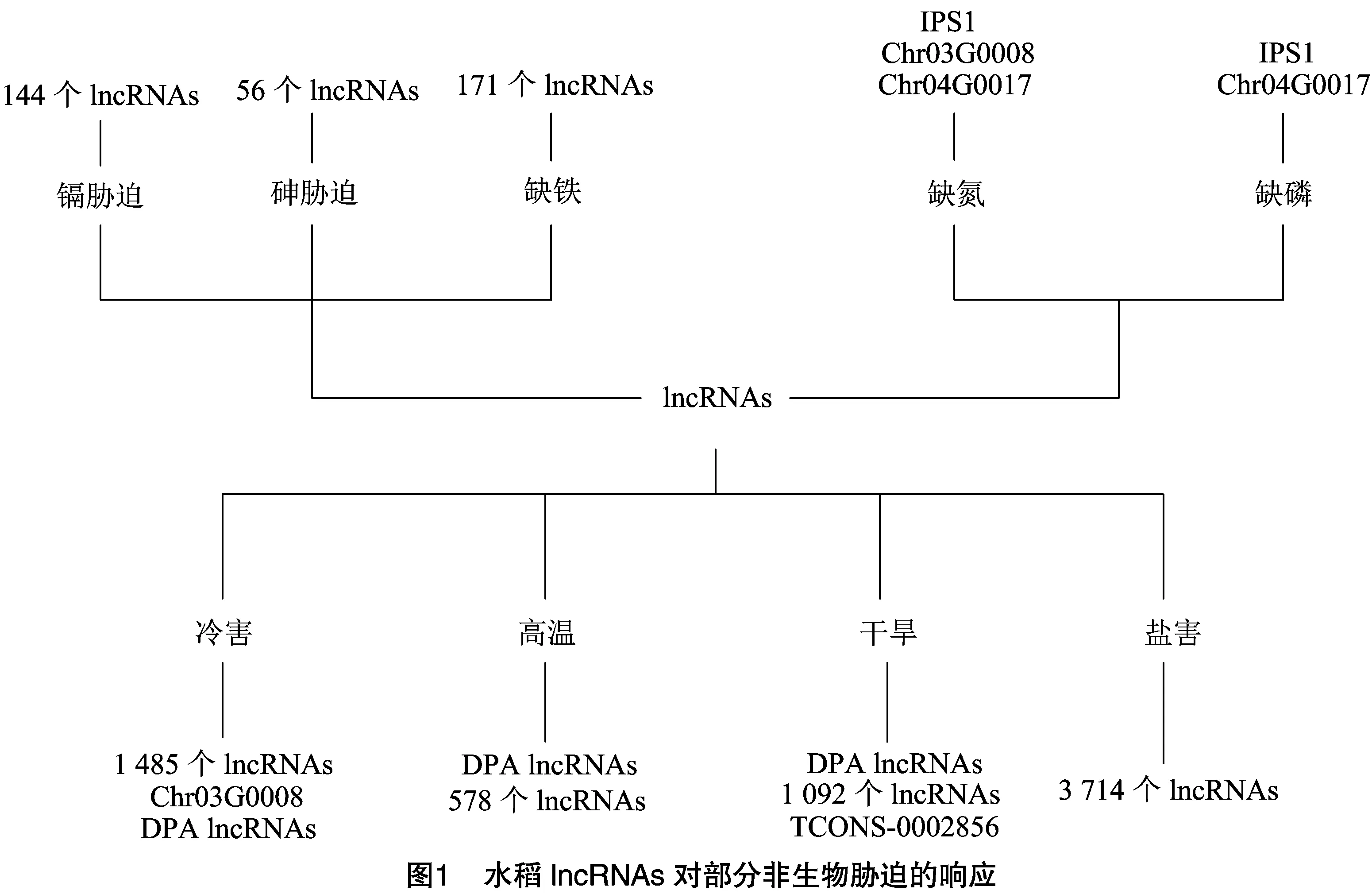
铁缺乏会直接影响水稻的生长发育,导致水稻产量和品质下降。Wang等发现,有171个lncRNAs在缺铁条件下在水稻茎和根中被鉴定出来,其中有数个lncRNAs可以作为miRNAs或内源性标靶模拟物来调节铁相关基因的表达。许多lncRNAs参与了水稻的铁转运过程及光合作用,表明lncRNAs在调节铁稳态中起作用。
2.2 生物胁迫
生物胁迫在水稻生产中存在着严重的风险。为了应对这一系列风险,水稻已经进化出一套防御机制。近年来研究发现,lncRNAs是水稻应对胁迫机制的重要组成部分(图2)。
水稻是全世界最主要的谷类作物之一,但在许多地区,由于稻瘟病的影响,水稻产量出现了严重损失。Jain等在研究水稻接种稻瘟病病菌时,从3种不同的抗性水稻品系中分别发现了1 429、1 927、1 981个lncRNAs参与其中。Wang等发现了与稻瘟病病菌相关的水稻基因,其中有83个lncRNAs在感染稻瘟病病菌后上调,78个下调,其中1个上调的lncRNAs来源于茉莉酸合成基因,茉莉酸调节水稻的稻瘟病抗性,由此可知lncRNAs可通过参与茉莉酸的合成调节水稻稻瘟病的抗性。Tang等在水稻的感染和发育过程中发现了与稻曲病相关的lncRNAs,并对其做了RNA-Seq分析,得到了1 724个lncRNAs。Cao等研究水稻防御黑条矮缩病毒(RBSDV)和条纹病毒(RSV)时,发现了1 925个lncRNAs。其中可能以脱落酸通路中涉及的脱落酸应答元件结合因子()()为靶点,通过靶向病原体的方式参与脱落酸信号通路,从而影响脱落酸的表达。Zhang等对感染和未感染黑条矮缩病毒的水稻进行了转录组分析,发现22个lncRNAs被鉴定为差异表达基因,其中大多数基因参与水稻和病原体的相互作用,并在感染后表现为上调表达。
了解昆虫lncRNAs的特性和功能对于水稻病虫害防御有很大作用。Xiao等在研究褐飞虱对水稻的影响中检测到2 439个lncRNAs,这些lncRNAs在褐飞虱的高繁殖力和毒力适应性方面均有重要作用。Chen等对条纹叶枯病病毒在灰飞虱中的传播过程中发现,lncRNA()可能参与了水稻免疫系统抵抗病毒入侵的活动。袁祝婷对褐飞虱lncRNAs基因的表达及功能分析中发现,lncRNAs参与了褐飞虱的生殖发育,通过干扰褐飞虱的生殖过程可以达到阻止病毒传播的目的。研究发现,lncRNAs有可能在未来发展成为有价值的抗植物病原体的分子农药。
2.3 生长发育
lncRNAs在调节水稻的生长发育方面也发挥着重要作用(表2)。Wang等研究发现了1种由相邻基因簇的反义链转录而来的lncRNA,命名为。过表达时,几个基因表达水平上调,达到水稻增产的效果。Zhang等研究发现,和作为标靶模拟物作用于相关基因,其在雌蕊、花药、早期穗及种子的表达中极高,在穗部发育和繁殖中也发挥作用。这些结果表明,lncRNAs可以调节花和种子发育,在穗发育和繁殖中也发挥着重要作用。Wang等证明了通过靶向模拟的方式减弱了osa-miR160在花药发育早期对(mRNA)的抑制,通过负调控osa-miR160的方式增强了的表达。上述研究表明具有靶向模拟功能的lncRNAs可能作为时间调节因子来调节特定发育阶段的miRNAs的作用,以此参与水稻的生殖生长过程。Luan等报道了可能是来源于基因的反义lncRNA,其表观遗传修饰在水稻穗部发挥作用。Wang等发现了与种子形态和叶片形态相关的基因和。这些lncRNAs在水稻生长发育中起重要作用。内源性 lncRNA()是从水稻中转录因子基因座的相反链转录而来的,的下调和的过表达都会导致转基因水稻的叶片畸变,表明可能顺式调控,使叶片形态在生长过程中发生改变。
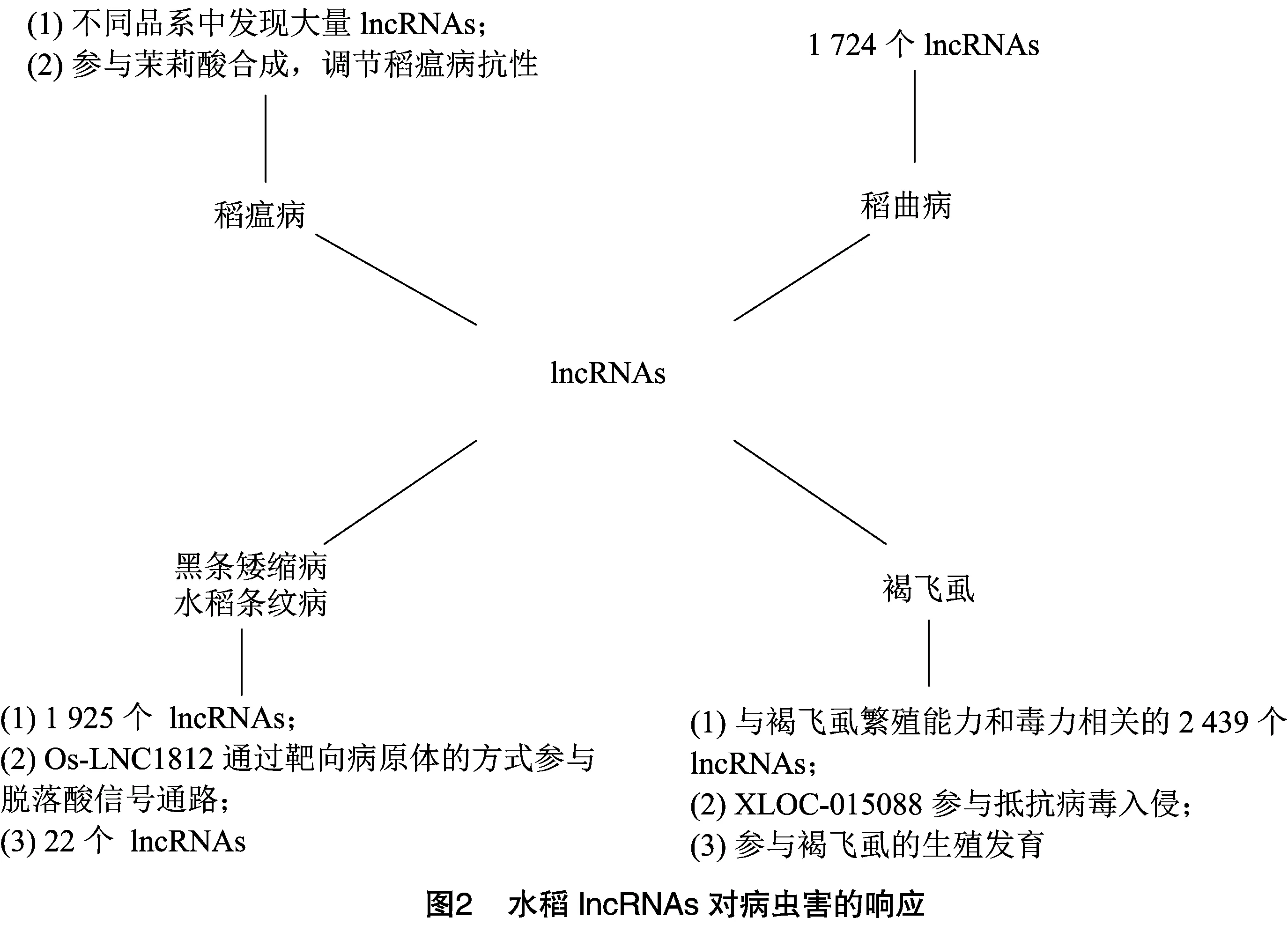

表2 lncRNAs在调控水稻生长发育中的作用
雄性不育系是杂交水稻育种过程中的一个重要基础。Fan等发现由光周期敏感雄性不育()基因座产生的lncRNAs与水稻光周期敏感不育有关。位点参与的编码过程。在长日照条件下,参与了花药的积累过程。当这些lncRNAs积累到一定水平时,会导致雄性不育。Ding等克隆了1个长度为1 236个碱基的基因,用于控制光周期敏感雄性不育,命名为。在长日照条件下,水稻花粉的正常发育需要一定水平的基因表达量,而是通过影响启动子区域的DNA甲基化来调节水稻光周期敏感雄性不育。
Liu等在水稻生殖发育过程中发现了大量的lncRNAs。预测这些lncRNAs可能参与了水稻细胞减数分裂、有丝分裂及雌配子体的发育等过程。在此过程中,lncRNAs可能通过作用于相关基因、作为miRNA前体或诱饵分子等机制参与水稻胚珠发育和雌配子体败育。但这一研究仍处于预测阶段,具体功能仍待进一步验证。
综上所述,水稻lncRNAs调节光周期敏感雄性不育、减数分裂前生殖细胞的发育、减数分裂的进程、胚珠发育和雌配子体发育的过程,从而调节水稻生殖过程。lncRNAs通过影响重要的农艺性状,如籽粒产量、雄性不育和叶型,在调节水稻生长发育中起着复杂的作用。lncRNAs的功能不仅仅是在应激反应中,还有更多lncRNAs在细胞发育过程、激素信号和生理需求中的功能还有待探索。对水稻生长发育相关基因的进一步研究可能会促进水稻产量、品质等的提高。
3 展望
现有的科学研究丰富了水稻lncRNAs的研究内容,补充了该领域的研究范畴,填补了研究空白,对水稻lncRNAs的发展具有非常重要的作用。展望未来,可以从以下几个方面加强lncRNAs的深入研究。
(1)lncRNAs的结构复杂,样本数量不足,识别较为困难。虽然存在许多lncRNAs数据库,但大多数数据库只关注少数特定物种,对于水稻lncRNAs注释的数量很少。许多已发表的报告证实了水稻lncRNAs的关键生物学作用(例如细胞分化、基因调控、胁迫响应等)。然而,对于这种特定的lncRNAs,仍然缺乏以水稻lncRNAs为主的数据存储和功能注释的数据库。目前已知的水稻相关lncRNAs数据库规模很小(仅包含数百个lncRNAs),并且在其重点(疾病或相互作用)方面具有特定性。对此,可以针对水稻实现统一的数据库管理,提出相应的lncRNAs注释及预测,有效提高水稻lncRNAs的研究。
(2)lncRNAs预测软件在动物上特别是癌症方面研究较多,在植物上,尤其是水稻方面研究较少,目前还没有一种科学界一致同意的预测方法,因此很难进行跨物种的比较。如果想进一步探究 lncRNAs在水稻中的调控机制,还需要结合多个学科进行研究,如生物信息学、遗传学等。同时,随着相关技术的进一步发展,我们将对lncRNAs的功能和机制进行更加完整的理解和研究。lncRNAs也将在作物遗传育种、生物资源开发、植物细胞工程等领域发挥更加重要的作用。
(3)虽然目前对lncRNAs在水稻中的功能研究越来越多,但有关水稻的研究仍然较少。目前已有大量研究发现lncRNAs参与水稻胁迫、抗逆及生长发育等过程,但针对其响应的全面调查仍然有限,许多lncRNAs仍然不知道确切的基因组注释和功能意义。随着lncRNAs新的来源不断被发现的同时,其分类的方法也应不断更新。此外,lncRNAs并不是以单一的方式发挥作用,通常与其他非编码基因和蛋白编码基因相互作用而发挥功能。目前很多研究都表明lncRNAs可与miRNA相互作用发挥调控机制,一条lncRNA可能受不同作用机制影响。加强对lncRNAs功能方面更加科学有效的研究,使lncRNAs在水稻育种、种质创新等方面发挥更重要的作用。
