不同海拔的小粒咖啡光合特性日变化研究
2022-08-10胡发广刘红明毕晓菲付兴飞李亚男吕玉兰黄家雄
胡发广,刘红明,毕晓菲,付兴飞,李亚男,杨 旸,吕玉兰,黄家雄
(云南省农业科学院 热带亚热带经济作物研究所,云南 保山 678000)
咖啡是茜草科(Rubiaceae)咖啡属(Coffea)多年生常绿灌木或小乔木,原产于非洲。咖啡的产量、产值和消费量均居于全球三大饮料作物之首[1],是仅次于石油的世界第二大贸易商品。咖啡已有数百年的饮用历史,是一类产业链长、经济价值高的经济作物。咖啡饮品主要由咖啡豆制成,因焙炒咖啡豆含有丰富的咖啡因、生物碱、绿原酸、粗多糖、有机酸、咖啡油脂等成分,具有独特的醇香口味和提神作用,咖啡已成为风靡全球的日常饮品。截止到2020年底,云南省咖啡种植面积92533.33 hm2、产量14.50万t、农业产值22.28亿元,均占全国98%以上。云南省咖啡产业的种植面积、产量和产值均居全国第一。
作物的光合能力是产量和品质形成的基础,逾90%的干物质来自叶片的光合产物,光合速率影响光合产物的积累,在一定程度上决定产量的高低[2-3]。海拔高度对果树的生态影响主要是通过光、热、水、气等生态因子起作用[4]。随着海拔的升高,太阳辐射、气温、大气中O2和CO2分压等均呈现一定的变化趋势[5]。目前,国内外已有学者开展了海拔高度对小粒咖啡相关理化性状影响的研究工作,结果表明,高海拔地区的小粒种咖啡在咖啡鲜果与咖啡豆的产量上高于低海拔地区,且咖啡豆颗粒大,所含的圆豆与缺陷豆量较少,高海拔地区的咖啡果实品质优于低海拔地区[6]。海拔高度与咖啡豆千粒重、糖含量呈显著相关,与咖啡因含量呈极显著负相关,与粗脂肪含量呈显著负相关[7]。
本文以保山市潞江镇新寨村委会万亩咖啡园为研究地点,比较小粒咖啡[Catimor CIFC 7963(F6)]随着海拔的升高,净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)、叶片温度(Tleaf)、光合有效辐射(PAR)等光合特性差异,探究不同海拔高度对小粒咖啡光合作用及果实品质的影响,以期为优化不同海拔区域的小粒咖啡栽培管理措施提供参考依据。
1 材料与方法
1.1 材料
1.1.1 试验地概况 样品采集地位于保山市潞江镇新寨村委会万亩咖啡园,属低纬度准热带季风雨林偏干热河谷过渡类型气候,干热少雨,地面蒸发量大,干湿季分明,旱季一般从11月开始到次年5月,光照充足,年温差小,昼夜温差大。年平均气温21.5 ℃,年平均降雨量755.3 mm,绝对最高温40.4 ℃,绝对最低温0.2 ℃,全年基本无霜,≥10℃的活动积温7800.0 ℃·d,年日照时数2333.7 h,年均空气湿度70%。
1.2 试验设计
试验于2019年10月选择种植年限超过10年以上的咖啡老园(定植规格为株距1 m,行距2 m),品种为Catimor CIFC 7963(F6)。分别选取海拔730、917、1064、1285、1489 m等5个海拔点的光合作用日变化进行田间测定,每个测试点基本特性见表1。各测试点选取生物学性状基本一致的15株参与测定。试验采用LCpro-SD全自动便携式光合测定仪(英国ADC公司),选择晴朗无云天气,从8:00~18:00每隔2 h测定1次,每个品种随机选取3株,每株5片叶测定。测定时选取树冠中上部南面的当年成熟叶片,叶龄相对一致。测定时每片叶片重复测定3次,取其平均值。主要测量指标:净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)、叶片温度(Tleaf)、光合有效辐射(PAR)等。

表1 测试点基本特性
1.3 数据分析
叶片水分利用率(WUE)计算公式如下:WUE=Pn/Tr,光能利用效率(LUE由计算公式:LUE(%)=Pn×100/PAR计算得到。运用Microsoft Excel 2013软件和SPSS 13.0统计分析软件完成数据统计及相关性分析。
2 结果与分析
2.1 环境因子的变化特征
2.1.1 大气温度 由图1可知,在整个试验阶段,不同海拔的咖啡大气温度变化趋势基本一致,即都是先升后降的单峰型。但不同海拔的咖啡大气温度变化也不尽相同,其中低海拔样点(730 m)从上午8:00~14:00温度逐渐升高,12:00~16:00为一天中的最高温时期,16:00后温度逐渐下降。中海拔样点(917、1064 m)从上午8:00~10:00温度迅速升高,10:00~16:00为一天中的最高温期间,高温期间显著延长,16:00后温度迅速下降。高海拔样点(1285、1489 m)从上午8:00~10:00温度迅速升高,10:00~14:00为一天中的最高温期间,14:00后温度逐渐下降。

图1 咖啡叶片大气温度日变化
2.1.2 叶片温度 由图2可知,在整个试验阶段,咖啡叶面温度变化趋势与大气温度变化趋势基本一致,即都是先升后降的单峰型。但不同海拔咖啡大气温度变化也不尽相同,其中低海拔样点(730 m)从上午8:00~14:00温度逐渐升高,12:00~16:00为一天中的最高温期间,16:00后温度逐渐下降。中海拔样点(917、1064 m)从上午8:00~10:00温度迅速升高,10:00~16:00为一天中的高温期间,高温期间显著延长,16:00后温度迅速下降。高海拔样点(1285、1489 m)从上午8:00~10:00温度迅速升高,10:00~14:00为一天中的最高温期间,14:00后温度逐渐下降。

图2 咖啡叶片叶面温度日变化
2.1.3 光合有效辐射 从图3 可以看出,不同海拔样点光合有效辐射变化趋势都呈单峰型,但不同海拔样点光合有效辐射日变化存在一定差异。其中低海拔样点(730、917 m)从8:00~10:00快速升高,10:00~16:00为一天中的光合有效辐射峰值期间,光合有效辐射峰值期间显著延长,16:00后光合有效辐射迅速下降;中高海拔样点(1064、1285、1489 m)从8:00~10:00快速升高,从10:00~14:00为一天中的光合有效辐射峰值期间,光合有效辐射峰值期间缩短,14:00后光合有效辐射快速下降。
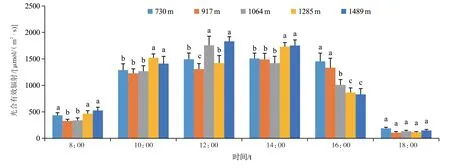
图3 光合有效辐射日变化
2.2 不同海拔梯度咖啡光合参数日变化比较
2.2.1 不同海拔梯度咖啡胞间CO2浓度日变化比较从图4可以看出,5个不同海拔高度咖啡叶片的胞间CO2浓度日变化存在一定差异。低海拔样点(730、917 m)变化趋势呈“W”型,从上午8:00开始急剧下降,后开始回升,12:00又开始下降,海拔713 m样点在16:00出现第2个低点后急剧上升;海拔930 m样点在14:00出现第2个低点后缓慢回升。中海拔样点(1064、1285 m)变化趋势呈“U”型,从上午8:00~14:00缓慢下降,后迅速回升。高海拔样点(1489 m)变化趋势呈“W”型,从上午8:00开始急剧下降,后开始回升,12:00又开始下降,14:00出现第2个低点后缓慢回升。平均胞间CO2浓度从大到小排列为:1064 m>1285 m>1489 m>917 m>730 m。

图4 不同海拔梯度咖啡胞间CO2 浓度日变化
2.2.2 不同海拔梯度咖啡的净光合速率日变化比较从图5可以看出,大部分咖啡品种在潞江坝干热河谷区的净光合速率日变化呈双峰变化趋势,有明显的光合“午休”现象;低海拔样点(730 m)则呈单峰曲线变化,从8:00开始急剧上升,至上午10:00达到一天中的最大值,后缓慢下降。5个海拔样点Pmax值从大到小排列为:1285 m>730 m>1064 m>1489 m>917 m。

图5 不同海拔梯度咖啡的净光合速率日变化
2.2.3 不同海拔梯度咖啡的气孔导度日变化比较从图6可以看出,不同海拔高度咖啡叶片气孔导度变化趋势一致,均为“J型”曲线变化趋势,上午8:00,所有海拔梯度咖啡的气孔导度均为最大值,后急剧下降。从10:00和18:00气孔导度则呈缓慢下降趋势,18:00时低海拔的730 m处气孔最先关闭。

图6 不同海拔梯度咖啡的气孔导度日变化
2.2.4 不同海拔梯度咖啡的水分利用率日变化比较从图7可以看出,不同海拔咖啡水分利用率日变化差异显著,所观测的5个海拔样点咖啡均呈现出2次峰值。其中低海拔(730、917 m)水分利用率的第1次峰值出现在10:00,中海拔(1064、1285 m)水分利用率的第1次峰值出现在8:00,高海拔(1489 m)水分利用率的第1次峰值出现在14:00。5个海拔样点咖啡水分利用率的第2次峰值均在18:00。各海拔咖啡的日均水分利用率从大到小依次为1489 m>917 m>1285 m>1064 m>730 m。

图7 不同海拔梯度咖啡的水分利用率日变化
2.2.5 不同海拔梯度咖啡的光能利用率日变化比较从图8可以看出,5个海拔样点咖啡光能利用率在8:00最高,8:00~14:00之间快速下降,直到光合午休时达到最低值,然后逐步回升。各海拔咖啡的日均光能利用率从大到小依次为1064 m>917 m>730 m>1285 m>1489 m。
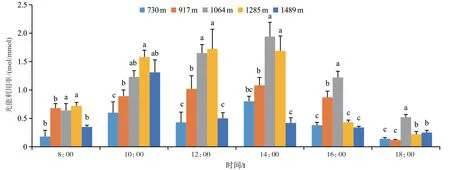
图8 不同海拔梯度咖啡的光能利用率日变化
3 讨论
光合作用是植物生长发育的基础和干物质积累的主要来源,是植物光合生产力的重要衡量指标,也是作物产量和品质构成的决定性因素[8]。光合作用是一个极其复杂的生理过程,受植物本身的生理状况和生态因子的双重影响 。海拔高度直接导致生态因子的改变,太阳辐射、气温、大气中的O2和CO2分压等随着海拔的升高而呈现一定的变化趋势[9]。云南省地处中国西南边陲,山地较多,地貌复杂,海拔变化差异大,其特殊的地理位置影响作物的种植园规划和建设。海拔高度的变化对作物的生长发育和光合特性有明显的影响[10]。
环境是植物赖于生存和发展的条件,光合作用是植物物质代谢和能量代谢的重要基础,同时也是受环境影响变化最显著的生理过程之一[11]。海拔变化会导致光照、温度、湿度、大气压及CO2分压等环境的变化,影响作物的光合能力[9]进而影响植物的生态和生理特征[12]。
张翠仙等[13]在杧果上的研究表明,杧果的净光合速率、叶肉瞬时羧化效率随着海拔的升高而降低,蒸腾速率随着海拔的升高呈现先升高后降低的趋势。国内外有很多文献从咖啡品质的角度对不同海拔咖啡进行了探讨,但尚未涉及海拔对光合作用影响的报道。研究表明,咖啡豆千粒重和总糖含量均与海拔高度呈显著正相关;咖啡因含量与海拔高度之间呈极显著负相关;粗脂肪含量与海拔高度之间呈显著负相关;粗蛋白质、粗纤维、水浸出物、灰分与海拔高度之间的相关性不显著[7]。张洪波等[14]对云南小粒种咖啡高海拔种植适应性进行了分析和评价,进一步明确在适宜的范围内,海拔越高,杯品质量越好。井锐锐等[15]对4个不同海拔区域种植的尤力克柠檬研究表明,4个海拔区域尤力克柠檬的光合有效辐射和大气温度均呈单峰型变化曲线,叶表面的相对湿度呈低谷型变化曲线,峰值和低谷均在中午12:00;4个海拔区域尤力克柠檬的净光合速率均呈单峰型变化曲线,随着海拔的升高呈增加趋势,到1110 m出现下降。800和1110 m 显著高于420和300 m 的净光合速率。
龚荣高等[16]发现随着海拔的升高,青脆李的最大净光合速率、暗呼吸速率和光补偿点都出现了先增后降的变化。本研究表明,对同一咖啡品种来说,不同海拔地区的光合特性存在较大差异。不同海拔咖啡的大气温度、叶片温度和光合有效辐射变化趋势基本一致,即都是先升后降的单峰型,不同之处在于温度升高后再降低的时间点不同。Catimor咖啡在潞江坝不同海拔地区的光合特性存在较大差异,除最低海拔样点(730 m)外,最大净光合速率随着海拔的升高,出现了先增后降的变化趋势。不同海拔咖啡的气孔导度、水分利用率和光能利用率最大值均出现在上午8:00,随着海拔的升高,均呈现先增后降的变化。
温度因子是植物生长中重要且复杂的影响因素之一,在一定温度范围内,光合速率随温度的升高而增大,这是因为温度直接影响光合作用关键酶-Rubisco的活性[17];而海拔对温度的影响最大,随着海拔的升高,温度不断下降,植物的光合能力也随之下降,其营养生长和生殖生长的时间也随之缩短[18]。在本研究中,不同海拔光合速率与大气温度的变化趋势一致,随着海拔的升高,出现了先增后降的变化。
