基本情绪的神经基础:来自fMRI与机器视觉技术研究的证据*
2022-08-09肖婷炜王福顺
梁 飞 江 瑶 肖婷炜 董 洁 王福顺
基本情绪的神经基础:来自fMRI与机器视觉技术研究的证据*
梁 飞 江 瑶 肖婷炜 董 洁 王福顺
(四川师范大学脑与心理科学研究院, 成都 610066)
基本情绪理论(basic emotion theory)是情绪科学领域最具代表性的理论, 该理论认为人类情绪是由有限的几种基本情绪组成的, 如恐惧、愤怒、喜悦、悲伤等。基本情绪是为了完成基本生命任务(fundamental life task)进化而来的, 每一种基本情绪都有独特的神经结构和生理基础。尽管基本情绪理论被广泛接受, 但是对于基本情绪的种类却莫衷一是。近几十年来, 许多fMRI研究试图确定各种基本情绪的独特神经结构基础, 而且取得了许多重要发现, 比如厌恶和脑岛有关, 悲伤和前扣带回有关, 杏仁核是与恐惧有关的重要边缘结构等。但是, 最近有人进行了元分析研究, 发现许多基本情绪存在混淆的大脑区域, 因此对基本情绪的特定脑区理论提出质疑, 甚至否定基本情绪理论。通过对基本情绪及其神经基础的探讨, 以及对基本情绪理论的最新功能性磁共振成像研究进行梳理分析, 提出有关基本情绪理论的争论来源于基本情绪种类的确定, 因为许多所谓的不同基本情绪实际上是同一种基本情绪, 提出人类可能只有3种基本情绪。未来研究可以利用机器视觉技术进一步推动基本情绪脑影像研究。
基本情绪, 情绪维度, fMRI, 机器视觉, 脑影像
1 引言
情绪是人类生命活动的基本组成部分(LeDoux et al., 2016), 在人类生活和精神疾病中起着至关重要的作用。许多精神疾病涉及情绪功能障碍, 深入了解情绪的本质和机制对情绪研究和情感疾病治疗都至关重要(Dolensek & Gogolla, 2021)。基本情绪理论(basic emotion theory)是情绪研究最经典的成果之一(Ekman, 1992, 1999; Ekman & Davidson, 1994; Scarantino & Griffiths, 2011; Hutto et al., 2018), 它具有很大的影响力, 推动了面部表情检测、神经影像研究和进化心理学等许多实证研究(Hutto et al., 2018)。基本情绪理论认为, 人类具有在生理和心理上有限的几种“基本的”情绪, 例如恐惧, 愤怒, 喜悦, 悲伤等(Wilson- Mendenhall et al., 2013; Gu et al., 2019b), 每一种基本情绪都有其独特的神经结构基础和生理功能表现。尽管基本情绪理论被广泛接受, 但是人类具体有几种基本情绪却莫衷一是。甚至, 最近元分析研究发现某些基本情绪存在共用的大脑区域, 因此对基本情绪的特定脑区理论提出质疑, 甚至否定基本情绪理论, 从而重提情绪维度理论, 两个理论的争论持续了近百年(Lindquist et al., 2012)。
脑影像技术研究试图确定各个基本情绪的独特神经结构, 为基本情绪理论提供支持。情绪通常被理解为大脑皮层和皮层下区域的限定区域的活动(Pessoa, 2017)。研究情绪的神经元机制的主要障碍之一是难以将可观察的行为与正在进行的神经活动联系起来(Dolensek & Gogolla, 2021)。而功能性磁共振成像技术(functional magnetic resonance imaging, fMRI)可探测到深部脑区的活动, 因此常被用以探索基本情绪的大脑基础和中枢神经活动。脑影像研究为基本情绪理论提供了重要的实验证据, 比如Vytal和Hamann (2010)的激活似然估计(activation likelihood estimation, ALE)元分析结果支持了基本情绪理论:每一种被检测的情绪(恐惧、愤怒、厌恶、悲伤和快乐)都具有一致的神经相关性, 它们与局部大脑激活存在可靠的相关性。但是, Barrett及其同事却认为, 在诱发愤怒、悲伤、恐惧、厌恶和快乐情绪时, 边缘区和非边缘区的激活度都存在一致增加, 但不具有特异性(Touroutoglou et al., 2015)。他们认为, 大脑的神经结构中并不存在每种情绪的特定脑网络。基本情绪与特定脑区、神经结构是否存在一一对应的联系, 成为基本情绪理论是否成立的关键。
我们认为出现这些脑影像学不一致的现象可能是因为有些情绪不是基本情绪。尽管基本情绪理论目前被广泛接受, 但是具体的基本情绪种类还不确定。基于此, 本文以基本情绪理论及其争论为线索, 提出这些争论来源于基本情绪的种类确定, 因为某些所谓的不同基本情绪实际上是同一种基本情绪。具体来说, 本文将(1)以基本情绪理论为核心对情绪理论进行梳理讨论; 提出以基本情绪理论为基础对情绪理论进行整合的方案, 以推动情绪理论之争的解决; (2)对当前基本情绪的fMRI实证研究进行梳理, 探讨基本情绪的神经基础; (3)对未来基本情绪的脑影像研究进行展望, 提出将机器视觉技术应用于基本情绪的脑影像研究, 构建人类面部表情和fMRI成像的基本情绪原型。由此, 以期能够为基本情绪理论的实证研究提供新的思路和验证。
2 最具代表性的情绪理论:基本情绪理论和情绪维度理论
情绪科学领域存在一个长期的争论:情绪是按照不同的类别组织起来的(如分为恐惧、悲伤、快乐和惊讶等类别), 还是按效价、唤醒等情绪维度进行连续排列的(Barrett et al., 2018), 即情绪维度理论与基本情绪理论, 这是最具代表性的两种情绪理论。在此, 我们将重点对两种经典情绪理论进行讨论, 同时对其他情绪理论进行梳理。
2.1 基本情绪理论
基本情绪理论(basic emotion theory, BET)认为, 某些情绪是生物进化过程中最基本的、具有跨文化一致性的情感过程(Ekman, 1972; Griffiths, 1997; DeLancey, 2002, Hutto et al., 2018)。基本情绪理论是最盛行的理论之一, 该理论认为人类情绪是由有限的几种基本情绪组成的, 如恐惧、愤怒、喜悦、悲伤等, 它们之所以“基本”, 是因为它们能够完成独特的生理功能, 并且各自有着独特的生理表现和神经结构(Saarimäki et al., 2016; Celeghin et al., 2017; Williams, 2017; Hutto et al., 2018; Song & Hakoda, 2018; Vetter et al., 2018; Wang et al., 2018; Gu et al., 2019a; 梁飞等, 2021)。基本情绪理论提出, 情绪的神经结构基础是不同的, 每种情绪都有其独特的神经回路(Gu et al., 2019a)。
基本情绪理论研究最早起源于达尔文的研究,盛行于Ekman和Friesen在新几内亚的研究(Ekman et al., 1969; Keltner, Sauter et al., 2019)。Ekman等通过使用典型情绪面部表情的静态照片,提出了6种基本情绪, 包括高兴(happiness)、悲伤(sadness)、愤怒(anger)、厌恶(disgust)、恐惧(fear)、惊讶(surprise)等6种(王福顺等, 2018; Keltner, Tracy et al., 2019)。随后许多心理学家从不同的角度对基本情绪做了大量研究, 并得到了一致的结论。许多学者提出了不同种类的基本情绪, 详见表1 (Ortony & Turner, 1990; Clore & Ortony, 2013; Gu et al., 2019a)。对表中15种基本情绪分类进行统计, 有12种包含愤怒, 为最多的一种情绪, 接着依次为快乐11种、恐惧10种、厌恶8种、悲伤8种(e.g., Ekman et al., 1969; Levenson, 2003; Kapucu et al., 2021)。最多的要数Cowen和Keltner (2020)提出的25种基本情绪, 他们从情绪的计算理论(Computational Approach to Emotion)提出了情绪的语义空间理论(Semantic Space Theory), 使用广泛的自然刺激和开放式统计技术来捕捉与情绪相关行为的系统变化, 结果表明人类至少有25种不同的基本情绪。比较少的是Jack等人(2014)提出的4种基本情绪, 他们在对动态面部表情的研究中, 通过结合知觉期望模型、信息论和贝叶斯分类器发现, 在信号动力学的早期, 面部表情系统地传递很少的、有生物学基础的面部信号, 支持更少的基本类别的分类(例如, 趋近/回避)。进一步的多学科跨文化研究中, Jack等人(2016)的数据揭示了4种潜在的、文化上常见的面部表情模式(喜、怒、哀、惧)。除此之外, 我们实验室在国际上提出了新的基本情绪理论——情绪三原色学说。该理论认为, 人类有3种基本情绪:喜悦、恐惧、悲伤, 这3种基本情绪分别具有不同的单胺类神经递质基础:去甲肾上腺素(norepinephrine, NE)与应对压力事件的“战斗或逃跑”反应有关, 多巴胺(dopamine, DA)参与奖赏机制, 五羟色胺(serotonin, 5-HT)与惩罚有关如图1 (Gu et al., 2019b)。情绪三原色学说原创性地从单胺类神经递质的角度来区分基本情绪, 探讨了更少的基本情绪存在的可能性。
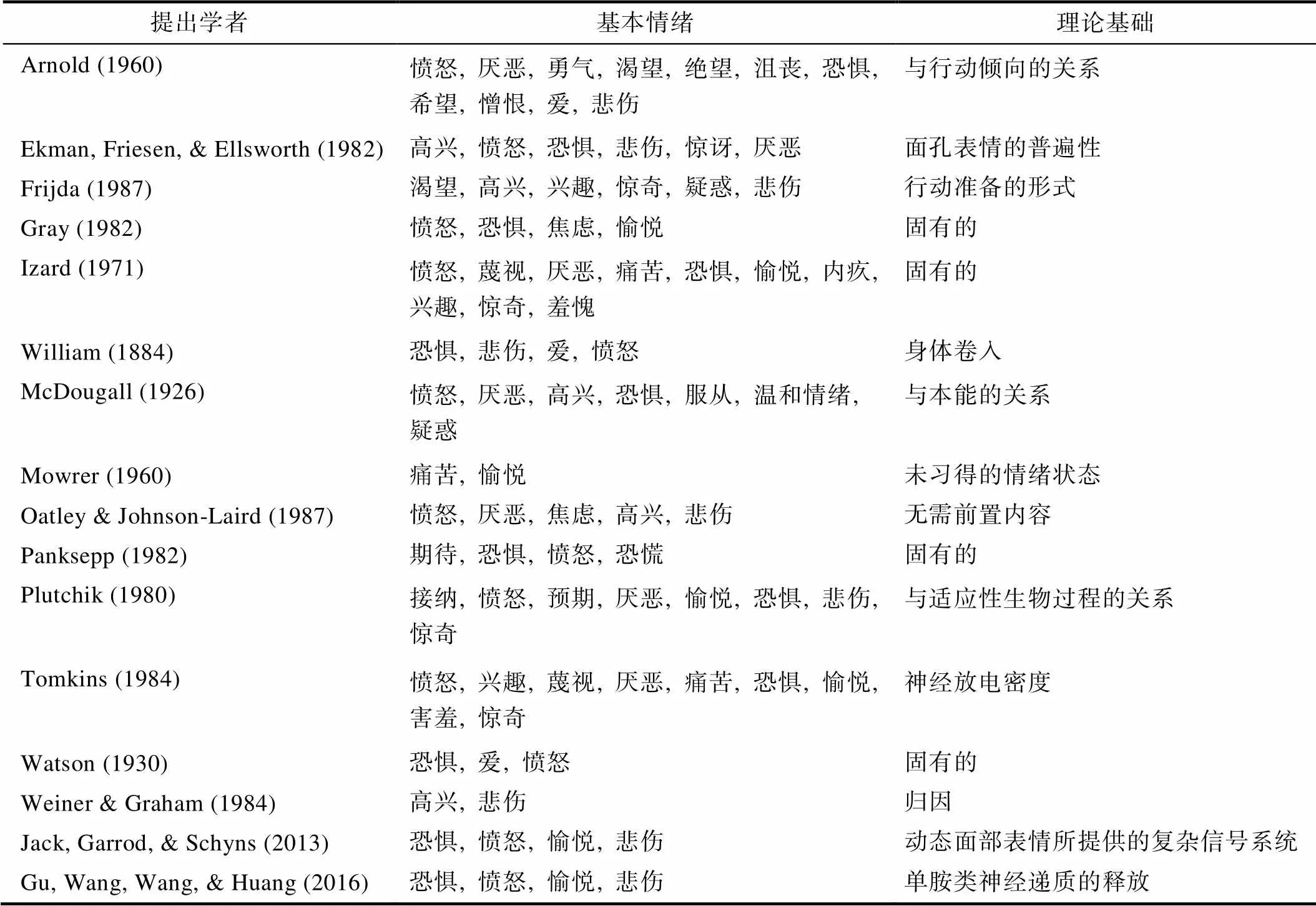
表1 基本情绪的分类

图1 基本情绪的三原色模型。在本模型中, 作者提出了人类只有3种基本情绪:恐惧(愤怒)、快乐、悲伤; 这3种基本情绪也可以称为核心情绪, 它们会通过释放3种单胺类神经递质:去甲肾上腺素−恐惧(愤怒)、多巴胺−奖励和五羟色胺−惩罚而起作用(Gu et al., 2018)。
2.2 情绪维度理论
基本情绪理论认为每种基本情绪都具有其独特的神经生理特点; 但是最近元分析研究对基本情绪的特定脑区理论提出质疑, 甚至否定基本情绪理论, 从而重提情绪维度理论(Lindquist et al., 2012)。情绪维度理论认为, 情绪维度是所有情绪体验的核心, 分为效价(valence)和唤醒度(arousal) (Cowen & Keltner, 2017), 它们构成了一个象限的两个独立的维度。情绪维度理论最早由冯特(Wundt)提出, 他当时提出了情绪的三维度学说, 后来的许多实验证明只要两个维度就可以代表情绪的特点:效价和唤醒度(Gu et al., 2019a)。唤醒与效价评分之间不相关的证据证实了情绪维度模型, 即情绪有两个不同的维度; 根据维度模型, 情绪被定义为两个主要维度, 唤醒度即从平静到兴奋, 效价从悲伤到喜悦(Russell, 1980; Kapucu et al., 2021)。但是情绪维度理论也有不足之处, 因为效价维度很难有程度上的区别; 换言之, 效价维度上没有办法表达喜悦的程度。喜悦和更喜悦应该是在效价维度上的区别, 还是在唤醒度上的区别, 这一问题是模糊的。因此, Russel提出了情绪环学说, 认为所有的情绪都可以落在一个圆环上(Gu et al., 2019a)。
Gu等人(2019a)认为, 基本情绪和维度研究并不是对立的, 相反, 它们可以使彼此更加清晰地解释情绪; 他们回顾了脑成像和神经心理学的数据, 并尝试用整合模型来解决基本情绪与维度理论之争(图2)。Gu等人提出了4种基本情绪:快乐−处于快乐维度的正轴上, 悲伤−处于快乐维度的负轴上, 恐惧和愤怒是“双胞胎”情绪, 它们位于垂直维度的顶端(Gu et al., 2019a)。这种新的整合模型有望成为基本情绪理论与维度理论争论的出路, 改进和重新构建基本情绪的神经生物学模型。
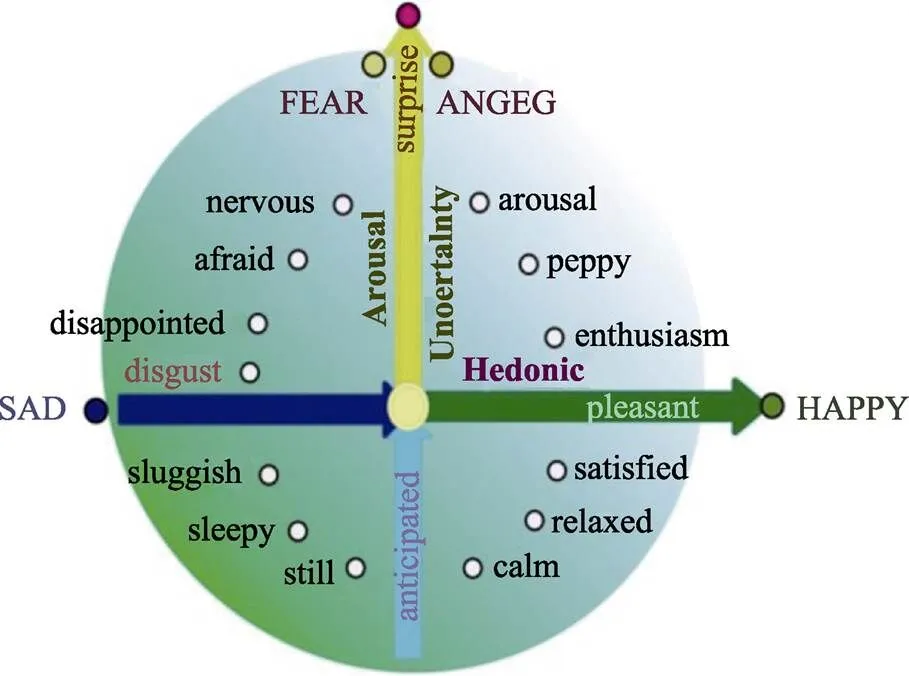
图2 基本情绪理论与建构取向(维度理论)的整合模型(integrative model)。整合理论认为基本情绪理论和维度理论不矛盾, 基本情绪之所以基本, 是因为它们落在了坐标轴上。因此, 它们代表了情绪的某一个特点:喜悦落在了横坐标的右侧, 代表了正性效价, 悲伤落在了横轴的左侧, 代表了负性效价, 恐惧(愤怒)代表了最强烈的唤醒度(Zheng et al., 2016; Gu et al., 2019a)。
综合上述, 尽管基本情绪理论受到了来自情绪维度理论及其他情绪理论的质疑, 但是基本情绪理论的观点得到了许多研究的支持, 许多研究报告了基本情绪的神经相关。如Saarimäki等人(2016)采用多元模式分析法(multivariate pattern analysis)对厌恶、恐惧、快乐、悲伤、愤怒和惊讶6种基本情绪的脑活动模式进行分类, 验证了情绪的分类模型, 即人类基本情绪在神经和生理上是不同的。我们将在下文对最新的基本情绪脑影像研究进行探讨。
3 基本情绪的脑影像研究——脑激活与情绪的相关
基本情绪理论提出, 每种情绪都有其独特的神经回路。因此, 研究神经基础可能是区分基本情绪和探讨基本情绪数量的最佳途径。脑影像研究为基本情绪理论提供了重要的实验证据, 比如, 脑影像研究发现, 大脑中有一些特定的核团负责特定的基本情绪, 而其他区域通常参与情绪的感知、评估或调节(Phan et al., 2002; Lindquist et al., 2012, 2013; Gu et al, 2019a)。比如, Ekman所推崇的6种基本情绪都有其特定的神经结构。
3.1 快乐(Happy)
快乐情绪比中性情绪诱发出更大的视觉皮层活动(Britton et al., 2006)。Vytal和Hamann (2010)对功能磁共振的大量基本情绪研究进行元分析发现, 与中性刺激相比, 快乐情绪激活吻侧前扣带回(crostral ACC)和右颞上回(the right superior temporal gyrus, STG); 而Suardi等(2016)的研究报告喜悦情绪激活了更广泛的区域, 包括前扣带皮层(anterior cingulate cortex), 前额叶皮层(prefrontal cortex)和脑岛(insula)。快乐音乐能激活双侧听觉皮层、左侧海马旁回和辅助运动区(Bogert et al., 2016)。当母亲看到自己孩子快乐的面部表情时, 海马被激活(Kluczniok et al., 2017)。以快乐图片作为刺激材料的fMRI研究表明, 快乐比悲伤在楔前叶和后扣带皮层表现出更强的激活(An et al., 2018)。腹侧被盖区(ventral tegmental area, VTA)是中脑边缘多巴胺能系统的起源, 该系统向蓝斑、前额叶皮层和前扣带皮层投射和释放多巴胺, 负责积极情绪的认知效应(Gu et al., 2019a)。这表明, 快乐情绪激活了较为广泛的大脑区域, 可能与前扣带皮层、前额叶皮层、脑岛、海马等存在关联。不同刺激类型诱发的快乐情绪可能存在一致的神经基础。
3.2 恐惧(Fear)
杏仁核是一个与恐惧至关重要的边缘结构(Anthony et al., 2014; Isosaka et al., 2015; Reynaud et al., 2015; Han et al., 2017; Gu et al., 2019a)。恐惧刺激比中性刺激显示出更大的海马、前扣带回、视觉皮层激活(Britton et al., 2006)。当呈现恐惧面孔时, 双侧的视觉皮质(visual cortex)和顶上小叶(superior parietal lobule)活动显著, 在海马回(parahippocampal gyri)、杏仁核(amygdala)和全海马(hippocampi)的边缘结构中检测到较大的激活簇(Rahko et al., 2010)。Thomas等人(2019)对新生儿的研究表明, 杏仁核−脑岛的连接与恐惧有关。Phan等人(2002)发表在Neuroimage的元分析文章也总结到, 恐惧特别能使杏仁核得到激活。即使是新生儿, 就已经显示出了恐惧情绪与杏仁核的相关。如新生儿杏仁核功能连接性的独特特征与恐惧、认知发展的出现有关(Graham et al., 2016)。众多的研究表明, 恐惧情绪可能离不开杏仁核的作用。虽然不同研究报告的结果有所差异, 但比较一致的实验证据支持恐惧情绪与杏仁核有关。
3.3 愤怒(Anger)
眶额皮质(orbitofrontal cortex, OFC)和杏仁核之间的相互作用与愤怒的调节有关(Coccaro et al., 2007; Fulwiler et al., 2012; Gu et al., 2019a)。愤怒面孔比中性面孔显示出更大的杏仁核、海马、腹内侧前额皮质(ventromedial prefrontal cortex, vmPFC)激活(Britton et al., 2006)。愤怒控制, 即试图控制愤怒表情的倾向, 与杏仁核−眶额皮质的连接呈正相关(Fulwiler et al., 2012)。右侧杏仁核的灰质体积与特征性愤怒相关, 这与杏仁核作为处理愤怒的核心区域的作用非常吻合(Wang, Kong et al., 2017)。攻击性与杏仁核对愤怒面孔的反应增强有关(Buades-Rotger & Krämer, 2018)。Dzafic等人(2019)近期研究发现, 右杏仁核与边缘区域的功能连接及快速发现、线索一致的愤怒有关。Tonnaer等人(2017)发现在暴力犯罪者愤怒期间, 腹侧前额叶(ventrolateral prefrontal)激活增加。Xiao等(2019)最近的研究表明, 背内侧前额皮质(dorsomedial prefrontal cortex, dmPFC)和vmPFC可能是替代性攻击的神经功能标志物。大量的证据支持了眶额皮质、杏仁核与愤怒情绪可能存在相关关系, 而眶额皮质和杏仁核之间的相互作用与愤怒的调节有关。
3.4 悲伤(Sad)
悲伤与胼胝体扣带回的活动有关(Phan et al., 2002), 回忆悲伤与前脑岛的激活增加有关(Lane et al., 1997)。悲伤面孔比中性面孔能诱发更强的vmPFC、视觉皮层激活, 在呈现悲伤(悲伤在阈下水平)的面部表情时, 前扣带回明显被激活(Britton et al., 2006)。Kluczniok等(2017)的研究也发现, 当母亲们看到自己孩子悲伤的面孔时, 她们的杏仁核和前扣带皮层表现出活跃。Ramirez-Mahaluf等人(2018)悲伤诱发任务的fMRI结果确定了两个主要的脑网络模块:背侧和腹侧, 以及它们的中枢区域:左背外侧前额叶皮层(the left dorsolateral prefrontal cortex, dlPFC)和左前额叶内侧极(left medial frontal pole, mFP)。这些中枢区域并没有在悲伤后调节它们的相互功能连接, 但是它们通过一个中间区域来调节, 即亚属前扣带回皮层(the subgenual anterior cingulate cortex, sACC)。
以声音作为刺激材料诱发悲伤时也有类似的发现。如Bogert等(2016)用fMRI来记录音乐诱发情绪的神经加工, 结果表明悲伤与左侧前扣带回、额中回的激活和眶额皮质的下调有关。
前扣带回皮层的某些部分之间的联系导致了负面情绪的持续存在, 并且前扣带回皮层是在日常生活中维持悲伤情绪非常重要的大脑机制(Schwartz et al., 2019), 悲伤情绪与前扣带回激活关系密切。
3.5 厌恶(Disgust)
厌恶是一种自然的防御情绪, 是为了保护自己免受潜在污染源的伤害(Oaten et al., 2018)。脑岛与厌恶有关, fMRI数据进一步表明后脑岛(posterior insula, PI)和前脑岛(anterior insula, AI)之间存在明显的功能性分离, 其中PI比AI更多地参与初级道德厌恶水平, 而AI比PI更多地参与二级道德厌恶水平(Ying et al., 2018)。早在2004年, Wright的研究结果便支持脑岛对选择性厌恶的加工(Wright et al., 2004)。虽然脑岛的激活并非厌恶的唯一原因, 但前脑岛和基底神经节(在较小程度上)与毒性和疾病相关的厌恶表现形式有关(Chapman & Anderson, 2012)。但是, Ying等人(2018)的研究表明, PI和AI之间存在明显的功能分离, 其中PI比AI更参与初级道德厌恶, 而AI比PI更参与二级道德厌恶。
文字或阅读诱发的厌恶情绪可能也由脑岛参与了加工。近期, Viol等人(2019)的研究也发现脑岛皮层参与了厌恶加工。带厌恶情绪的文字同样也能诱发厌恶情绪。Ziegler等人(2018)发现, 被试在观察厌恶的面部表情或阅读含有厌恶内容的文字时, 左前脑岛的同一区域都会做出反应。不同的刺激材料诱发的厌恶情绪能较为稳定地激活脑岛, 脑岛可能是厌恶情绪加工的重要脑区, 但前脑岛与后脑岛存在明显的功能分离。
3.6 惊讶(Surprise)
根据fMRI研究的元分析, 惊讶诱发的大脑区域主要在皮层下, 包括杏仁核和纹状体, 以及一些皮层区域, 如腹内侧前额叶皮层和扣带皮层(Behrens et al., 2009; Bartra et al., 2013; Gu et al., 2019a)。Fouragnan等人(2018)发现了一个非常明显的惊讶编码网络的证据, 包括前扣带皮层、前岛叶和背侧纹状体。最新的研究表明, 基于价值的决策和知觉中的惊讶能够激活前脑岛(Loued- Khenissi al., 2020)。在奖励动机中, 惊讶增强了海马体的激活, 而在惩罚动机中, 惊讶增强了海马体旁皮质(parahippocampal cortex)的激活(Murty et al., 2016)。神经影像数据显示, 杏仁核反应可追踪惊讶面部表情的效价, 而不受唤醒度的干扰(Kim et al., 2017)。杏仁核、纹状体、扣带皮层等皮层下脑区可能参与了惊讶情绪的加工。
综上所述, 大量的脑影像数据都报告了特定基本情绪与特定脑区相关联, 不同刺激诱发的某一种基本情绪比较稳定地激活了一致的脑区。图3为脑影像学关于基本情绪的实验结果, 描述了假设与情绪类别相关的大脑区域:恐惧−杏仁核; 厌恶−脑岛; 愤怒−眶额皮质; 悲伤−前扣带回(Lindquist et al., 2012)。因此, 特定情绪的诱发存在稳定对应的脑区激活, 这或许可以为基本情绪理论提供实证支持。但是, 这些研究也提出了一些问题, 正如Barrett所提到的, 杏仁核几乎参与了所有的负性情绪, 比如恐惧、愤怒和惊讶。因此, 杏仁核很可能只是参与了唤醒度(arousal)的调节。相反, 我们认为导致这些研究不一致的原因是, 人类可能只有3种基本情绪:喜悦、恐惧(愤怒)、悲伤(图1)。恐惧和愤怒可能是一种基本情绪, 惊讶可能是诱发恐惧和愤怒情绪的认知成分, 它们有着共同的神经基础——杏仁核。有研究指出, 恐惧情绪可以促进愤怒情绪, 支持了情绪之间相互促进的假设:不同类型的情绪具有相互促进和抵消关系(Zhan et al., 2015, 2017); Zhan等人(2018)随后的fMRI进一步为恐惧情绪促进愤怒情绪的假设提供了脑影像证据。这也暗示了恐惧和愤怒是双胞胎情绪的假设(Gu et al., 2019a)。
4 应用机器视觉技术推进基本情绪脑影像研究
尽管大量研究证明了特定基本情绪与特定脑区相关联, 证明了某一基本情绪能够比较稳定地激活了某一特定脑区, 但并不是所有研究的都能得出一致的结果, 因此遭到了部分研究者的质疑(如Lindquist et al., 2012)。最新研究发现, 应用机器视觉技术有望推进基本情绪的脑影像研究, 可为基本情绪理论研究提供新思路和技术手段。
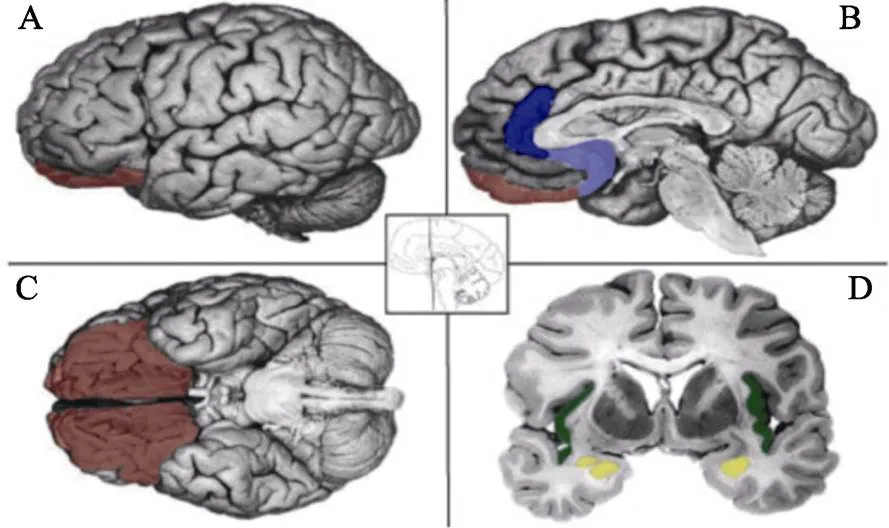
图3 脑影像学关于基本情绪的实验结果(Locationist Hypotheses of Brain–Emotion Correspondence) A: Lateral view. B: Sagital view at the midline. C: Ventral view. D: Coronal view. Brain regions hypothesized to be associated with emotion categories are depicted. Fear: amygdala (yellow); Disgust: insula (green); Anger: OFC (rust); Sadness: ACC (blue) (资料来源:Lindquist et al., 2012).
4.1 基本情绪脑影像研究遭遇的质疑
如上所述, 基本情绪研究的脑影像研究最近遇到了一些问题, 部分研究得到了不一致的结论。比如在面对恐惧或愤怒的面孔时, 杏仁核的反应没有群体差异(Cunha-Bang et al., 2019); 各种情绪面部表情均可能诱发正常人的杏仁核活动(Borgsted et al., 2018), 且杏仁核和脑岛都与情绪感知有关(Sokolov et al., 2020)。Saarimäki等人(2016)的fMRI研究结果显示, 基本情绪是由几个大脑区域内的离散神经信号支持的, 这一点可以从血液动力学大脑信号对情绪的高分类精度得到证明。Cowen等(2020)以音乐作为刺激材料的研究发现, 与音乐相关的情绪具有连续的梯度。换言之, 音乐诱发情绪的相关研究结果也不支持基本情绪理论。
Lindquist等(2012)对基本情绪的脑影像研究进行了元分析, 也发现不同研究的实验结果特异性很差, 难以确定独立的基本情绪与独特脑区相关。比如, 他们发现不同的基本情绪可能会激活相同的脑区, 杏仁核和几乎所有的负性情绪有关。他们研究发现与大脑其他区域相比, 恐惧体验并没有显示出杏仁核激活持续一致增加; 相反, 双侧杏仁核在厌恶体验时的激活相比于其他情绪有更一致的增加(Lindquist et al., 2012)。因此, 她们认为脑影像实验不能支持独特的基本情绪可以持续地、具体地定位到某个大脑特定区域的观点; 相反杏仁核可能与核心情绪的唤醒度(arousal)有关, 或者与不确定或意外的刺激有关(Gu et al., 2019a)。因此他们认为基本情绪的特定神经结构学说(独立的基本情绪一致且具体地对应于某个不同的大脑区域)应当让步于情绪维度学说(独立的情绪类别是由更一般的大脑网络构成的, 而不是特定于这些类别)。我们认为导致这些不一致的原因可能是由于人类只有3种基本情绪:喜悦、悲伤和恐惧(愤怒)。惊奇、恐惧和愤怒可能都是一种情绪, 因此他们都可以诱发杏仁核以及肾上腺素等交感神经反应, 也就是唤醒度(arousal)。
4.2 应用机器视觉技术推进基本情绪的脑影像研究
最近有研究者尝试应用机器视觉技术(machine-vision)对基本情绪的脑网络进行研究, 利用机器视觉技术解码情绪识别与分类; 尝试构建多种形式情绪原型, 探讨是否可以对多种形式的情绪原型共有特征和独特特征进行区分。为此, 我们设想可以通过不同脑区间的功能连接模式及脑网络特征研究, 从而确定3种基本情绪为基本情绪(Saarimäki et al., 2016)。或者说, 可以尝试从不同情绪的激活脑区网络中, 寻找几种基本情绪共同激活的脑区网络, 从而探讨某些基本情绪之间的紧密联系, 以便确定它们可能是一种基本情绪。如Pessoa (2017)就发现几种基本情绪的跨整个神经轴的网络交互作用; Satpute和Lindquist (2019)也指出默认模式网络(default mode network)能够构建某些基本情绪(愤怒、恐惧、厌恶等)之间的联系。
另外, 我们也可以从面部表情之间的联系来确定某些基本情绪之间的相关性, 从而确定这些相关情绪是否为一种同一个基本情绪。达尔文的面部表情研究为当今的表情研究提供了研究基础。一个人的情绪状态可以很容易地从其面部动作(即表情)推断出来(Barrett et al., 2019), 从达尔文的情绪研究开始, 大量的情绪研究都围绕表情展开, 如表情的分类与识别。Kragel等人(2019)通过fMRI研究证明了编码情绪类别的人类视觉皮层活动模式与模式输出相关, 并能解码多种情绪体验。也就是说, 丰富的、类别特定的视觉特征可以可靠地映射出不同的情绪, 并且它们在人类视觉系统中以分布式表示形式编码。因此, 可以在情绪识别的任务过程中, 利用机器视觉技术提取不同类型的情绪识别对应的视觉皮层特征, 通过对情绪编码和解码的聚类分析, 探索是否存在两者相对应的类别。
情绪识别与情绪产生的过程中, 眼睛的相关特征也会发生变化。情绪研究中记录fMRI的同时, 可借助兼容的眼动技术记录被试眼部相关特征的变化。特征提取是各种模式识别应用的主要阶段之一(Ghaffari et al., 2020), 我们也可以通过特征提取的办法应用于人脸及fMRI影像图。HOG (histogram of oriented gradients)是一种常用的特征提取算法, 在许多计算机视觉应用中被证明是有用的, 包括人类检测、汽车探测和一般的目标识别(Ghaffari et al., 2020)。如Dolensek等(2020)发表在Science 上的文章应用了机器学习技术, 将小鼠面部表情分为不同的类别, 反映了同一感官刺激在不同的稳态或情绪条件下所遇到的内在价值的变化。他们使用了HOG来处理实验鼠的面部特征, 以解读实验鼠的面部表情。利用机器视觉技术的量化特性, 可以证明面部表情揭示了情绪的核心属性, 如强度、持久性、灵活性和效价(Dolensek & Gogolla, 2021)。
基本情绪的脑影像研究结合机器视觉技术可能是确定基本情绪的数目的最佳途径。通过对几种基本情绪诱发前后的人类面部表情特征及fMRI影像图的网络特征分别进行处理分析, 形成HOG向量描述符, 观察某些不同基本情绪是否可以归为一类。接着通过主成分分析(principal component analysis, PCA)和t-分布随机邻居嵌入法(t-distributed stochastic neighbor embedding, t- SNE)来降低数据维度, 将面部表情分解成离散的情绪; 训练一个随机森林分类器(random forest classifier, RFC)以区分不同面部表情(Dolensek & Gogolla, 2021)。构建几种基本情绪的人类面孔及fMRI影像情绪原型, 对比其他被试基本情绪fMRI影像图特征与相应情绪原型的相似性, 从而确定某些基本情绪的关系。由此可检验情绪原型的充分性(Dolensek et al., 2020)。由于fMRI成像以图形形式呈现, 使我们通过HOG向量的处理技术来构建基本情绪的fMRI影像图成为可能。未来研究可以聚焦多种形式(如上述的行为、生理、视觉皮层、眼动、面部表情、fMRI影像图、EEG地形图等数据特征)的共同特征与独特特征、特征间的连接(如脑功能连接), 考虑从多个角度尝试构建情绪原型, 有望形成多角度、多种形式的情绪原型, 为基本情绪理论研究提供新思路。
5 总结
基本情绪理论历经几十年的发展, 取得了众多研究成果。基本情绪理论的观点在情绪科学领域和相关学科得到广泛认可与应用, 对于人类的生存生活、信息沟通及心身疾病的诊疗也至关重要(Liang et al., 2021)。许多基本情绪脑影像及元分析研究支持了基本情绪理论。但是, 基本情绪的种类依然存在争议。比较经典的基本情绪分类是Ekman的6种基本情绪分类:高兴, 愤怒, 恐惧, 惊奇, 厌恶, 悲伤。数目最少的是我们最近研究提出的3种基本情绪观点, 认为人类可能只有3种基本情绪:喜悦、悲伤、恐惧(愤怒)。大量的fMRI研究也支持我们的观点, 比如许多fMRI研究发现:恐惧和愤怒都与杏仁核有关, 惊讶也与杏仁核、扣带皮层等下皮层脑区相关。然而, 这些结果还没有结束基本情绪理论与情绪维度理论的争论。多学科交叉融合并考虑将机器视觉技术应用于基本情绪脑影像研究或许是一个富有成效的探索路线, 有望为基本情绪理论提供可信度较高的证据。
梁飞, 李红, 王福顺. (2021). 基本情绪的电生理活动: 来自ERP的证据.(1), 42−56.
王福顺, 成敬, 张艳萍, 姜长青. (2018).. 北京: 人民卫生出版社.
An, S., Han, X., Wu, B., Shi, Z., Marks, M., Wang, S., Wu, X., & Han, S. (2018). Neural activation in response to the two sides of emotion., 140–144.
Anthony, T. E., Dee, N., Bernard, A., Lerchner, W., Heintz, N., & Anderson, D. J. (2014). Control of stress-induced persistent anxiety by an extra-amygdala septohypothalamic circuit.,(3), 522–536.
Arnold, M. B. (1960).. New York: Columbia University Press.
Bartra, O., McGuire, J. T., & Kable, J. W. (2013). The valuation system: A coordinate-based meta-analysis of BOLD fMRI experiments examining neural correlates of subjective value., 412–427.
Barrett, L. F., Adolphs, R., Marsella, S., Martinez, A. M., & Pollak, S. D. (2019). Emotional expressions reconsidered: Challenges to inferring emotion from human facial movements.(1), 1–68.
Barrett, L. F., Khan, Z., Dy, J., & Brooks, D. (2018). Nature of emotion categories: Comment on Cowen and Keltner.(2), 97–99.
Behrens, T. E., Woolrich, M. W., Walton, M. E., & Rushworth, M. F. (2009). Learning the value of information in an uncertain world., 1214–1221.
Bogert, B., Numminen-Kontti, T., Gold, B., Sams, M., Numminen, J., Burunat, I., Lampinen, J., & Brattico, E. (2016). Hidden sources of joy, fear, and sadness: Explicit versus implicit neural processing of musical emotions., 393–402.
Borgsted, C., Ozenne, B., Mc Mahon, B., Madsen, M. K., Hjordt, L. V., Hageman, I., ... Fisher, P. M. (2018). Amygdala response to emotional faces in seasonal affective disorder., 288–295.
Britton, J. C., Taylor, S. F., Sudheimer, K. D., & Liberzon, I. (2006). Facial expressions and complex IAPS pictures: Common and differential networks.(2), 906–919.
Buades-Rotger, M., & Krämer, U. M. (2018). From words to action: Implicit attention to antisocial semantic cues predicts aggression and amygdala reactivity to angry faces in healthy young women.(6), 624–637.
Celeghin, A., Diano, M., Bagnis, A., Viola, M., & Tamietto, M. (2017). Basic emotions in human neuroscience: Neuroimaging and beyond., 1432–1432.
Chapman, H. A., & Anderson, A. K. (2012). Understanding disgust.(1), 62–76.
Clore, G. L., & Ortony, A. (2013). Psychological construction in the OCC model of emotion.(4), 335–343.
Coccaro, E. F., McCloskey, M. S., Fitzgerald, D. A., & Phan, K. L. (2007). Amygdala and orbitofrontal reactivity to social threat in individuals with impulsive aggression.,(2), 168–178.
Cowen, A. S., Fang, X., Sauter, D., & Keltner, D. (2020). What music makes us feel: At least 13 dimensions organize subjective experiences associated with music across different cultures.(4), 1924–1934.
Cowen, A. S., & Keltner, D. (2017). Self-report captures 27 distinct categories of emotion bridged by continuous gradients.(38), E7900–E7909.
da Cunha-Bang, S., Fisher, P. M., Hjordt, L. V., Holst, K., & Knudsen, G. M. (2019). Amygdala reactivity to fearful faces correlates positively with impulsive aggression.(2), 162–172.
DeLancey, C. (2002).. Oxford: Oxford University Press.
Dolensek, N., Gehrlach, D. A., Klein, A. S., & Gogolla, N. (2020). Facial expressions of emotion states and their neuronal correlates in mice.(6486), 89-94.
Dolensek, N., & Gogolla, N. (2021). Machine-learning approaches to classify and understand emotion states in mice.(1), 250–251.
Dzafic, I., Oestreich, L., Martin, A. K., Mowry, B., & Burianová, H. (2019). Stria terminalis, amygdala, and temporoparietal junction networks facilitate efficient emotion processing under expectations.(18), 5382–5396.
Ekman, P. (1972).. New York, NY: Pergamon Press.
Ekman, P. (1992). An argument for basic emotions?, 169–200.
Ekman, P. (1999). Basic emotions. In T. Dalgleish & M. J. Power(Eds).(pp.45–60)Chichester: John Wiley and Sons.
Ekman, P., & Davidson, R. J. (Eds.). (1994).. New York, NY: Oxford University Press.
Ekman, P., Friesen, W. V., & Ellsworth, P. (1982). What emotion categories or dimensions can observers judge from facial behavior? In P. Ekman (Ed.),(pp. 39−55). New York: Cambridge University Press.
Ekman, P., Sorenson, E. R., & Friesen, W. V. (1969). Pan- cultural elements in the facial display of emotions., 86–88.
Fouragnan, E., Retzler, C., & Philiastides, M. G. (2018). Separate neural representations of prediction error valence and surprise: Evidence from an fMRI meta-analysis.(7), 2887–2906.
Frijda, N. H. (1987). Comment on Oatley and Johnson- Lairdʼs “Towards a cognitive theory of emotions.”, 51–58.
Fulwiler, C. E., King, J. A., & Zhang, N. (2012). Amygdala- orbitofrontal resting-state functional connectivity is associated with trait anger.,(10), 606–610.
Ghaffari, S., Soleimani, P., Li, K. F., & Capson, D. W. (2020). A novel hardware-software co-design and implementation of the HOG algorithm.(19), 5655.
Graham, A. M., Buss, C., Rasmussen, J. M., Rudolph, M. D., Demeter, D. V., Gilmore, J. H., ... Fair, D. A. (2016). Implications of newborn amygdala connectivity for fear and cognitive development at 6-months-of-age., 12–25.
Gray, J. A. (1982).. Oxford: Oxford University Press.
Griffiths, P. E. (1997).. Chicago, IL:University of Chicago Press.
Gu, S., Gao, M., Yan, Y., Wang, F., Tang, Y.-Y., & Huang, J. H. (2018). The neural mechanism underlying cognitive and emotional processes in creativity., 1924.
Gu, S., Wang, F., Cao, C., Wu, E., Tang, Y.-Y., & Huang, J. H. (2019a). An integrative way for studying neural basis of basic emotions with fMRI., 628–628.
Gu, S., Wang, F., Patel, N. P., Bourgeois, J. A., & Huang, J. H. (2019b). A model for basic emotions using observations of behavior in drosophila., 781.
Gu, S., Wang, W., Wang, F., & Huang, J. H. (2016). Neuromodulator and emotion biomarker for stress induced mental disorders., 2609128.
Han, W., Tellez, L. A., Rangel, M. J., Jr, Motta, S. C., Zhang, X., Perez, I. O., ... de Araujo, I. E. (2017). Integrated control of predatory hunting by the central nucleus of the amygdala.,(1–2), 311–324.e18.
Hutto, D. D., Robertson, I., & Kirchhoff, M. D. (2018). A new, better BET: Rescuing and revising basic emotion theory., 1217–1217.
Isosaka, T., Matsuo, T., Yamaguchi, T., Funabiki, K., Nakanishi, S., Kobayakawa, R., & Kobayakawa, K. (2015). Htr2a- Expressing cells in the central amygdala control the hierarchy between innate and learned fear.,(5), 1153–1164.
Izard, C. E. (1971).. Appleton-Century- Crofts.
Jack, R. E., Garrod, O., & Schyns, P. G. (2014). Dynamic facial expressions of emotion transmit an evolving hierarchy of signals over time.(2), 187–192.
Jack, R. E., Sun, W., Delis, I., Garrod, O. G., & Schyns, P. G. (2016). Four not six: Revealing culturally common facial expressions of emotion.(6), 708–730.
James, W. (1884). What is an emotion?, 188–205.
Kapucu, A., Kılıç, A., Özkılıç, Y., & Sarıbaz, B. (2021). Turkish emotional word norms for arousal, valence, and discrete emotion categories.(1), 188–209.
Keltner, D., Sauter, D., Tracy, J., & Cowen, A. (2019). Emotional Expression: Advances in basic emotion theory.(2), 133–160.
Keltner, D., Tracy, J. L., Sauter, D., & Cowen, A. (2019). What basic emotion theory really says for the twenty-first century study of emotion.,(2), 195–201.
Kim, M. J., Mattek, A. M., Bennett, R. H., Solomon, K. M., Shin, J., & Whalen, P. J. (2017). Human amygdala tracks a feature-based valence signal embedded within the facial expression of surprise.(39), 9510–9518.
Kluczniok, D., Hindi Attar, C., Stein, J., Poppinga, S., Fydrich, T., Jaite, C., ... Bermpohl, F. (2017). Dissociating maternal responses to sad and happy facial expressions of their own child: An fMRI study.(8), e0182476.
Kragel, P. A., Reddan, M. C., LaBar, K. S., & Wager, T. D. (2019). Emotion schemas are embedded in the human visual system.(7), eaaw4358.
Lane, R. D., Reiman, E. M., Ahern, G. L., Schwartz, G. E., & Davidson, R. J. (1997). Neuroanatomical correlates of happiness, sadness, and disgust.(7), 926–933.
LeDoux, J., Phelps, L., & Alberini, C. (2016). What we talk about when we talk about emotions.(6), 1443–1445.
Levenson, R. W. (2003). Blood, sweat, and fears.(1), 348–366.
Liang, F., Feng, R., Gu, S., Jiang, S., Zhang, X., Li, N., ... Wang, F.. (2021). Neurotransmitters and electrophysiological changes might work as biomarkers for diagnosing affective disorders., 9116502.
Lindquist, K. A., Siegel, E. H., Quigley, K. S., & Barrett, L. F. (2013). The hundred-year emotion war: Are emotions natural kinds or psychological constructions? Comment on Lench, Flores, and Bench (2011).(1), 255–263.
Lindquist, K. A., Wager, T. D., Kober, H., Bliss-Moreau, E., & Barrett, L. F. (2012). The brain basis of emotion: A meta-analytic review.(3), 121–143.
Loued-Khenissi, L., Pfeuffer, A., Einhäuser, W., & Preuschoff, K. (2020). Anterior insula reflects surprise in value-based decision-making and perception., 116549.
McDougall, W. (1926).. Boston: Luce.
Mowrer, O. H. (1960).New York: Wiley.
Murty, V. P., LaBar, K. S., & Adcock, R. A. (2016). Distinct medial temporal networks encode surprise during motivation by reward versus punishment.(Pt A), 55–64.
Oaten, M., Stevenson, R. J., Williams, M. A., Rich, A. N., Butko, M., & Case, T. I. (2018). Moral violations and the experience of disgust and anger., 179.
Oatley, K., & Johnson-Laird, P. N. (1987). Towards a cognitive theory of emotions., 29–50.
Ortony, A., & Turner, T. J. (1990). Whatʼs basic about basic emotions?(3), 315–331.
Panksepp, J. (1982). Toward a general psychobiological theory of emotions.(3), 407–467.
Pessoa L. (2017). A network model of the emotional brain.(5), 357–371.
Phan, K. L., Wager, T., Taylor, S. F., & Liberzon, I. (2002). Functional neuroanatomy of emotion: A meta-analysis of emotion activation studies in PET and fMRI.(2)331–348.
Plutchik, R. (1980). A general psychoevolutionary theory of emotion. In R. Plutchik & H. Kellerman (Eds.),(pp. 3−31). New York: Academic Press.
Rahko, J., Paakki, J.-J., Starck, T., Nikkinen, J., Remes, J., Hurtig, T., ... Kiviniemi, V. (2010). Functional mapping of dynamic happy and fearful facial expression processing in adolescents.(2), 164–176.
Ramirez-Mahaluf, J. P., Perramon, J., Otal, B., Villoslada, P., & Compte, A. (2018). Subgenual anterior cingulate cortex controls sadness-induced modulations of cognitive and emotional network hubs.(1), 8566.
Reynaud, E., Guedj, E., Trousselard, M., El Khoury- Malhame, M., Zendjidjian, X., Fakra, E., ... Khalfa, S. (2015). Acute stress disorder modifies cerebral activity of amygdala and prefrontal cortex.,(1), 39–43.
Russell, J. A. (1980). A circumplex model of affect.(6), 1161–1178.
Saarimäki, H., Gotsopoulos, A., Jääskeläinen, I. P., Lampinen, J., Vuilleumier, P., Hari, R., ... Nummenmaa, L. (2016). Discrete neural signatures of basic emotions.(6), 2563–2573.
Satpute, A. B., & Lindquist, K. A. (2019). The default mode networkʼs role in discrete emotion.(10), 851–864.
Scarantino, A., & Griffiths, P. (2011). Don’t give up on basic emotions.(4), 444–454.
Schwartz, J., Ordaz, S. J., Kircanski, K., Ho, T. C., Davis, E. G., Camacho, M. C., & Gotlib, I. H. (2019). Resting-state functional connectivity and inflexibility of daily emotions in major depression., 26–34.
Sokolov, A. A., Zeidman, P., Erb, M., Pollick, F. E., Fallgatter, A. J., Ryvlin, P., Friston, K. J., & Pavlova, M. A. (2020). Brain circuits signaling the absence of emotion in body language.(34), 20868– 20873.
Song, Y., & Hakoda, Y. (2018). Selective impairment of basic emotion recognition in people with autism: Discrimination thresholds for recognition of facial expressions of varying intensities.(6), 1886–1894.
Suardi, A., Sotgiu, I., Costa, T., Cauda, F., & Rusconi, M. (2016). The neural correlates of happiness: A review of PET and fMRI studies using autobiographical recall methods.(3), 383–392.
Thomas, E., Buss, C., Rasmussen, J. M., Entringer, S., Ramirez, J. S. B., Marr, M., ... Graham, A. M. (2019). Newborn amygdala connectivity and early emerging fear., 100604.
Tomkins, S. S. (1984). Affect theory. In K. R. Scherer & P. Ekman (Eds.),(pp. 163–195). Hillsdale, NJ: Erlbaum.
Tonnaer, F., Siep, N., van Zutphen, L., Arntz, A., & Cima, M. (2017). Anger provocation in violent offenders leads to emotion dysregulation.(1), 3583.
Touroutoglou, A., Lindquist, K. A., Dickerson, B. C., & Barrett, L. F. (2015). Intrinsic connectivity in the human brain does not reveal networks for 'basic' emotions.(9), 1257–1265.
Vetter, N. C., Drauschke, M., Thieme, J., & Altgassen, M. (2018). Adolescent basic facial emotion recognition is not influenced by puberty or own-age bias., 956.
Viol, K., Aas, B., Kastinger, A., Kronbichler, M., Schöller, H. J., Reiter, E.-M., ... Schiepek, G. K. (2019). Erroneously disgusted: fMRI study supports disgust-related neural reuse in obsessive-compulsive disorder (OCD)., 81.
Vytal, K., & Hamann, S. (2010). Neuroimaging support for discrete neural correlates of basic emotions: A voxel-based meta-analysis.(12), 2864–2885.
Wang, F., Pan, F., Shapiro, L. A., & Huang, J. H. (2017). Stress induced neuroplasticity and mental disorders., 9634501.
Wang, H., Chen, Y., & Zhang, Q.. (2018). The effects of low and high levels of sadness on scope of attention: An ERP study., 2397–2397.
Wang, Y., Kong, F., Kong, X., Zhao, Y., Lin, D., & Liu, J. (2017). Unsatisfied relatedness, not competence or autonomy, increases trait anger through the right amygdala.(5), 932–938.
Watson, J. B. (1930).. Chicago: University of Chicago Press.
Weiner, B., & Graham, S. (1984). An attributional approach to emotional development. In C. E. Izard, J. Kagan, & R. B. Zajonc (Eds.),(pp. 167–191). New York: Cambridge University Press.
Williams, R. (2017). Anger as a basic emotion and its role in personality building and pathological growth: The neuroscientific, developmental and clinical perspectives., 1950.
Wilson-Mendenhall, C. D., Barrett, L. F., & Barsalou, L. W. (2013). Neural evidence that human emotions share core affective properties.(6), 947–956.
Wright, P., He, G., Shapira, N. A., Goodman, W. K., & Liu, Y. (2004). Disgust and the insula: FMRI responses to pictures of mutilation and contamination.(15), 2347–2351.
Xiao, M., Zhu, W., Wei, J., Lei, X., & Xia, L.-X. (2019). The relationship among resting-state brain activity and connectivity, agreeableness and displaced aggression: Two possible mediation models., 641–649.
Ying, X., Luo, J., Chiu, C.-Y., Wu, Y., Xu, Y., & Fan, J. (2018). Functional dissociation of the posterior and anterior insula in moral disgust., 860.
Zhan, J., Ren, J., Fan, J., & Luo, J. (2015). Distinctive effects of fear and sadness induction on anger and aggressive behavior., 725.
Zhan, J., Ren, J., Sun, P., Fan, J., Liu, C., & Luo, J. (2018). The Neural Basis of Fear Promotes Anger and Sadness Counteracts Anger., 3479059.
Zhan, J., Tang, F., He, M., Fan, J., Xiao, J., Liu, C., & Luo, J. (2017). Regulating Rumination by Anger: Evidence for the Mutual Promotion and Counteraction (MPMC) Theory of Emotionality., 1871.
Zheng, Z., Gu, S., Lei, Y., Lu, S., Wang, W., Li, Y., & Wang, F. (2016). Safety needs mediate stressful events induced mental disorders., 8058093.
Ziegler, J. C., Montant, M., Briesemeister, B. B., Brink, T. T., Wicker, B., Ponz, A., ... Braun, M. (2018). Do words stink? Neural reuse as a principle for understanding emotions in reading.(7), 1023–1032.
Basic emotion and its neural basis: Evidence from fMRI and machine-vision studies
LIANG Fei, JIANG Yao, XIAO Tingwei, DONG Jie, WANG Fushun
(Institute of Brain and Psychological Science, Sichuan Normal University, Chengdu 610066, China)
Basic emotion theory is the most representative theory in the field of emotion science, which holds that human emotions are composed of a limited number of basic emotions, such as fear, anger, joy, sadness and so on. Basic emotions are evolved for fundamental life tasks, and each basic emotion has its own unique neural structure and physiological basis.Although basic emotion theories are widely accepted, there is little agreement on specific basic emotions. In recent decades, many fMRI studies have attempted to determine the unique neural structural underpinnings of each of the basic emotions, and made many important findings, such as disgust is related to the insula, sadness is related to the anterior cingulate gyrus, and the amygdala is an important limbic structure related to fear. However, recent meta-analysis studies have found that many basic emotions share many brain regions. Therefore, the theory of specific brain regions of basic emotions has been questioned and even denied. Through the discussion of the basic emotions and neural basis, the new functional magnetic resonance imaging (fMRI) study of basic emotion theory were summarized, and put forward the basic theory of emotional debate comes from the determination of basic emotion type, because many of the so-called different basic emotion is actually the same basic emotions, possible only three basic human emotions. In the future, machine-visioncan be used to further promote the research of basic emotion brain imaging.
basic emotion, emotional dimension, fMRI, machine-vision, brain imaging
2021-07-08
* 教育部人文社会科学基金(19YJAZH083)
王福顺, E-mail: 13814541138@163.com
B842; B845
