长白山木本植物组成及随海拔变化规律研究
2022-08-08李仕吉
李仕吉
(吉林省林业勘察设计研究院, 吉林 长春 130022)
木本植物资源负载着森林植物的遗传多样性及基因资源,关系到可持续发展和将来基因工程的基本保障,是国家重大的战略资源,同时也是国家自然科技资源必不可少的基础物质条件。木本植物资源信息与实物的分享与共享,是国家自然科技资源平台建设的重要内容,也是国家科技体系建设及科技创新的内容,更是一项重要的基础性工作。
人类利用和转移木本植物资源由来已久,有上百万年历史。例如18世纪,在整个欧洲大陆,欧洲赤松(Pinussylvestris)、挪威云杉(Piceaabies)、欧洲落叶松(Larixdecidua)及橡树(Quercusspp.)等木本植物的种子被广泛交易。19世纪中后期至20世纪中期,木本植物种质资源的转移是为了提高当地景观的休闲娱乐价值和相应的经济收益。在20世纪的大部分时间里,木本植物的经济价值得到进一步重视。
随着我国经济的发展,人们日益增长的物质需求对木本植物开发提出了更高的要求,同时,公众环境保护意识的增强以及环境破坏的客观现实要求我们必须格外重视木本植物资源的调查、保存和保育工作。我国在21世纪初陆续开展了各种层次和规模的野生植物资源普查,重点开展了林木种质资源的调查,这些调查研究在各省的进度不一,取得的研究成果也相对粗放,尤其是信息标准化和大数据整合方面十分欠缺[1]。因此,国家林草局以及相关部门要求在彻底普查各省(市)和重点林区林木种质资源的基础上,进行数据整合,同时根据信息保密程度不同,开展相应的共享和服务。
长白山是我国东北地区的生态屏障,也是林木资源最丰富的地区之一。开展长白山木本植物资源调查研究,是吉林省乃至全国林木事业的基础,具有重要意义。
1 研究区概况
研究区位于吉林省东南部长白山的主脉和南岗岭以西,龙岗山脉以东,英额岭以南,鸭绿江和图们江以北的广大地域,主要包括长白、抚松、安图、和龙等县的部分地区,总面积约10 000 km2。
该区植被属长白山植物区系,主要植被类型为天然次生针阔混交林。辖区内森林资源受到长期的保护和合理的经营与管理,林木相对保存完整,物种丰富度和多样性都保持在较高水平。森林群落内垂直成层现象明显,乔木树种主要包括红松(Pinuskoraiensis)、春榆(Ulmusjaponica)、色木槭(Acermono)、核桃楸(Juglansmandshurica)、水曲柳(Fraxinusmandshurica)、紫椴(Tiliaamurensis)、白牛槭(Acermandshuricum)、蒙古栎(Quercusmongolica)、大青杨(Populusussuriensis)、裂叶榆(Ulmuslaciniata)、怀槐(Maackiaamurensis)等。灌木主要包括簇毛槭(Acerbarbinerve)、暴马丁香(Syringareticulata)、毛榛(Corylusmandshurica)、瘤枝卫矛(Euonymuspauciflorus)、东北鼠李(Rhamnusdavurica)、金银忍冬(Loniceramaackii)等。林下草本植物丰富,在不同季节表现出不同的林下景观,常见的草本植物有黑水银莲花(Anemoneamurensis)、多被银莲花(Anemoneraddeana)、朝鲜顶冰花(Gagealuteavar.nakaiana)、荷青花(Hylomeconjaponicum)、侧金盏(Adonisamurensis)、美汉草(Meehaniafargesii)、北重楼(Parisverticillata)、轮叶百合(Liliumdistichum)、白花碎米荠(Cardamineleucantha)、山茄子(Anisodusacutangulus)、蚊子草(Filipendulapalmata)、鹿药(Smilacinajaponica)、紫花变豆菜(Sanicularubriflora)、东北羊角芹(Aegopodiumalpestre)、荨麻(Urticafissa)、宽叶苔草(Carexsiderosticta)等[2]。
正是由于长白山的特殊地形地貌和独特的地理位置,孕育了近2 000种植物,树木种类占10 %以上[3]。历年以来,学者们相继在长白山开展了各种经济树种的专项研究以及各物种共存机制的生态调查,但是对长白山各种珍稀濒危植物的资源保有量,至今还没有较为详细和全面的普查结果,为此,进行长白山木本植物的研究具有重要意义。
2 调查方法
利用《中国树木志》《吉林省林木种质资源调查初列树种名录》、森林资源规划调查设计和其他有关林业资料,结合访问、座谈等方式,首先查清调查区域内木本植物树种的集中分布范围和面积,然后在其中选择有代表性的林分,设标准地进行典型调查。对通过上述方法不能确定树种集中分布区的,可通过线路踏查的方式确定其集中分布区,然后在其中选择有代表性的林分,设标准地进行调查。
2.1 线路踏查
2.1.1 线路踏查确定原则
充分考虑区域内的自然条件和林分特点,选择森林植被垂直分布比较复杂的地段,尽可能包括所有植物群落,并且具备土壤、植被代表性强的特点。
2.1.2 踏查线路的设置要求
先在地形图或卫星图片上设计踏查线路,线路密度根据自然条件的复杂程度和植物群落的类型来确定。
在长白山北坡、西坡、南坡各个坡面地段,踏查线路要与主山脊的分水岭走向垂直,从谷底向山脊沿海拔升高的方向设置;在河谷地段,踏查线路沿河岸由下游向上游设置。
2.2 典型调查
典型调查林分要优先选择调查树种集中分布的林地;优先选择符合优良林分要求的林地;优先选择天然林;优先选择纯林。
典型调查采取标准地调查方式。标准地形状为正方形,面积0.06 hm2。
标准地设置可根据树种、分布范围、地形地貌等情况而定,不能跨越河流、道路或伐开的调查线,且远离林缘。
3 研究方法
3.1 植物科属组成
对研究区内175个样方调查获取的乔木和灌木种类数据,进行科属分类,并统计其科属组成。
3.2 海拔梯度设置
研究区内175个样方分布在海拔440~2 045 m,按照海拔<900 m、900 m≤海拔≤1 100 m、1 100 m<海拔≤1 300 m、1 300 m<海拔≤1 500 m和海拔>1 500 m将所有样方分成5个梯度。根据不同海拔梯度分析不同海拔梯度范围内物种组成及其各物种重要值的变化。
3.3 不同海拔多样性指数
对研究区内175个样方按照其海拔梯度进行分类,由于175个样方中有2个样方的调查数据不完整,在进行多样性计算时未包括在内,只对173个样方进行统计分析。5个海拔梯度范围内样方数量分别为58个、78个、15个、13个和9个。计算不同海拔梯度范围内乔木、灌木以及乔木、灌木整合后群落的物种数量(S)、Shannon-winner指数(H)、Simpson指数(D)和Pielou均匀度指数(J)。
采用Microsoft Office Excel 2007对数据进行整理和分析,用SPSS 18.0软件进行one-way ANOVA(单因素方差分析),用LSD(最小显著差异法)进行多重比较(P<0.05水平下进行)。
4 结果与讨论
4.1 植物的科属组成
对海拔440~2 045 m范围内175个样方的物种组成进行调查,共调查到木本植物102种,分属25科51属。其中裸子植物4科6属9种,被子植物21科45属93种。乔木42种,13科21属;灌木59种,16科31属;另有藤本植物1种(五味子)。
乔木层中以落叶阔叶树种为主,共34种,占全部乔木树种的80.9 %;针叶树种8种,占全部乔木树种的19.1 %。灌木层中共调查到灌木树种59种,占全部调查区木本树种的57.8 %。
从属、种的组成来看,各科所含属的数量有明显差别,其中大于10个属的仅有1科即蔷薇科,包含13个属,占全属的25.5 %,含有18个种,占全种的17.7 %;含有2~4个属的有8个科,分别为松科(4属,7种)、忍冬科(3属,12种)、杜鹃花科(3属,7种)、虎耳草科(3属,7种)、桦木科(3属,7种)、杨柳科(2属,5种)、木犀科(2属,4种)和卫矛科(2属,3种)。以上9科占全科的36 %,包含35属,占全属的68.6 %,包含70种,占全部种的68.6 %;其余科都为单属科,共计16科,占全科的64 %。
4.2 不同海拔梯度物种组成
海拔<900 m梯度内木本植物组成为72种,分属21科38属。其中乔木33种,灌木38种,藤本1种。
900 m≤海拔≤1 100 m梯度内木本植物组成为69种,分属21科38属。其中乔木33种,灌木36种。
1 100 m<海拔≤1 300 m梯度内木本植物组成为45种,分属19科29属。其中乔木22种,灌木23种。
1 300 m<海拔≤1 500 m梯度内木本植物组成为46种,分属16科28属。其中乔木19种,灌木27种。
海拔>1 500 m梯度内木本植物组成为15种,分属7科11属。其中乔木4种,分属3科4属。
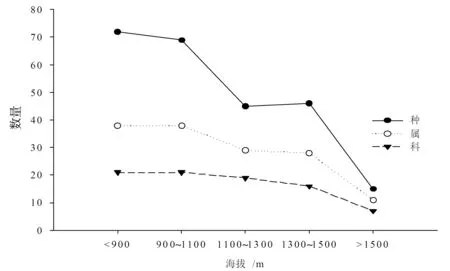
图1 物种科、属、种随海拔梯度变化趋势
物种科、属、种随海拔梯度变化趋势见图1。从图1中可以看出,随海拔梯度升高,物种科、属、种数量有逐渐降低的趋势,其中在海拔<900 m和900 m≤海拔≤1 100 m梯度物种科、属、种的数量无显著变化;而1 100 m<海拔≤1 300 m较900 m≤海拔≤1 100 m梯度物种属和种的数量显著降低,而科的数量降低趋势不明显;随着海拔升高,从1 100 m<海拔≤1 300 m到1 300 m<海拔≤1 500 m梯度物种属和种的数量无明显变化,而科的数量有稍降低趋势;海拔>1 500 m时,物种科、属、种的数量急剧降低。从以上可以看出,物种科、属、种的数量分布受到海拔的影响,其中对种数量的影响最为明显,特别是到高海拔区域,受到温度的影响,物种数量分布极少。
4.3 不同海拔物种多样性指数
4.3.1 乔木多样性指数随海拔的变化
由于有2个样方的调查数据不完整,在进行多样性计算时未包括在内,只对173个样方进行了统计分析,结果见表1。
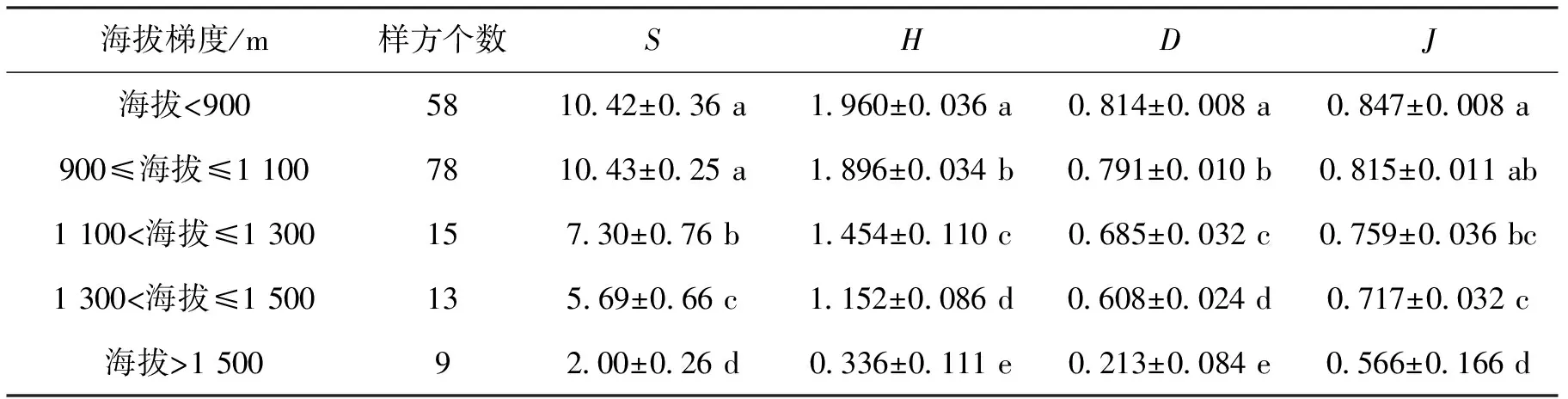
表1 不同海拔梯度乔木多样性指数
从表1中可以看出,乔木层中,在海拔<900 m和900 m≤海拔≤1 100 m 2个海拔梯度内物种数量无明显变化,当海拔>1 100 m,随海拔升高物种数量有显著降低的趋势(P<0.05);Shannon-winner多样性指数和Simpson指数均随海拔梯度的升高逐渐降低,且不同海拔范围内均差异显著(P<0.05);Pielou均匀度指数同样随海拔升高逐渐降低,除1 100 m<海拔≤1 300 m和1 300 m<海拔≤1 500 m梯度无显著差异外,其他海拔梯度均差异显著。整体来看,海拔梯度显著影响了乔木层物种多样性指标的变化,随海拔梯度的升高呈现逐渐降低的趋势。
4.3.2 灌木多样性指数随海拔的变化
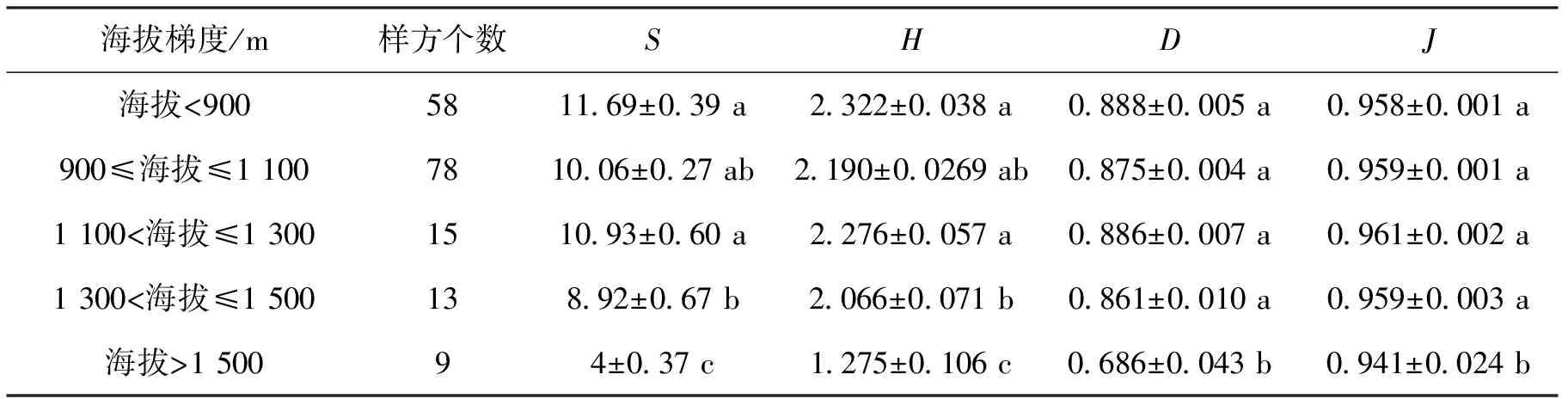
表2 不同海拔梯度灌木多样性指数
不同海拔梯度灌木多样性指数的变化见表2。可以看出,在海拔<900 m时,具有最高的物种数量(11.69),随后降低,从海拔≥900 m开始,随海拔梯度的升高,物种数量呈先增大后显著降低趋势;Shannon-winner多样性指数和Simpson多样性指数与之有相似的规律,相比而言,Shannnon-winner指数对灌木多样性指数随海拔变化趋势的敏感性更强;灌木Pielou均匀度指数除海拔>1 500 m显著低于其他海拔梯度外,其他各海拔梯度无显著差异。
4.3.3 乔木和灌木整体多样性指数随海拔的变化
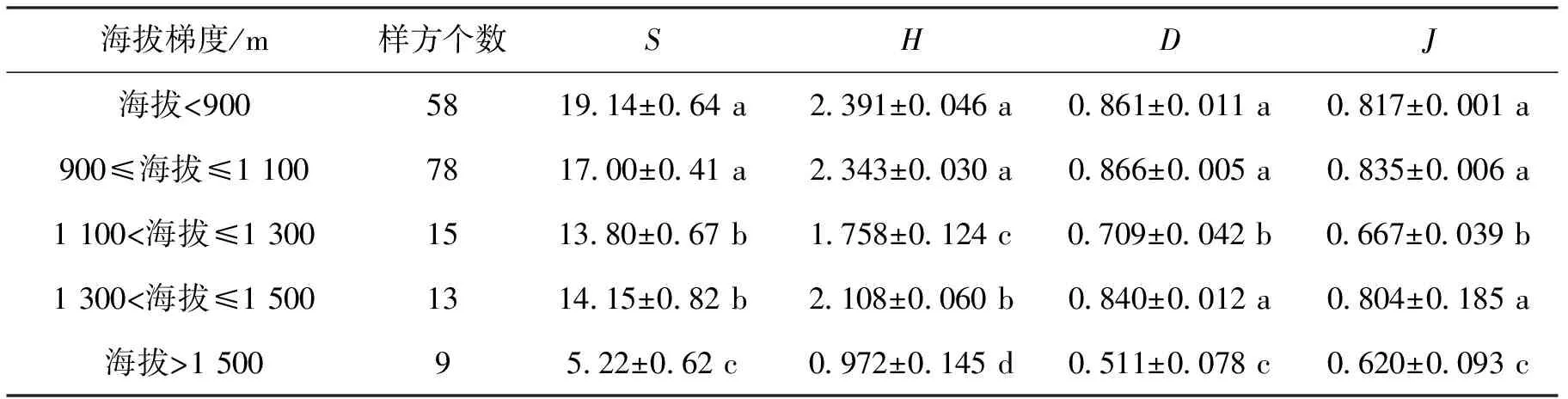
表3 不同海拔梯度乔木和灌木整体群落多样性指数
不同海拔梯度乔木和灌木整体群落多样性指数见表3。可以看出,物种数量在海拔<900 m和900 m≤海拔≤1 100 m梯度内无显著差异,但显著大于其他海拔梯度,1 100 m<海拔≤1 300 m和1 300 m<海拔≤1 500 m梯度内无显著差异,显著大于海拔>1 500 m梯度(P<0.05);Shannon-winner多样性指数表现为海拔<900 m和900 m≤海拔≤1 100 m梯度间无显著差异,显著大于其他各海拔梯度,其次为1 300 m<海拔≤1 500 m梯度显著大于1 100 m<海拔≤1 300 m和海拔>1 500 m梯度,同时1 300 m<海拔≤1 500 m梯度显著大于海拔>1 500 m梯度。Simpson多样性指数表现为海拔<900 m、900 m≤海拔≤1 100 m和1 300 m<海拔≤1 500 m梯度无显著差异,但显著高于1 100 m<海拔≤1 300 m和海拔>1 500 m梯度,仍以海拔>1 500 m梯度时最低。Pielou均匀度指数随各海拔梯度的变化与Simpson指数有相似的规律。从以上可以看出,乔木和灌木群落多样性指数的变化随海拔的升高呈先逐渐下降,至1 300 m<海拔≤1 500 m梯度时升高,后又逐渐下降的趋势。
5 结论
研究区调查到木本植物102种,分属25科,51属,其中裸子植物4科6属9种,被子植物21科45属93种。乔木42种,13科21属;灌木59种,16科31属;另有藤本植物1种(五味子)。
不同海拔梯度中的物种组成为海拔<900 m梯度内72种,分属21科38属。其中乔木33种,灌木38种,藤本1种;900 m≤海拔≤1 100 m梯度内69种,分属21科38属。其中乔木33种,灌木36种;1 100 m<海拔≤1 300 m梯度内45种,分属19科29属。其中乔木22种,灌木23种;1 300 m<海拔≤1 500 m梯度内46种,分属16科28属。其中乔木19种,灌木27种;海拔>1500 m梯度内15种,分属7科11属。其中乔木4种,分属3科4属。
不同海拔梯度乔木层中,在海拔<900 m和900 m≤海拔≤1 100 m 2个梯度内物种数量无明显变化,当海拔>1 100 m后,随海拔升高物种数量有显著降低的趋势(P<0.05);Shannon-winner多样性指数和Simpson指数均随海拔梯度的升高逐渐降低,且不同海拔梯度间均差异显著(P<0.05);Pielou均匀度指数同样随海拔升高呈逐渐降低趋势,除1 100 m<海拔≤1 300 m和1 300 m<海拔≤1 500 m梯度内无显著差异外,其他梯度均差异显著。整体来看,海拔高度显著影响了乔木层物种多样性指标的变化,随海拔升高呈现逐渐降低的趋势。
在海拔<900 m梯度时,灌木具有最高的物种数量,随后降低,从海拔≥900 m开始,随海拔梯度的升高,物种数量呈先增大后显著降低趋势;同样,Shannon-winner多样性指数和Simpson多样性指数与之有相似的规律,相比而言,Shannnon-winner指数对灌木多样性指数随海拔变化趋势的敏感性更强;灌木Pielou均匀度指数除海拔>1 500 m梯度显著低于其他梯度外,其他海拔梯度无显著差异。
