杉木涩籽形成中类黄酮类代谢物的积累特征
2022-08-08武忆寒林思祖
吴 超, 熊 升, 武忆寒, 林思祖, 陈 宇
(1.福建农林大学林学院,福建 福州 350002;2.福建农林大学计算机与信息学院,福建 福州 350002;3.国家林草局杉木工程技术研究中心,福建 福州 350002)
杉木[Cunninghamialanceolata(Lamb.) Hook.]具有速生、高产、材质优良等特点,其人工林面积及蓄积量均位列我国用材树种首位[1]。杉木涩籽属于一类败育种子,其外观、大小、重量、色泽等表型特征与具备正常萌发能力的健籽极为相似,难以用简单方法区分[2]。已有报道认为,类黄酮类代谢物及其下游衍生物单宁类代谢物是涩籽的主要填充物质[3-8]。类黄酮类代谢物是2个具有酚羟基的苯环通过三碳键相连的一类代谢物的统称[9],广泛存在于植物各组织器官中,具有影响组织色泽[10-11]、抵抗病原体[12-13]、抗氧化[14-15]、参与信号传递[16-17]等功能。类黄酮类代谢物对植物种子发育的影响主要体现在协同调控种胚生长素水平[18-22]、影响胚乳脂质代谢[23-24]、调控microRNA表达[25-26]等方面,其异常积累可造成种胚败育[8]。
杉木涩籽中类黄酮类代谢物约占38.3%,是健籽的2倍[7],其种类超过140种,包括黄酮、黄烷酮、黄酮醇、异黄酮、黄酮碳糖苷、黄酮木脂素、花青素、儿茶素衍生物等多种类型[8]。目前,有关杉木涩籽发生过程中类黄酮类代谢物的合成、衍生及变化尚未明确。因此,本研究以涩籽率极高(90%)和极低(24%)的2个杉木家系种子为研究对象,比较种子发育过程中类黄酮类代谢物含量、酶活性及调控基因相对表达量的动态变化与差异,以探明涩籽形成过程中类黄酮类代谢物的积累特征,为进一步揭示杉木涩籽形成机制提供参考。
1 材料与方法
1.1 试验材料
供试样品取自福建省尤溪国有林场杉木3代种子园。根据本课题组前期对该种子园不同家系的种检数据,筛选出涩籽率分别为24%(L家系)和90%(H家系)的2个家系。2019年3月进行人工授粉,分别于授粉后70、80、90、100、110 d取样。各家系随机选取大小适中、无明显病虫害和机械损伤、位于树冠中部同一朝向的球果30枚。清洗球果表面灰尘后迅速置入液氮中速冻15 min,置于干冰中带回实验室。在干冰上迅速剥出种胚,置入浸于液氮中的无菌离心管,于-80 ℃超低温冰箱保存备用。
1.2 测定项目及方法
1.2.1 类黄酮类代谢物含量 (1)总黄酮与花青素含量采用试剂盒法测定。分别称取0.1 g种子样品,充分研磨后加入pH为7.0的磷酸盐缓冲溶液,1 000g离心20 min,取上清液。按照ELISA酶联免疫试剂盒说明书处理后测定450 nm处的光密度,以标准品绘制标准曲线并计算样品浓度。(2)槲皮素、山奈酚和杨梅素含量采用高效液相色谱法测定。称取0.5 g样品置于添加甲醇∶25%盐酸水溶液=4∶1(体积比)的研钵中,充分研磨成匀浆,经80 ℃水浴1.5 h后进行超声提取,10 000g离心10 min,取上清液,过滤后进样检测。液相色谱检测条件为:C18色谱柱(4.6 nm×250 mm,5 μm);流动相为甲醇∶0.2%磷酸=60∶40;流速0.8 mL·min-1,检测波长360 nm,柱温35 ℃。(3)儿茶素和表儿茶素采用高效液相色谱法测定。取0.5 g样品充分研磨后,按液料比为35∶1(mL∶g)溶于5 mL 60%乙醇溶液中,在50 ℃下回流提取3 h,所得溶液过滤后冷冻干燥。将干燥后的粉末复溶于5 mL 50%甲醇溶液中,过0.45 μm滤头后进样检测。液相色谱检测条件:A相为体积分数为0.05%的硫酸水溶液,B相为甲醇;流速为0.5 mL·min-1;柱温30 ℃。
1.2.2 酶活性 采用ELISA酶联免疫试剂盒法检测查尔酮合成酶、查尔酮异构酶、黄烷酮3′羟基化酶、二氢黄酮醇4-还原酶、黄酮醇合成酶、无色花青素还原酶、花青素合成酶和花青素还原酶等8种关键酶活性。按照试剂盒说明书,利用标准品绘制标准曲线并计算线性回归方程。利用1.2.1中总黄酮含量测定试验所制备的提取液测定452 nm处光密度,并计算酶活性。
1.2.3 调控基因相对表达量 以天根多糖多酚植物RNA提取试剂盒提取各样品RNA,测定查尔酮合成酶基因(CHS)、查尔酮异构酶基因(CHI)、黄烷酮3′羟基化酶基因(F3′H)、二氢黄酮醇4-还原酶基因(DFR)、黄酮醇合成酶基因(FLS)、无色花青素还原酶基因(LAR)、花青素还原酶基因(ANR)等7个关键调控基因的相对表达量。以全式金TransScript one-step gDNA Removal and cDNA Synthesis SuperMix反转录试剂盒制备样品cDNA,以primer 5.0软件设计各基因引物(表1),并根据引物序列的退火温度进行PCR扩增。以Trans PCR SuperMix试剂盒进行PCR产物回收。以全式金TA克隆连接试剂盒进行克隆连接,并将连接产物置于Trans1-T1感受态细胞中培养。挑选白色单克隆体进行PCR阳性克隆鉴定。以EasypureHiPureMaxiPrep Kit质粒提取试剂盒提取质粒,全式金2xTransTap High Fidelity PCR SuperMix试剂盒进行荧光定量测定。以EF-1α标准质粒为内参,双标准曲线法建立Rt-PCR的标准曲线。以授粉后70 d时H家系各基因单位细胞拷贝数为参照,做归一化处理得到各基因的相对表达水平。
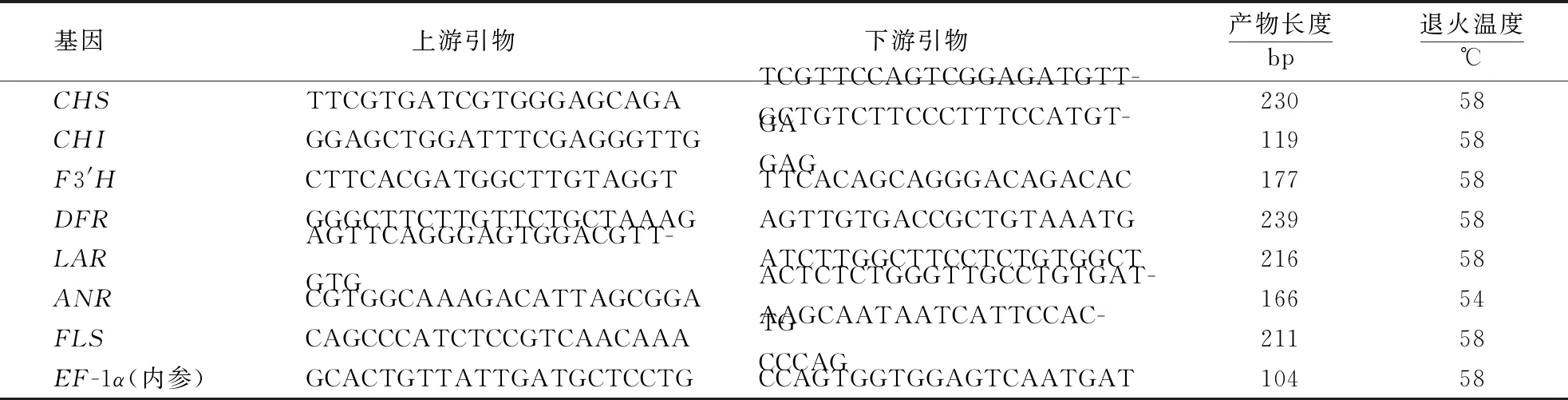
表1 基因引物设计
1.3 数据处理
以Origin V9.64软件对数据进行基本处理并绘图;以RStudio软件进行皮尔逊相关性分析;以TBtools的Heatmap功能绘制热图[27]。
2 结果与分析
2.1 杉木涩籽类黄酮合成途径代谢物含量比较
不同杉木家系种子类黄酮类代谢物含量及其变化趋势差异明显(图1)。由图1可知,授粉后70~110 d,L家系总黄酮含量变化不大;授粉后90~110 d,2个家系总黄酮含量差异逐渐扩大,至110 d时H家系总黄酮含量为L家系的2.17倍,且差异达极显著水平。花青素、槲皮素、山奈酚、杨梅素和儿茶素等中间代谢物含量整体上表现为H家系明显高于L家系。其中,H家系花青素含量随时间变化呈上升趋势,L家系则相反,授粉后70~80 d二者之间差异不显著,90~110 d则呈极显著差异,授粉后110 d差异最大,达6.45倍。H家系槲皮素和山奈酚含量均随时间变化呈先下降后上升的趋势,且授粉后70~90 d显著或极显著高于L家系。L家系杨梅素含量呈明显的下降趋势,授粉后70 d杨梅素含量高于H家系;随后H家系杨梅素含量快速上升,至授粉后110 d达到L家系的7.84倍,差异达极显著水平。H家系儿茶素含量呈明显的上升趋势,而L家系则先上升后下降,并在授粉后90 d达到最高值;授粉后70~90 d,2个家系儿茶素含量差异不显著;授粉后110 d,H家系儿茶素含量为L家系的2.44倍,差异极显著。表儿茶素含量表现与上述几种代谢物相反,授粉后80~110 d,H家系表儿茶素含量极显著低于L家系。
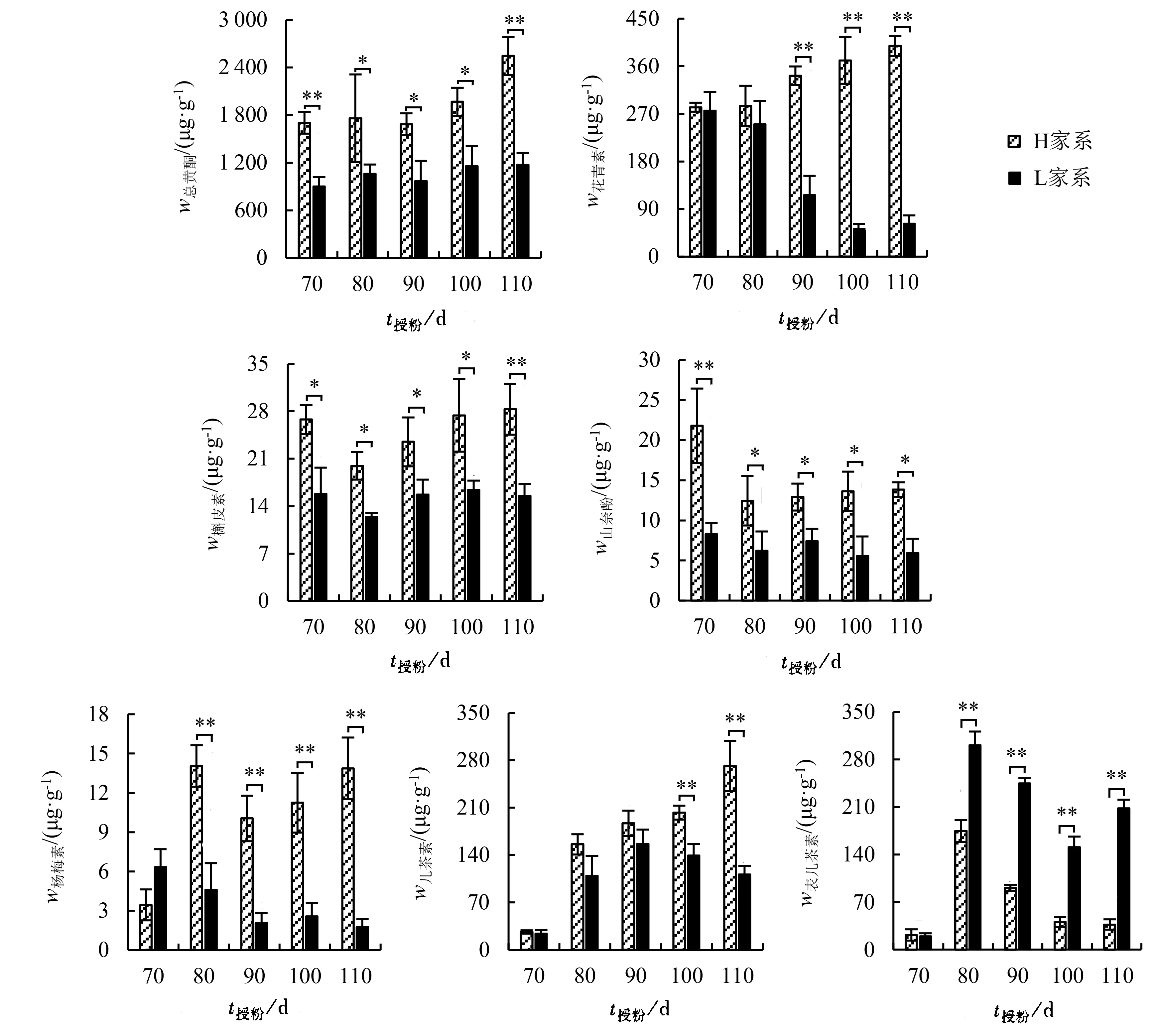
图1 不同家系杉木种子类黄酮合成途径代谢物含量比较
2.2 杉木涩籽类黄酮合成途径关键酶活性比较
2个家系间类黄酮合成途径关键酶活性存在差异(图2)。由图2可知,授粉后70 d,H家系查尔酮合成酶活性显著低于L家系,查尔酮异构酶和二氢黄酮醇4-还原酶活性则极显著低于L家系;授粉后80~110 d二者无显著差异。H家系黄烷酮3′羟基化酶、黄酮醇合成酶和无色花青素还原酶活性总体上高于L家系。其中,H家系黄烷酮3′羟基化酶活性随时间变化呈明显的上升趋势,而L家系仅在授粉后70~90 d有所上升,之后活性水平趋于平稳;授粉后70~80 d,2个家系黄烷酮3′羟基化酶活性水平差异不显著,90~110 d则呈现显著或极显著的差异水平。L家系黄酮醇合成酶和无色花青素还原酶活性均随时间变化而逐渐下降,授粉后100、110 d两种酶活性信号均已十分微弱,而H家系中这两种酶活性均较高;授粉后70~110 d,,2个家系间无色花青素还原酶活性差异均达极显著水平,黄酮醇合成酶活性除授粉后70 d两者无显著性差异外,其他时间两者差异均达极显著水平。H家系花青素合成酶活性随时间变化呈先上升后下降再上升的动态变化趋势;L家系则呈先上升后下降趋势,授粉后80 d活性最高;授粉后70 d,H家系花青素合成酶活性极显著低于L家系,授粉后100、110 d则极显著高于L家系。H家系花青素还原酶活性随时间变化呈逐渐升高趋势,L家系则呈先下降后上升再下降的动态变化趋势,授粉后70~90 d,H家系花青素还原酶活性极显著低于L家系,100、110 d则显著高于L家系。
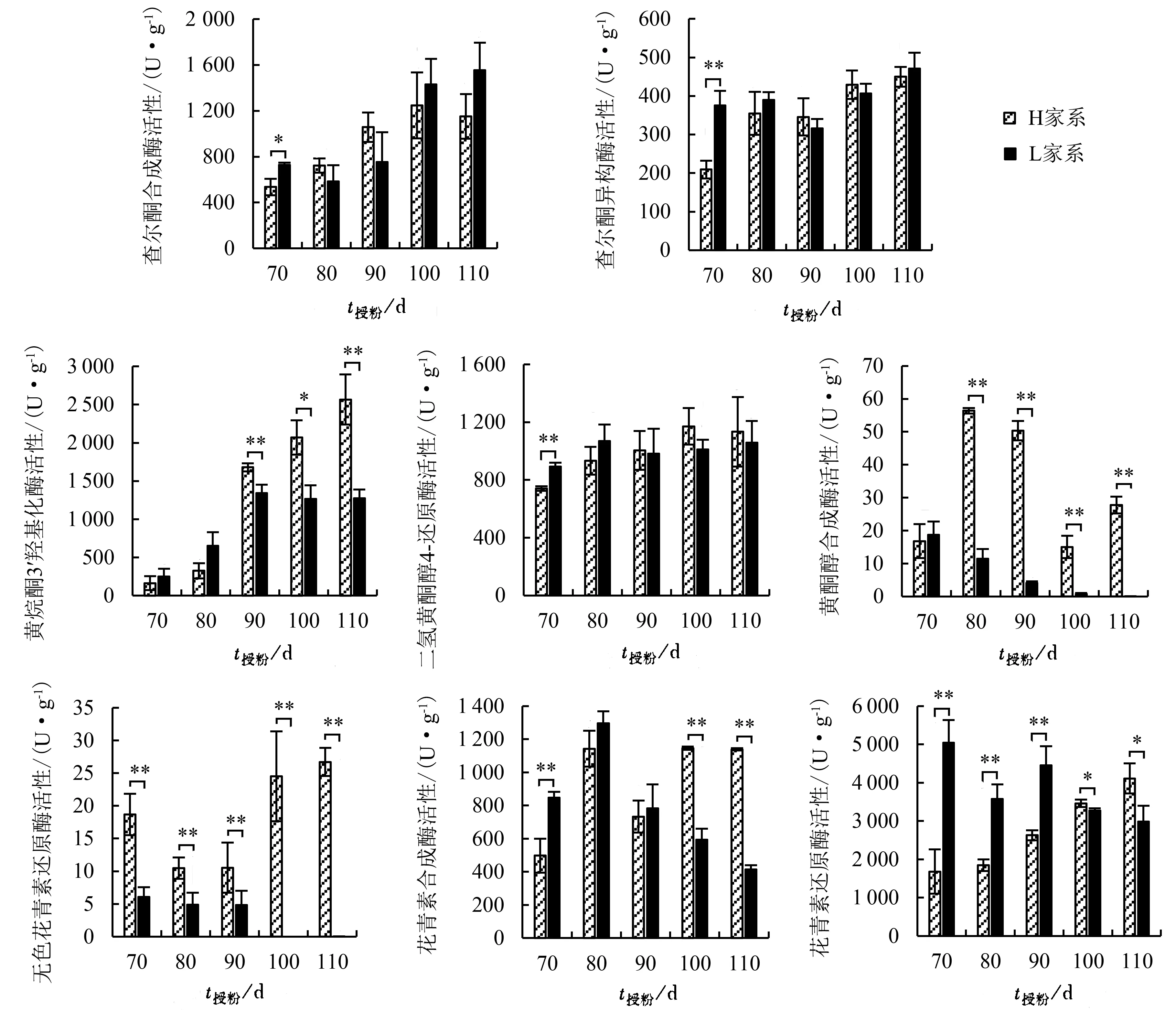
图2 不同家系杉木种子类黄酮合成途径关键酶活性比较
2.3 杉木涩籽类黄酮合成途径关键基因表达
2个家系类黄酮生物合成途径关键基因相对表达量见图3。2个家系种子的CHS、CHI相对表达量随时间变化呈先下降后上升趋势,且H家系均低于L家系。其中,两个家系的CHI相对表达量在授粉后90 d差异显著,其他处理差异均达极显著水平;CHS相对表达量在授粉后90~110 d差异极显著。H家系的F3′H相对表达量总体上随授粉天数变化而逐渐升高,而L家系则呈先上升后下降的趋势;授粉后70~90 d,H家系F3′H相对表达量极显著低于L家系,授粉后110 d则极显著高于L家系。2个家系DFR相对表达量均呈先上升后下降的趋势,授粉后90 d最大,除授粉后70 d二者无显著差异外,其他时间H家系均显著或极显著高于L家系。H家系FLS相对表达量随时间变化呈“W”型变化趋势,而在L家系中未检测到FLS基因的表达信号。L家系LAR与ANR的相对表达量均高于H家系,授粉后70~80 d两个家系的LAR相对表达量变化呈相反的趋势,80~110 d则均呈上升趋势,并在授粉后110 d差异达显著水平;授粉后70~110 d,H家系ANR相对表达量均极显著低于L家系。
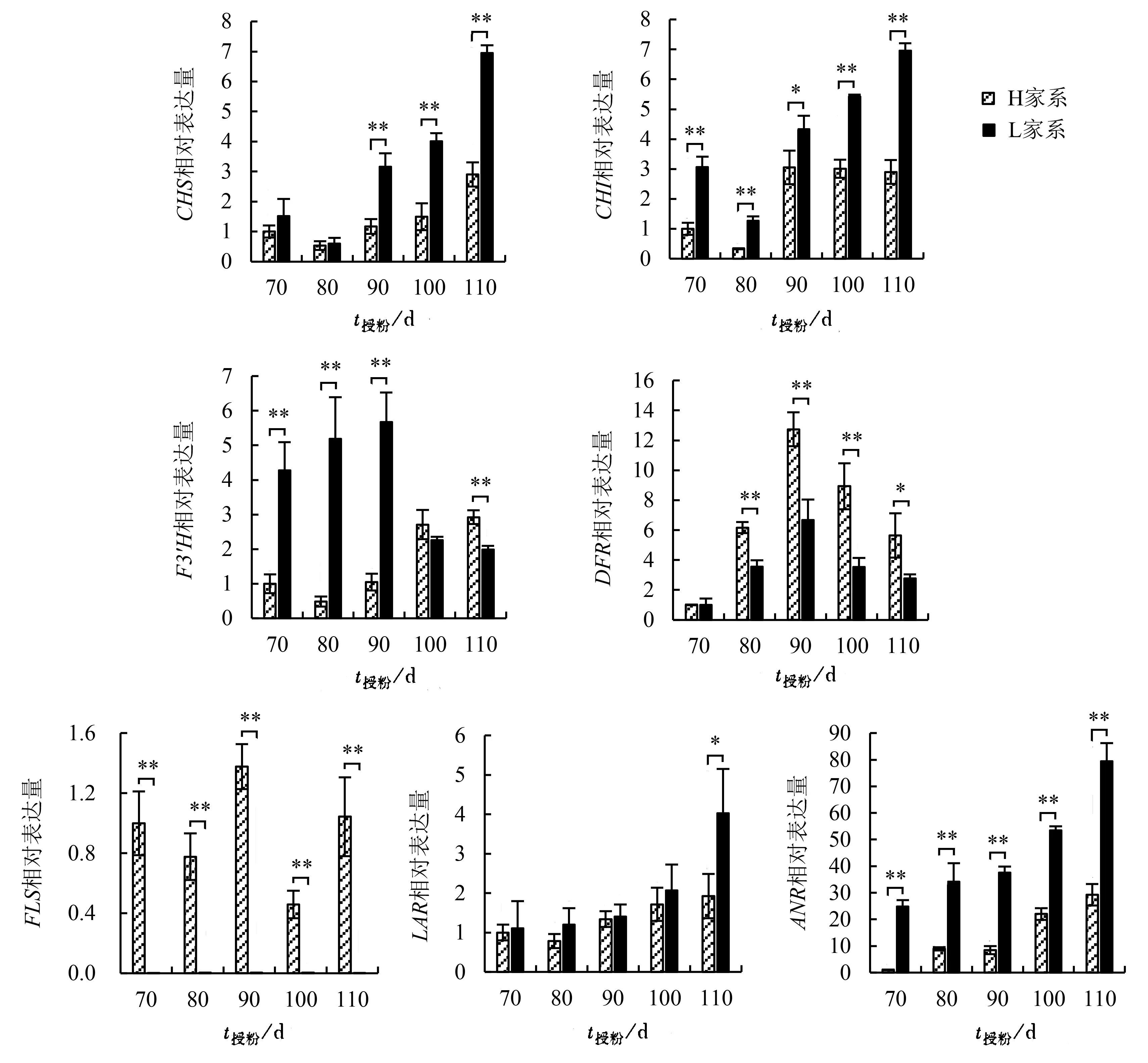
图3 不同家系杉木种子类黄酮合成途径关键基因表达比较
2.4 类黄酮类代谢物含量与酶活性及调控基因相对表达的相关性
杉木涩籽类黄酮类代谢物含量与关键酶活性、调控基因相对表达的相关性分析如表2所示。综合相关性系数和显著性指标,两个家系中与主要代谢物含量变化显著相关的酶活性及基因表达有所不同。L家系的总黄酮含量与查尔酮异构酶活性及CHS、LAR和ANR的表达呈显著正相关;H家系则与查尔酮异构酶、二氢黄酮醇4-还原酶、花青素合成酶活性及ANR的相对表达量呈显著正相关。L家系的花青素含量与查尔酮合成酶活性及CHS、CHI和ANR相对表达量呈显著正相关,与黄酮醇合成酶、无色花青素还原酶活性呈显著负相关;H家系则与查尔酮合成酶、黄烷酮3′羟基化酶、二氢黄酮醇4-还原酶、花青素还原酶活性及CHI和LAR相对表达量呈显著负相关。L家系的槲皮素含量与花青素合成酶活性呈显著负相关;H家系则与酶活性和基因相对表达量的相关性均不显著。L家系山奈酚含量与酶活性和基因表达的相关性均不显著;H家系则与查尔酮异构酶、花青素合成酶活性呈显著负相关。L家系杨梅素含量与二氢黄酮醇4-还原酶活性呈显著正相关,与花青素还原酶活性呈显著负相关;H家系则与查尔酮合成酶、黄烷酮3′羟基化酶、二氢黄酮醇4-还原酶活性呈显著负相关。L家系儿茶素含量与黄烷酮3′羟基化酶活性呈显著正相关,与黄酮醇合成酶活性呈显著负相关;H家系则与DFR的相对表达量呈显著正相关。两个家系表儿茶素含量与酶活性和基因表达的相关性均不显著。
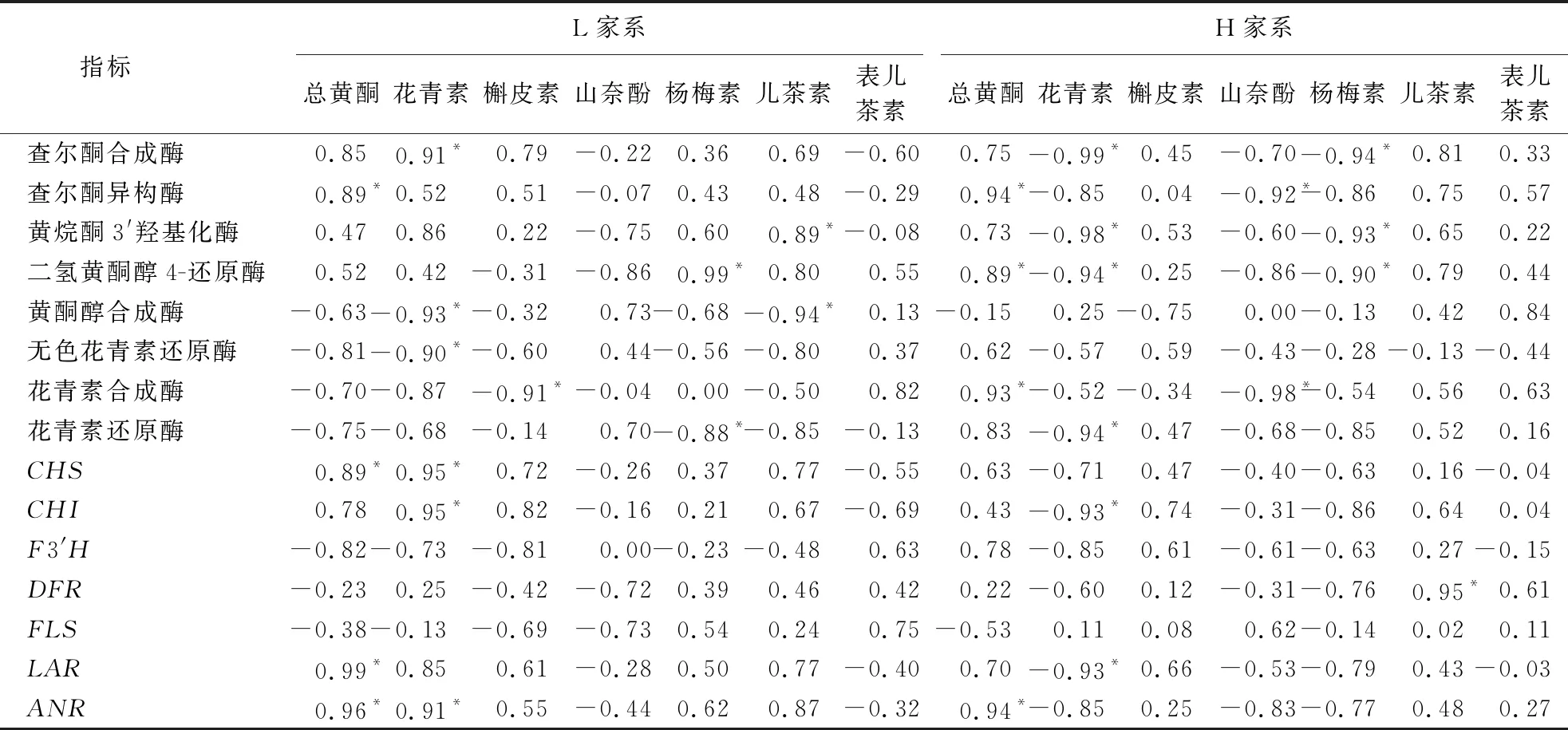
表2 类黄酮类代谢物含量与酶活性及调控基因相对表达的相关性分析1)
2.5 类黄酮生物合成途径差异性分析
通过比对可以看出(图4),上游的查尔酮合成酶和异构酶活性在两个家系间的差异不大,其编码基因CHS和CHI则在H家系中呈相对下调表达。中游的3个关键酶及其调控基因中,黄酮醇合成酶及其编码基因FLS在两个家系中的差异最为明显,在H家系中均为显著上调表达,其产物槲皮素、山奈酚和杨梅素含量也呈明显的上调趋势。下游的3个关键酶及其调控基因中,无色花青素还原酶活性有明显上调,其调控产物儿茶素含量也小幅上调,但编码基因LAR相对表达量的变化幅度不大。花青素还原酶编码基因ANR及其调控产物表儿茶素均有明显的下调表现,但其酶活性差异不大。
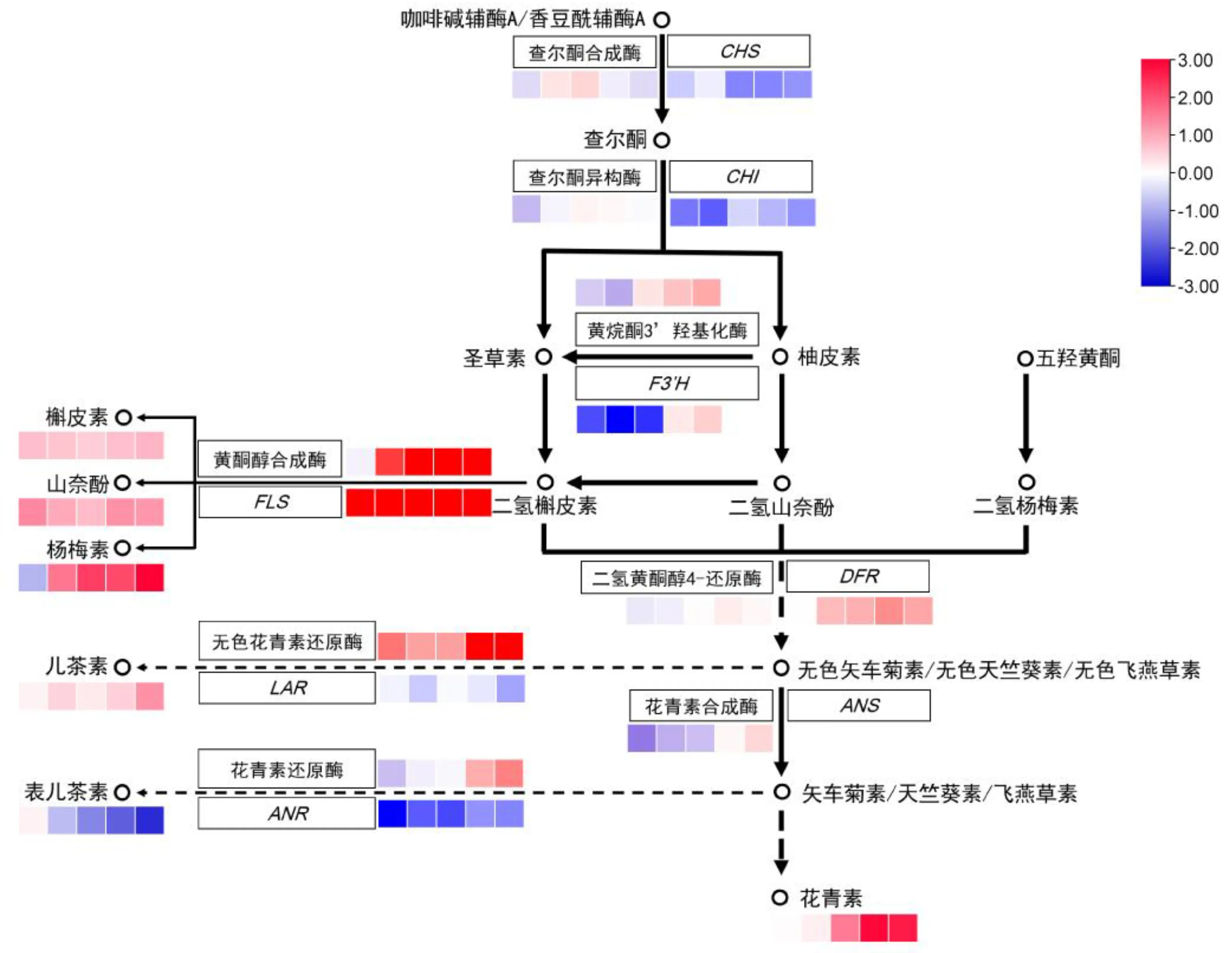
色块从左到右依次表示授粉后70~110 d H家系与L家系各代谢物含量比值的对数。
3 讨论
杉木涩籽形成过程中存在种胚萎缩败育和种腔被大量次生代谢物填充两个明显特征。早期研究认为,涩籽内的次生代谢物质主要为单宁类及其衍生物[3-7];而近期研究发现,在涩籽发生初期,包括黄酮、黄酮醇、黄酮炭糖苷、黄烷酮、异黄酮和花青素等类黄酮类代谢物及其衍生物的含量较高[8]。本研究表明,涩籽率较高的H家系种子发育过程中总黄酮和槲皮素、山奈酚、杨梅素、儿茶素、花青素等类黄酮生物合成途径中的重要代谢物含量明显高于L家系,表明涩籽发育初期类黄酮类次生代谢物质出现异常积累,这与林小琴[28]的研究结果相符。
查尔酮合成酶是类黄酮生物合成途径的第一个限速酶,其催化苯丙烷代谢途径的产物丙二酰辅酶A与香豆酰辅酶A或咖啡酰辅酶A缩合形成类黄酮生物合成途径的重要中间产物查尔酮[29]。在植物生殖发育系统中,查尔酮合成酶编码基因CHS被认为与雄性生殖成熟有关,通过干预CHS表达能够阻碍雄配子体的发育,进而影响胚胎形成[30]。杉木授粉期约在2月中旬至3月上旬,而受精期则在5月中旬[31]。授粉后小孢子将近2个月停留在大孢子珠孔附近[32],加之杉木球果种鳞为半开放状态,难以严密保护,使种胚发育易受外界环境的影响。H家系中CHS的下调表达反映了小孢子的异常发育可能与涩籽形成有关。
H家系中黄酮醇合成酶活性及其编码基因FLS的相对表达量均出现较大幅度的上调,同时槲皮素、山奈酚和杨梅素含量也明显增加,表明涩籽中二氢黄酮醇更趋向于向黄酮醇类代谢物方向衍化,尤其是杨梅素含量增加趋势最为明显,可见涩籽种胚内细胞的应激系统处于激活状态。相关研究表明,涩籽形成初期种胚上部开始逐渐萎缩,细胞破损解体[31],同时杨梅素已被证实参与多种植物逆境应激生理活动[33-34],这也印证了本文的推断。另外,黄酮醇类代谢物的其他下游产物,如儿茶素和花青素含量也有明显增加,这可能与底物含量的增加有关,也可能是由于涩籽内抗氧化酶系活性下降,相应的抗氧化类次生代谢物结合活性氧能力下降而导致活性氧积累所造成。
