外源精胺对碱胁迫下燕麦光合特性的影响
2022-08-08白健慧王育青
白健慧,王育青
(中国农业科学院草原研究所,内蒙古 呼和浩特 010010)
盐碱化土地面积的日益扩大,已成为全球的资源和生态问题,也是制约农业生产的重要因素[1]。内蒙古盐碱地面积较大,综合开发利用盐碱地已成为农学家主要的研究目标和热点[2-6]。种植抗性作物是开发利用盐碱地的重要途径之一,如种植碱蓬、田菁作物,可显著降低土壤盐碱度、提高土壤养分含量。据报道,在新疆塔里木盐碱化土地,碱蓬的移盐能力达2 116 kg/hm2[7];盐碱化土地种植苜蓿7年可显著降低土壤pH值[8]。有研究表明,种植抗盐禾本科作物,有利于在土壤中积累根系残体,降低0~40 cm土壤含盐量,脱盐率为67.3%[9]。燕麦属禾本科粮饲兼用作物,在内蒙古种植面积近13.3万hm2,约占全国的40%,居全国之首。燕麦具有抗盐碱、耐瘠薄等特点,有研究指出在高浓度盐胁迫下(200 mmol/L NaCl∶200 mmol/LNa2SO4=1∶1) 及高 pH 值环境下(pH值9.0)燕麦依然可生长[10]。因此,以燕麦为研究对象,进一步探讨燕麦耐盐碱生理机制具有生态价值[11],对改良盐碱化土地具有指导意义。
添加外源物质是改良作物抗性的重要手段,有助于抗性生理机制研究,并可为盐碱地作物栽培育种提供理论依据。多胺是有机、低分子量的含氮化合物,包括精胺、亚精胺、腐胺,多胺作为生长调节剂和胞内信使调节植物生长发育,参与植物抗逆生物过程[12]。同时多胺可作为抗氧化物质清除植物体内活性氧。有研究表明,外施多胺可提高作物非生物胁迫,如干旱、冷害、盐胁迫等抗性[13]。在盐胁迫盆栽试验中,外源多胺可提高番茄、小麦叶绿素含量,保护光合器官[14]。目前,精胺对作物抗碱性调控机制尚未进行深入研究。
植物产量的90%~95%来源于光合作用[15-16],对非生物胁迫环境下植物光合特性的研究十分必要。目前已有研究得出,在盐胁迫下,气孔因素导致燕麦光合速率降低[11];重度盐碱混合胁迫下,非气孔因素导致燕麦光合速率降低[17]。然而碱胁迫对燕麦光合系统的影响机制尚未进行深入研究。光系统是光合作用的重要位点,分为光系统Ⅱ和光系统Ⅰ,具有吸收、转化、传递光能的功能[18]。在非生物胁迫环境下,光系统Ⅱ发挥重要作用。叶绿素荧光曲线可用于研究光系统Ⅱ,协助探讨光系统Ⅱ反应中心光能吸收转化,过剩光能耗散,反应中心的供受体侧电子传递的变化[19]。目前有研究得出盐胁迫下,荧光曲线参数Vj降低,说明盐处理导致光系统Ⅱ反应中心出现不同程度关闭,导致单位面积内电子传递能量减少[19-21]。然而,该项技术尚未广泛应用于燕麦耐碱生理机制研究。本试验探讨了外施精胺对碱胁迫下燕麦光合特性的影响机制,旨在为盐碱地燕麦栽培育种提供参考。
1 材料和方法
1.1 试验材料
试验于2018年在农业农村部鄂尔多斯沙地草原生态环境试验站室内温室进行;所用材料为燕麦抗碱品种蒙燕1号和碱敏感品种太阳神,精胺购自Sigma公司,纯度大于99%。
1.2 试验设计
种植方式:采用盆栽试验,塑料盆规格280 mm×220 mm,种植基质为沙子,每盆装沙6.5 kg,播种35粒种子,间苗至20株,浇灌Hoagland营养液。
试验所用碱性盐成分为Na2CO3、NaHCO3,浓度设置为75 mmol/L。精胺选取2个处理浓度,分别为0.01、0.06 mmol/L,碱性盐与精胺直接溶解于Hoagland营养液。共设置4个处理(表1),重复4次。在燕麦两叶一心时进行碱及外施精胺处理,每次每盆加入溶液250 mL,碱胁迫后每隔2 d每盆加入1/2Hoagland营养液300 mL直到试验结束,于燕麦拔节、抽穗期进行数据测定。

表1 燕麦碱胁迫处理
1.3 测定指标与方法
光合参数测定:采用LI-6400(LI-CORBiosciences,Lincoln,NE,USA)光合仪,于燕麦拔节、抽穗期11:00时,测定倒数第二片叶光合速率(Pn)、气孔导度(Gs)、胞间 CO2浓度(Ci)、蒸腾速率(Tr),每个品种每个处理3次重复,取平均值。气孔限制值按照公式:Ls=1-(Ci/Ca)计算(Ci为胞间 CO2浓度,Ca为叶片周围CO2浓度)。
叶绿素含量:于燕麦拔节、抽穗期11:00时,使用便携式叶绿素仪测定倒数第二片叶SPAD值,每个品种每个处理3次重复,取平均值。叶绿素荧光参数:于燕麦拔节、抽穗期11:00时,运用Handy-PEA便携式植物效率分析仪直接导出OJIP曲线,测定燕麦倒数第二片叶初始叶绿素荧光(Fo)、光系统Ⅱ最大光化学效率(Fv/Fm)、单位面积吸收的光能(ABS/CSm)、单位面积捕获的用于还原QA的能量(TrO/CSm)、单位面积捕获的用于电子传递的能量(EtO/CSm)、性能指数(PIABS)。
1.4 数据处理分析
试验数据运用SPSS 17.0统计软件进行分析,组间数据进行单因素方差分析,运用Excel 2019软件绘制图表。
2 结果与分析
2.1 外施精胺对碱胁迫下燕麦叶绿素含量的影响
由图1可知,与CK相比,J处理拔节期蒙燕1号、太阳神的叶绿素含量分别降低37.50%、38.71%,抽穗期分别降低48.00%、53.06%,差异显著(P<0.05);蒙燕1号的叶绿素降幅低于太阳神,但差异不显著(P>0.05)。抽穗期两个燕麦品种的叶绿素降幅高于拔节期。与J处理相比,J+Spm1、J+Spm2处理可缓解碱胁迫对燕麦叶绿素含量的抑制,在拔节期蒙燕1号叶绿素含量分别提高25.00%、10.00%,太阳神分别提高15.78%、10.52%,差异显著(P<0.05);在抽穗期蒙燕1号叶绿素含量分别提高26.92%、15.38%,太阳神分别提高26.08%、13.04%,差异显著(P<0.05)。
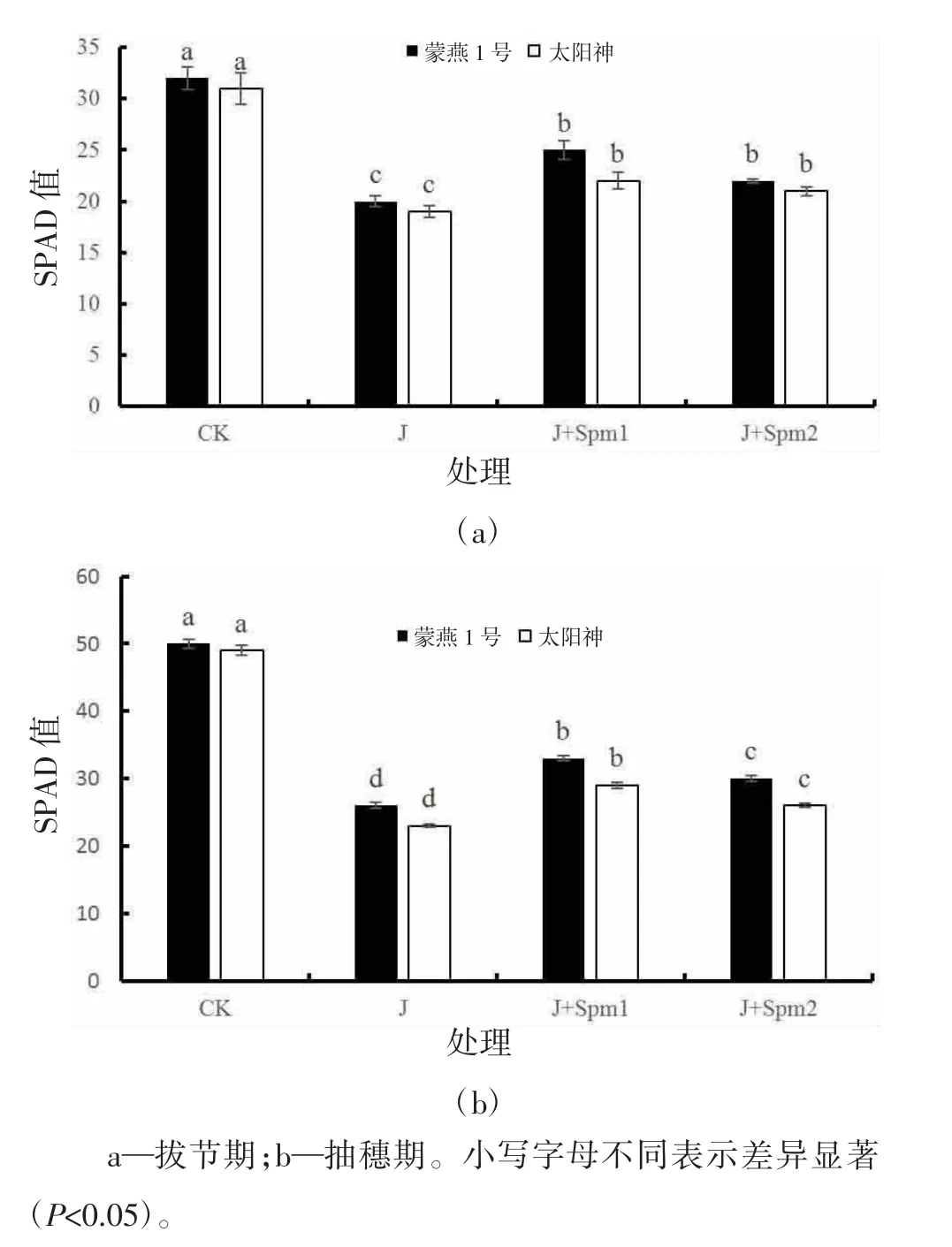
图1 外施精胺对碱胁迫下燕麦叶绿素含量的影响
2.2 外施精胺对碱胁迫下燕麦光合作用参数的影响
由表2可知,与CK相比,J处理两个燕麦品种光合速率显著降低(P<0.05),蒙燕1号、太阳神光合速率拔节期分别降低60.88%、58.93%,抽穗期分别降低71.07%、79.57%。从两个生育时期比较,J处理抽穗期的光合速率降幅高于拔节期,表明J处理对抽穗期光合速率的抑制作用高于拔节期;从两个品种比较,J处理蒙燕1号在抽穗期光合速率降幅低于太阳神。与J处理相比,J+Spm1处理在拔节期蒙燕1号、太阳神光合速率分别提高112.08%、61.64%,J+Spm2处理分别提高72.69%、27.06%;在抽穗期蒙燕1号、太阳神光合速率分别提高164.61%、200.42%,J+Spm2处理分别提高83.77%、160.90%。这表明外施精胺可缓解碱胁迫处理对燕麦光合速率的抑制,0.01 mmol/L精胺处理的缓解效果优于0.06 mmol/L;且在抽穗期外施精胺对太阳神碱胁迫光合速率的抑制作用缓解高于蒙燕1号。

表2 外施精胺对碱胁迫下燕麦光合特性的影响
与CK相比,J处理拔节期和抽穗期均显著降低气孔导度、蒸腾速率(P<0.05)。与J处理相比,不同浓度外施精胺显著提高两个品种的气孔导度、蒸腾速率(P<0.05)。J处理两个品种胞间CO2浓度显著低于CK(P<0.05),气孔限制值提高,说明气孔因素是光合速率降低的主要因素;J+Spm1、J+Spm2处理可促进胞间CO2浓度提高。
2.3 外施精胺对碱胁迫下燕麦叶绿素荧光参数的影响
由表3可知,与CK相比,J处理拔节期蒙燕1号、太阳神Fo分别提高9.78%、29.18%,抽穗期分别提高28.32%、32.39%;抽穗期Fo增幅高于拔节期,太阳神的Fo增幅高于蒙燕1号,Fo提高说明PSⅡ反应中心受到损伤。与J处理相比,J+Spm1、J+Spm2处理均降低蒙燕1号、太阳神Fo,J+Spm1处理下的Fo低于J+Spm2处理。
Fv/Fm是PSⅡ光化学反应状况的荧光参数,反映了植物的潜在最大光合能力。与CK相比,J处理Fv/Fm呈降低趋势;拔节期蒙燕1号、太阳神Fv/Fm分别降低3.90%、9.08%,抽穗期分别降低7.60%、10.56%。蒙燕1号的Fv/Fm降幅低于太阳神。与J处理相比,J+Spm1、J+Spm2处理对蒙燕1号、太阳神Fv/Fm差异不显著(P>0.05)。
ABS/CSm表示单位面积吸收的光能参数,与CK相比,J处理ABS/CSm呈降低趋势;拔节期蒙燕1号、太阳神分别降低9.70%、14.30%,抽穗期分别降低4.61%、15.65%。J+Spm1、J+Spm2对蒙燕1号、太阳神的ABS/CSm与J处理相比差异不显著(P>0.05)。
TrO/CSm表示单位面积捕获的光能。与CK相比,J处理TrO/CSm呈降低趋势;拔节期蒙燕1号、太阳神TrO/CSm分别降低13.25%、22.08%,抽穗期分别降低10.78%、24.57%。蒙燕1号TrO/CSm的降幅小于太阳神。与J处理相比,抽穗期J+Spm1处理下太阳神的TrO/CSm有所提高,且二者差异显著(P<0.05)。
EtO/CSm表示单位面积传递的光能。与CK相比,J处理EtO/CSm呈降低趋势;拔节期蒙燕1号、太阳神EtO/CSm分别降低25.55%、36.11%,抽穗期分别降低27.59%、42.37%。蒙燕1号EtO/CSm降幅低于太阳神。与J处理相比,拔节期J+Spm1、J+Spm2处理可提高太阳神EtO/CSm(P<0.05);抽穗期J+Spm1处理促进EtO/CSm提高,蒙燕1号、太阳神EtO/CSm分别提高6.13%、15.22%(表3)。
PIABS可准确表示植物光合机构整体状态,是反映胁迫处理对光合器官影响状态较为敏感的一个参数。由表3可知,J处理蒙燕1号、太阳神PIABS显著低于CK(P<0.05),拔节期蒙燕1号、太阳神PIABS分别降低56.58%、77.48%;抽穗期蒙燕1号、太阳神PIABS分别降低74.97%、79.51%。蒙燕1号的PIABS降幅低于太阳神;抽穗期蒙燕1号、太阳神PIABS降幅高于拔节期。与J处理相比,J+Spm1处理显著提高PIABS(P<0.05),拔节期蒙燕1号、太阳神PIABS分别提高24.45%、125.25%;抽穗期分别提高48.91%、23.96%;J+Spm2处理两个品种PIABS均低于J+Spm1处理。
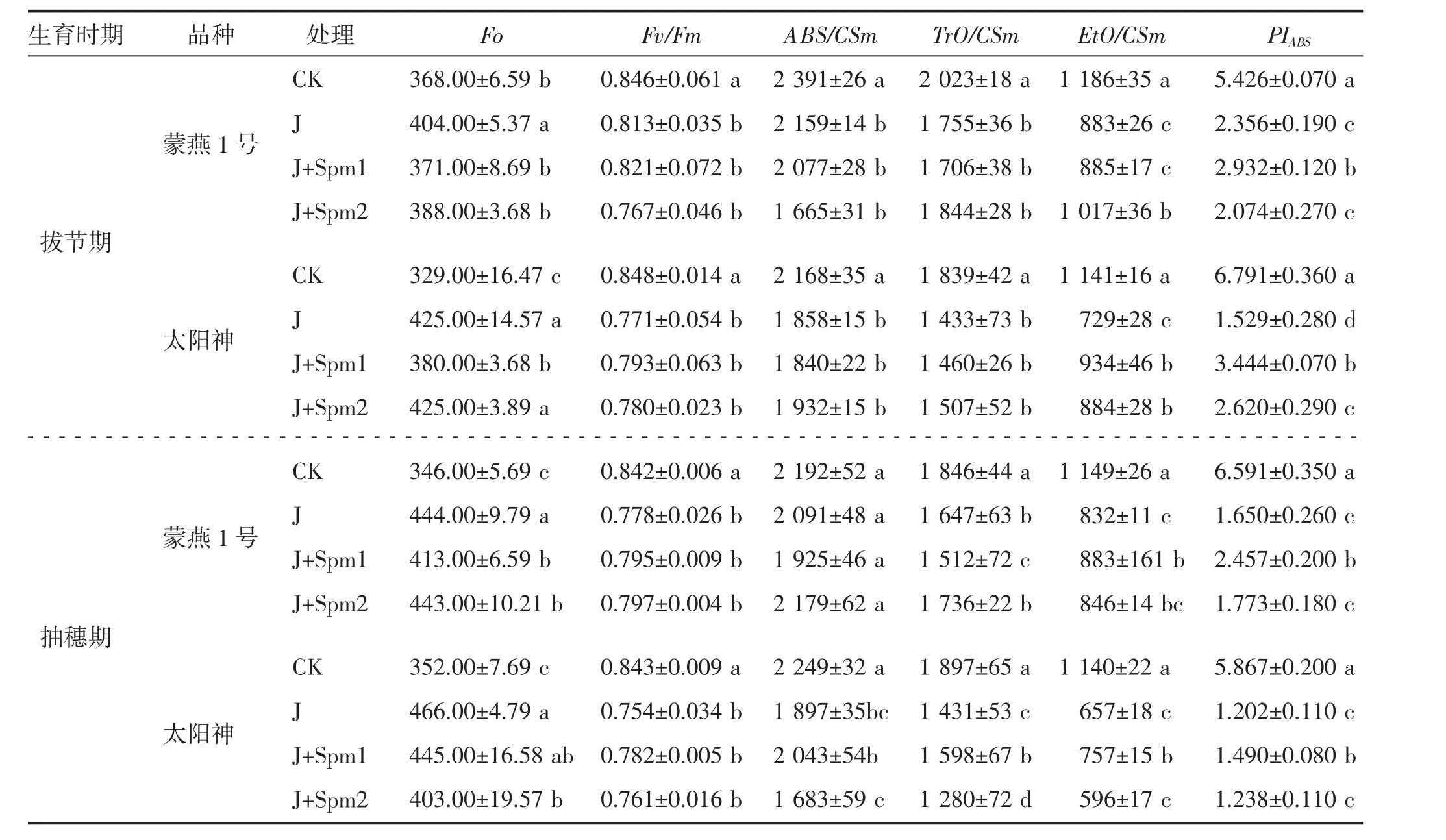
表3 外施精胺对碱胁迫下燕麦叶绿素荧光参数的影响
3 讨论
光合作用是产量形成的基础,是作物生产碳水化合物的生物过程[22-24]。前人研究多为盐胁迫、盐碱混合胁迫对植物光合作用的影响,对碱胁迫研究较少[25-26]。叶绿素是光合作用的重要色素,关系到作物生长发育及生产[27]。本试验结果表明,碱胁迫下,燕麦叶绿素含量呈降低趋势,这与李英丽等[28]碱胁迫导致燕麦叶绿素含量降低的研究结果一致。叶绿素含量降低会直接导致类囊体光合复合物组装错误,影响蛋白复合体功能,降低光合速率。本研究得出碱胁迫下,外施精胺可提高叶绿素含量,有助于光合作用进行。
气孔因素和非气孔因素是胁迫环境降低光合速率的主要原因。气孔因素是气孔开度降低导致胞间CO2浓度下降,用于光合作用原料减少,非气孔因素导致CO2溶解度下降、Rubisco酶活性降低、1,5-二磷酸核酮糖再生能力下降、光合器官关键结构稳定性降低[29-31]。本试验得出碱胁迫下,燕麦光合速率、胞间CO2浓度低于对照,气孔限制值提高,说明气孔限制因素是光合速率降低的主要原因。这与刘建新等[32]提出的碱胁迫下非气孔因素导致燕麦光合速率降低的研究结果不一致,这可能与品种差异有关。碱胁迫对抽穗期光合特性的抑制作用高于拔节期,这可能与胁迫时间较长有关。
本试验发现,在碱胁迫环境下,外施精胺对光合速率、气孔导度、蒸腾速率有一定促进作用。与碱胁迫相比,0.01 mmol/L精胺处理显著提高气孔导度说明外源精胺可促进CO2向叶绿体的运输,缓解胁迫环境导致的光合作用原料供应不足。与碱胁迫相比,0.01 mmol/L精胺处理可显著提高两个燕麦品种的蒸腾速率,说明精胺可通过增加蒸腾拉力,促进矿物质营养与水分的吸收与运输,从而有助于燕麦适应胁迫环境。与碱胁迫相比,0.01、0.06 mmol/L精胺处理可显著降低两个燕麦品种的气孔限制值,说明外施精胺能缓解胁迫环境对气孔造成的限制,促进进入气孔的CO2浓度增加,提高碳同化能力[33]。因此,外源精胺促进碱胁迫下两个燕麦品种的光合速率提高,这可能与蒸腾速率、气孔导度升高,气孔限制值降低有关。
为了进一步阐明外施精胺对碱胁迫下燕麦光能捕获、传递能力的影响,本试验检测了叶绿素荧光参数。叶绿素荧光参数常用来分析PSⅡ的性能变化,PSⅡ对逆境胁迫非常敏感,是衡量逆境胁迫损伤光合器官的有效指标[34]。Fo是初始荧光量,为PSⅡ反应中心处于完全开放时的荧光产量,Fo升高说明色素吸收能量用于光化学反应的部分减少[35],PSⅡ反应中心破坏或可逆失活会导致Fo升高,非光化学能量耗散、光合色素破坏会导致Fo升高[36]。本试验中,碱胁迫下,Fo高于对照,抽穗期两个燕麦品种的Fo高于拔节期,说明随着胁迫时间的增加PSⅡ反应中心开始失活。外施精胺较碱胁迫有利于降低Fo,说明精胺可协助保持PSⅡ反应中心稳定。Fv/Fm为PSⅡ最大光能转化效率,光抑制导致Fv/Fm降低,Fv/Fm已被广泛用作光抑制的诊断指标[36]。在碱胁迫下,两个燕麦品种的Fv/Fm低于对照。说明碱处理导致燕麦产生光抑制,这与番茄、高粱在盐碱处理下Fv/Fm降低的研究结果一致[37-38]。精胺处理对两个燕麦品种的Fv/Fm无显著影响。
PIABS是以吸收光能为基础的光合性能指数,可准确反映植物光合机构的整体状态,是反映光合系统受到环境影响的最敏感参数[38],碱胁迫处理下,两个燕麦品种的PIABS显著低于对照说明碱处理损伤光合机构。这与碱胁迫下高粱PIABS降低的研究结果一致[39]。碱胁迫下,两个燕麦品种的参数ABS/CSm、TrO/CSm、EtO/CSm呈降低趋势,说明在胁迫条件下,PSⅡ反应中心捕获传递光能的能力下降,反应中心失活,电子传递受阻[38-39]。本试验得出在抽穗期,与碱胁迫相比,0.01 mmol/L外施精胺可促进两个燕麦品种的TrO/CSm、EtO/CSm、PIABS值显著升高,说明精胺有利于燕麦增加单位面积用来还原QA的激发能和用于电子传递的能量,缓解PSⅡ反应中心失活,抑制光合机构损伤,改良光系统Ⅱ的整体性能。
4 结论
75 mmol/L碱胁迫显著降低燕麦拔节、抽穗期叶绿素含量、光合速率、气孔导度、蒸腾速率,提高气孔限制值,抽穗期光合速率降幅高于拔节期。75 mmol/L碱胁迫显著提高燕麦拔节、抽穗期Fo,降低Fv/Fm、TrO/CSm、EtO/CSm、PIABS值。外施精胺可促进碱胁迫下燕麦叶绿素含量、气孔导度、蒸腾速率提高,进而提高光合速率。抽穗期,0.01 mmol/L精胺较碱胁迫燕麦增加单位面积用来还原 QA的激发能(TrO/CSm)和用于电子传递的能量(EtO/CSm),进而提高PIABS值,改良碱胁迫下燕麦光系统Ⅱ整体性能。
