牦牛RBM3的基因克隆及其在不同繁殖时期卵巢、输卵管、子宫中的表达定位
2022-08-05潘阳阳王靖雷黄嘉馨余四九
张 晖,潘阳阳,王靖雷,张 瑞,黄嘉馨,余四九,崔 燕
(甘肃农业大学动物医学院,兰州 730070)
RNA结合基序蛋白3(RNA-binding motif protein 3, RBM3)作为一种已知的冷休克蛋白,在正常条件下存在于多种组织和细胞中,当动物机体与细胞在受到低温、低氧和局部缺血应激时可以刺激它的表达,并通过促进其他相关蛋白的合成维护应激条件下机体与细胞的正常生理功能。从人类组织分离得到的RBM3分子量为17 ku,最长的开放阅读框编码157个氨基酸。从结构上来看,RBM3包含两个结构域:一个是氨基酸末端的RNA识别基序(RRM),它包含两个高度保守序列:核糖核蛋白1(ribonucleoprotein1, RNP1)和核糖核蛋白2(ribonucleoprotein2, RNP2);另一个是富含甘氨酸、精氨酸和酪氨酸的羧基末端序列。RBM3作为一种富含甘氨酸的蛋白质,通过加速核糖体组装、稳定mRNA和减少microRNA的表达来促进蛋白质的翻译。Wellmann等在人类成纤维细胞内的研究发现,RBM3的mRNA和蛋白质在缺氧条件下的表达量有所增加。李云龙等也提出,在哺乳动物体内RBM3蛋白的表达量会随着温度的降低而不断增加。
研究证实,RBM3可以在包括人体细胞k562、HepG2、T24在内的多种细胞系中表达。而有关小鼠()的研究表明,RBM3可通过减少microRNA表达来诱导小鼠神经母细胞瘤细胞(N2a)中的整体翻译水平。Yan等研究发现,RBM3促进了在缺氧条件下神经干细胞(neural stem cell, NSC)的增殖。另外,RBM3在黑熊()这类冬眠动物的肌肉、肝和心中都有表达,并且在松鼠()冬眠晚期的心、肝、脑组织中高表达。2020年,潘阳阳等通过试验验证了RBM3参与牦牛()这一高原特有哺乳动物卵丘细胞低温应激调控。因此,研究在高寒低氧环境中哺乳动物经受低温时RBM3的表达变化,有利于RBM3参与调控动物机体缺氧、低温的机制研究。而牦牛作为在我国青藏高原地区的特有动物,由于长期生活在高寒低氧的高原地区,生态学特征独特。受到温度、含氧量等因素以及生存环境的综合影响,牦牛的繁殖能力低下,在自然条件下一年一胎或两年一胎限制了高原畜牧业的发展。所以,研究低温相关蛋白RBM3对牦牛生理调控机制具有重要作用。
本研究采集不同繁殖时期(卵泡期、黄体期和妊娠期)雌性牦牛的卵巢、输卵管、子宫,进行牦牛3基因克隆和相关生物信息学分析,并利用实时荧光定量PCR(quantitative real-time PCR, qRT-PCR)检测3 mRNA的表达量,利用蛋白质免疫印迹(Western blot, WB)检测RBM3蛋白的表达量,利用免疫组织化学法(immunohistochemistry, IHC)检测RBM3蛋白的表达定位,为探讨RBM3在高原环境下对牦牛生殖的调控提供理论依据。
1 材料与方法
1.1 样品采集
于2019年10月在青海省马佳肴屠宰场采集样品。卵泡期、黄体期和妊娠期的4~6岁健康雌性牦牛各3头。采集卵巢上见成熟卵泡(卵泡期)和新鲜黄体(黄体期)同侧以及妊娠3个月(妊娠期)妊娠侧的卵巢、子宫和输卵管。将所采样品修剪后,用生理盐水冲洗干净,一部分储存于-80 ℃,另一部分室温下保存于4%多聚甲醛溶液中。
1.2 主要仪器与试剂
普通PCR仪购于德国Eppendorf公司,qRT-PCR仪购于瑞士Roche公司,显微拍照仪购于日本Olympus公司,多功能成像仪购于美国GE公司;BeyoEcL PLus液购于上海碧云天公司,Trizol试剂盒购于北京全式金生物技术有限公司,TB GreenPremix Ex TaqⅡ、JM109感受态细胞购于大连TaKaRa公司;荧光定量PCR所用试剂、反转录试剂盒均购于美国Promega公司;DNA纯化回收试剂盒购于北京天根生化科技有限公司;基因克隆试验配制培养基所用试剂购于北京Solarbio公司;IHC试剂盒、WB所用抗体Rabbit Anti-beta-Actin antibody、Rabbit Anti-RBM3 antibody、Goat Anti-Rabbit IgG HH&L/RP以及DAB显色液购于北京Bioss公司。其他试剂均为国产分析纯。
1.3 牦牛RBM3基因克隆
1.3.1 引物设计 参照GenBank公布的牛()3基因序列(XM_024987852.1)进行引物设计,肌动蛋白(-)基因为内参,由上海生工进行引物合成,引物信息见表1。

表1 目的基因和内参基因引物序列Table 1 Primer sequences amplifying target and house-keeping genes
1.3.2 总RNA提取 试剂盒法提取样品总RNA,测定所提取的RNA浓度和OD/OD值后,将达到浓度和OD/OD值参照标准的RNA进行反转录,合成的cDNA保存在-20 ℃备用。
1.3.3 牦牛3基因克隆 以样品cDNA为模板扩增3基因。总反应体系为20 μL:TaqPCR Master Mix 10 μL, ddHO 8 μL, cDNA模板1 μL, 10 μmol·L的上、下游引物各0.5 μL;反应条件:95 ℃预变性3 min;95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸10 s,72 ℃保存5 min,循环35次进行PCR反应。将PCR产物进行凝胶电泳,回收产物于-20 ℃保存。用TaKaRa试剂盒构建重组质粒,将回收产物与pMD19-T Vector连接,然后把连接产物(10 μL)转化至 JM109感受态细胞(100 μL)。 把连接转化后的产物加入890 μL SOC液体培养基中,摇床培养(37 ℃, 200 r·min)15 h, 涂布于LB琼脂平板培养基(含有Amp, X-gal, IPTG)上进行培养(37 ℃, 14 h),挑取平板培养基上的单个白色菌落并将其置于含有Amp的LB液体培养基,37 ℃振荡培养16 h。最后,将得到的菌液进行测序,根据测序结果对3基因进行后续分析。
1.3.4 生物信息学分析 对牦牛3基因的测序结果进行分析,具体分析网站信息见表2。

表2 生物信息学分析网站Table 2 Bioinformatics analysis websites
1.4 qRT-PCR检测RBM3基因的表达
qRT-PCR检测牦牛不同繁殖阶段卵巢、输卵管、子宫中3基因的表达水平。在20 μL (TB GreenPremix Ex TaqⅡ 10 μL,Free Water 6.4 μL, cDNA模板2 μL,10 μmol·L的上、下游引物各0.8 μL)的体系下以95 ℃预变性30 s,95 ℃变性5 s、 60 ℃退火30 s、72 ℃延伸10 s,循环40次的条件进行PCR反应,重复4次。按照2计算3基因在每组样品中的相对表达量。
1.5 蛋白免疫印迹法检测RBM3蛋白的表达
1.5.1 蛋白样品的制备 将试验样品低温研磨后取110 mg,按照蛋白裂解液(Radio Immunoprecipitation assa, RIPA)与蛋白酶抑制剂(phenylmethanesulfonyl fluoride, PMSF)100∶1的比例配制蛋白裂解液并取110 μL加入研磨好的试验样品。然后4 ℃振荡裂解2 h后,4 ℃离心(12 000×g)5 min, 吸取上清在-80 ℃保存备用。
1.5.2 蛋白质免疫印迹 经十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate polyacrylamide gel electrophoresis, SDS-PAGE),电泳后电转膜(湿转)至聚偏二氟乙烯膜(polyvinylidene fluoride, PVDF)上,5%脱脂牛奶室温振荡封闭2 h。 将目的条带和内参条带分别加一抗兔抗RBM3抗体(1∶1 000 PBST稀释)和兔抗beta-Actin抗体(1∶2 000 PBST稀释)4 ℃孵育11 h后用PBST(PBS∶Tween20=2 000∶1) 溶液洗10次(6 min·次); 山羊抗兔IgG H&L/HRP(1∶3 000 PBST稀释)二抗室温摇床孵育1 h后PBST溶液洗6次, 每次15 min;在膜上滴加电化学发光液避光孵育2 min后WB扫描成像仪扫描蛋白条带,重复3次, 最后分析RBM3蛋白相对表达量。
1.6 IHC定位RBM3蛋白表达
固定组织,依次脱水、包埋、切片(厚度4 μm)、脱蜡、热修复。再进行如下步骤:滴加3% HO并在湿盒孵育10 min(37 ℃);滴加封闭液在湿盒孵育15 min(37 ℃);滴加一抗兔抗RBM3抗体(1∶400 PBS稀释)在湿盒孵育过夜(4 ℃),同时设置只加PBS(0.02 mol·L)的对照;滴加二抗(B液)在湿盒孵育15 min(37 ℃);滴加C液在湿盒孵育15 min (37 ℃);显色。最后苏木精复染、脱水、透明、树脂封片并拍照。
1.7 数据分析
依据qRT-PCR结果计算3的相对表达量,利用Image J软件灰度值分析RBM3蛋白的表达量(目的灰度数值/内参灰度数值),运用SPSS软件对基因和蛋白相对表达量的差异显著性进行单因素方差分析,用软件GraphPad Prism 8绘制数据图。
2 结 果
2.1 牦牛RBM3基因的扩增与克隆
PCR结果显示(图1),9组样品在618 bp处出现预期的整齐条带。与NCBI数据库与参考序列将3基因克隆测序结果进行比对后,相似性100%。已将牦牛3序列提交至GenBank(登录号:MF142258.1)。本部分试验检测出了牦牛3序列,此结果将继续用于后续生物信息学分析等试验内容。
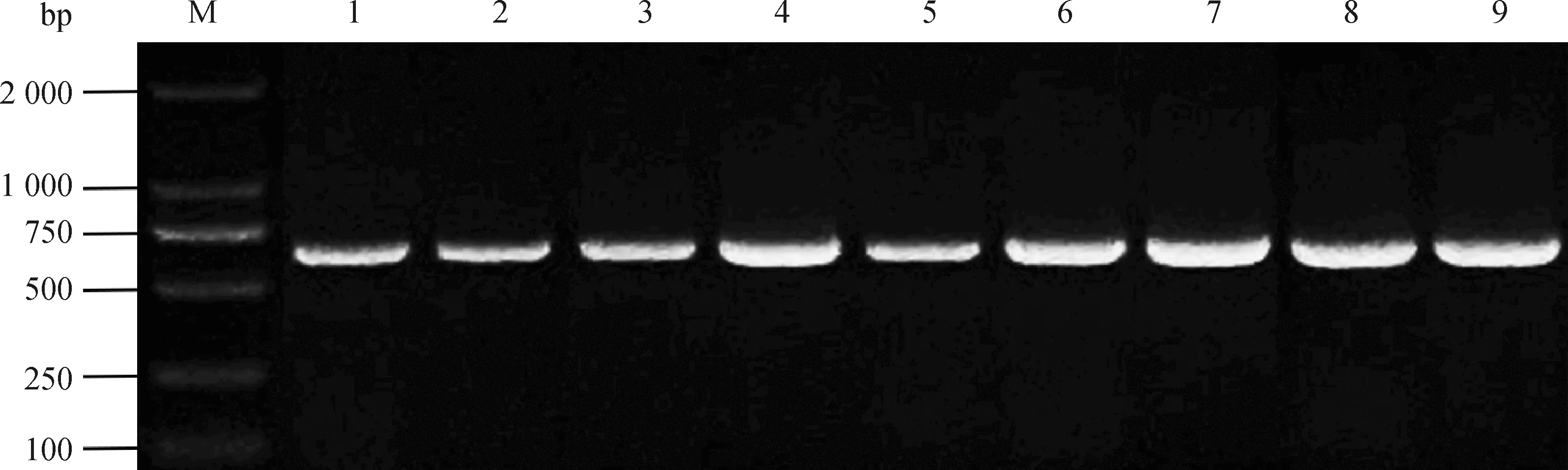
M.DNA相对分子质量标准;1~3.卵泡期、黄体期、妊娠期卵巢;4~6.卵泡期、黄体期、妊娠期输卵管;7~9.卵泡期、黄体期、妊娠期子宫M.DL2000 DNA Marker; 1-3. The ovary at follicular, luteal, gestation phases; 4-6. The fallopian tube at follicular, luteal, gestation phases; 7-9. The uterus at follicular, luteal, gestation phases图1 RBM3 PCR扩增电泳结果Fig.1 The electrophoresis results of RBM3 PCR amplification
2.2 生物信息学分析
2.2.1 牦牛3基因开放阅读框分析 开放阅读框分析发现,牦牛3的开放阅读框(ORF)全长为483 bp,编码了160个氨基酸。选取具有代表性的动物进行相似性比对,结果显示牦牛3基因与野牦牛()相似性最高(99.79%),相似性较高的还有数值为99.59%的瘤牛()与99.17%的普通牛();与其他动物的相似性从高到低依次为:山羊()98.96%、弯角剑羚()98.76%、水牛()97.17%、绵羊()96.16%、单峰驼()91.36%、羊驼()91.36%、野骆驼()91.36%、西伯利亚虎()91.30%。经MEGA 7.0软件比对(图2)后,构建系统进化树结果显示(图3),牦牛3基因与野牦牛亲缘关系最近。将牦牛与包括基因相似性最高的野牦牛在内的所有在列动物相比,3基因编码区序列在第98位核苷酸(图2黑色箭头处)有差异,且对应编码的第33位氨基酸在牦牛上是苯丙氨酸(Phe),而在其他动物上编码的结果是丝氨酸(Ser)。另外,第51位核苷酸的比对中,牦牛、野牦牛和除西伯利亚虎之外的所有在列动物都有差异,但其编码的氨基酸均为苏氨酸(Thr),没有改变。

黑色箭头标记是第51、98位核苷酸Black arrows point to nucleotide No.51 and No.98图2 不同物种间RBM3基因序列比对Fig.2 Sequence alignment of RBM3 orthologues among different species

图3 RBM3基因的系统进化树Fig.3 Phylogenetic tree of RBM3 gene
2.2.2 牦牛3基因编码蛋白的理化性质分析和结构预测 RBM3蛋白预测分子量大小17.5 ku。分析后得知,3基因编码了含有18种氨基酸在内的160个氨基酸,甘氨酸(Gly)的含量最高,为21.2%;含量最低的是组氨酸(His)、异亮氨酸(Ile)、赖氨酸 (Lys)、和甲硫氨酸(Met),含量为1.9%;半胱氨酸(Cys)、色氨酸(Trp)的含量为0。3基因编码的氨基酸中含有带负电的氨基酸(Asp+Glu)20个(12.5%),带正电氨基酸(Arg+Lys)22个(13.8%)。该编码区共2 361个蛋白原子,理论等电点(PI)8.83,不稳定指数为24.58,是稳定蛋白。二级结构预测结果显示(图4A),构成RBM3蛋白α-螺旋的氨基酸个数为21个,占氨基酸总数的13.1%;构成RBM3蛋白延伸链的氨基酸有30个, 占18.8%;109个氨基酸无规则卷曲,占68.1%。RBM3蛋白的三级结构预测结果如图4B所示。
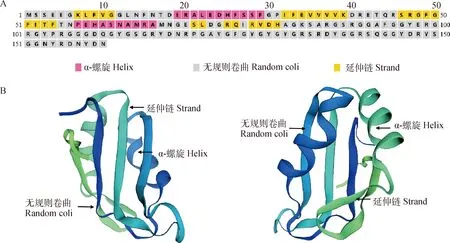
A. 牦牛RBM3蛋白的二级结构预测;B.牦牛RBM3蛋白三级结构预测的两个不同角度A. The secondary structure prediction of RBM3 protein in yak; B. Prediction of tertiary structure of yak RBM3 protein from two different angles图4 牦牛RBM3蛋白结构预测Fig.4 The structure prediction of RBM3 protein in yak
2.2.3 牦牛RBM3蛋白跨膜结构域和磷酸化位点分析 蛋白跨膜结构域预测结果(图5A)表明RBM3蛋白不是跨膜蛋白。蛋白磷酸化位点分析结果(图5B)表明,在组成RBM3蛋白的氨基酸中有10个丝氨酸(Ser)、2个苏氨酸(Thr)、5个酪氨酸(Tyr)的磷酸化位点具备成为蛋白质激酶磷酸化位点的条件。

A.牦牛RBM3蛋白跨膜区域分析;B.牦牛RBM3蛋白磷酸化位点分析A.The protein transmembrane regional analysis of RBM3 protein in yak; B. The protein phosphorylation site analysis of RBM3 protein in yak图5 牦牛RBM3蛋白跨膜区域和磷酸化位点分析Fig.5 The protein transmembrane region and phosphorylation sites analysis of RBM3 protein in yak
2.3 RBM3基因表达检测
qRT-PCR结果(图6)显示,3基因在不同繁殖阶段的牦牛卵巢、输卵管、子宫中都有表达。在卵巢中,3基因在卵泡期的表达量显著低于黄体期和妊娠期(<0.05);在输卵管中,3基因在卵泡期的表达量显著高于黄体期和妊娠期(<0.05);在子宫中,3基因在各时期表达量存在明显差异,卵泡期最低,黄体期和妊娠期较高(<0.05)。3基因在牦牛发情周期不同阶段的主要生殖器官上呈差异性表达。

β-actin为内参基因;n=4。不同字母代表差异显著(P<0.05)β-actin is the reference gene; n=4. Different letters indicate significanct difference (P<0.05)图6 RBM3 mRNA在牦牛不同组织中的相对表达量Fig.6 Relative expression of RBM3 mRNA in different tissues in yak
2.4 RBM3蛋白表达检测
WB结果(图7A)表明,RBM3蛋白在卵泡期、黄体期、妊娠期的牦牛卵巢、输卵管、子宫中都有表达且存在差异。通过定量分析(图7B)在卵巢上,RBM3蛋白在黄体期的表达量最高,妊娠期次之,卵泡期最低,3个样本之间差异显著(<0.05);在输卵管上,RBM3蛋白在黄体期和妊娠期表达较高且两者之间差异不显著(>0.05),卵泡期的表达量最低,和同组的其他两个样本之间的差异均显著(<0.05); 在子宫中,RBM3蛋白在妊娠期的表达量最低,卵泡期的表达量最高,黄体期的表达量介于卵泡期和妊娠期之间,3个样本间差异显著(<0.05)。
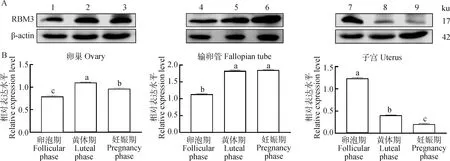
A.RBM3和β-actin蛋白在牦牛不同组织中的检测结果;B.RBM3蛋白在牦牛不同组织中的相对表达量。1~3. 卵泡期、黄体期、妊娠期卵巢;4~6. 卵泡期、黄体期、妊娠期输卵管;7~9. 卵泡期、黄体期、妊娠期子宫A. Detection results of RBM3 and β-actin in different tissues of yak; B. Relative expression of RBM3 in different tissues of yak. 1-3. The ovary at follicular, luteal, gestation phases; 4-6. The fallopian tube at follicular, luteal, gestation phases; 7-9. The uterus at follicular, luteal, gestation phases图7 RBM3蛋白在牦牛不同组织中的表达情况Fig.7 Expression of RBM3 protein in different tissues of yak
2.5 牦牛RBM3蛋白在主要生殖器官的定位检测结果
通过免疫组织化学方法检测(图8)发现,RBM3蛋白在牦牛发情不同阶段的卵巢、输卵管、子宫切片中都有阳性表达,且均为细胞质表达。在卵巢上(图8A、8B、8C)RBM3的主要表达分布在卵泡颗粒层(follicle granular layer, SG)、卵泡膜(theca follicle, TF)、黄体细胞(luteal cells, LC);RBM3在输卵管上(图8D、8E、8F)的主要表达位置为黏膜上皮(epithelium mucosae, EM);RBM3在子宫上(图8G、8H、8I)的主要表达位置为子宫内膜(endometrium, EN)和子宫腺(uterine glands, UG)。

A-I. 阴性表达;A1、A2、B1、B2、C1、C2. 卵巢;D1、D2、E1、E2、F1、F2. 输卵管;G1、G2、H1、H2、I1、I2. 子宫。SG. 卵泡颗粒层;TF. 卵泡膜;LC. 黄体细胞;EM. 黏膜上皮;EN. 子宫内膜;UG. 子宫腺。卵泡期. A组、D组、G组;黄体期. B组、E组、H组;妊娠期. C组、F组、I组A-I. Negative expression; A1, A2, B1, B2, C1, C2. Ovary; D1, D2, E1, E2, F1, F2. Fallopian tube; G1, G2, H1, H2, I1, I2. Uterus. SG. Follicle granular layer; TF. Theca follicle; LC. Luteal cells; EM. Epithelium mucosae; EN. Endometrium; UG. Uterine glands. Follicular phase. Group A, D and G; Luteal phase. Group B, E and H; Gestation phase. Group C, F and I图8 RBM3蛋白在在牦牛不同组织中的分布Fig.8 Distribution of RBM3 proteins in different tissues of yak
3 讨 论
RBM3作为动物体内的一种应激调节基因,其与冷应激的关系在1997被发现,但越来越多的研究表明它在细胞中有着较为复杂的功能和作用。在正常生理条件下,它可以保护细胞免受内在刺激造成的损伤,而在机体遭受低温、低氧等应激刺激时,则会诱导其表达并通过多种途径调节应激反应。有研究表明,当动物机体内的RBM3表达量增加时,机体对抗外界有害刺激的能力就会有所增加,从而在恶劣环境下为细胞提供一个较为良好的内部环境。近些年来,RBM3被认为是潜在的原癌基因进行了大量与之相关的研究,但对其参与生殖调控的研究却不多。本研究成功克隆了牦牛3基因,分析后发现其N端含有特殊蛋白结构域RNP1和RNP2,与前人研究结果一致,存在RGG结构域。有学者发现,RBM3与部分冷休克蛋白(cold shock proteins, CSPs)具有中度核苷酸序列和功能相似性,在进化过程中保守,本研究结果显示牦牛3与野牦牛、瘤牛、普通牛亲缘性较近,其在物种间高度保守,与前人研究RBM3是高度保守的RNA结合蛋白的结论一致。3的核苷酸序列与其他物种相比存在差异:第51位核苷酸和与除西伯利亚虎之外的所有比对动物都有差异,但编码的氨基酸没有发生改变(苏氨酸,Thr);第98位核苷酸也存在差异,所编码氨基酸由参与机体糖代谢、脂肪代谢的苯丙氨酸(Phe)变为在脂肪、脂肪酸新陈代谢和肌肉的生长中发挥重要作用的丝氨酸(Ser),可能为牦牛3基因特点,需进一步研究。
本试验分别对RBM3的mRNA和蛋白在牦牛子宫、卵巢和输卵管中相对表达量进行了检测,发现卵泡期、黄体期、妊娠期3个不同繁殖时期的RBM3在mRNA和蛋白层面的表达趋势存在差异。这可能是由于3基因在本试验涉及到的雌性牦牛不同时期、不同部位存在翻译和转录层面的调控,如磷酸化和甲基化,都对3生物学功能有重要的调节作用,由此出现时空表达差异。这一差异的存在为后续进一步深入开展3生理功能和翻译后修饰的相关研究提供了思路,有待进一步研究。
在雌性哺乳动物生殖过程中,卵巢作为维持雌性特征最重要的腺体器官,在生殖过程以及相关调控中发挥着重要作用。本研究发现,RBM3在牦牛卵巢中的主要表达位置为卵泡颗粒层、卵泡膜以及黄体细胞,并在黄体期和妊娠期呈现高表达。黄体作为在卵巢内形成的内分泌腺体样结构,通过产生孕酮、雌激素以及其他激素来调控个体发情周期和妊娠。卵泡颗粒细胞作为参与调控原始卵泡的启动、发育、卵泡选择、成熟以及排卵过程的重要角色,与卵泡膜细胞之间也存在着一定相互作用,而且这种相互作用在卵泡发育和卵母细胞的成熟过程中有重要意义。在卵巢中,卵泡破裂排卵后退化形成黄体,前人研究表明黄体细胞可以通过自分泌与旁分泌方式在黄体功能调节中起到局部调节作用,其中黄体期黄体通过分泌孕酮来抑制卵巢排卵、妊娠期黄体持续分泌孕酮维持妊娠。因此,RBM3在牦牛黄体期、妊娠期卵巢的高表达可能与黄体功能调节和妊娠维持有关系。
输卵管作为动物体内精子获能、受精及胚胎早期发育的场所。在本试验中,RBM3在牦牛输卵管中的主要表达位置为黏膜上皮,黏膜上皮可以通过分泌物影响精卵结合和合子的形成,对早期胚胎的发育和转运具有重要作用。从已知的研究结果来看,输卵管在卵泡期受卵巢分泌雌激素的影响,黏膜上皮细胞快速生长、发育和增殖,分泌输卵管液为受精和早期胚胎发育提供稳定的环境。卵巢排卵后,卵泡液和卵子一起进入输卵管,在黄体期输卵管中卵泡液和输卵管液的融合为精子到达受精部位提供了保障,妊娠期输卵管变粗、伸长、弯曲变多并促进早期胚胎的发育。本试验发现,RBM3在黄体期和妊娠期输卵管的表达量显著高于卵泡期,结合免疫组化结果推测RBM3在牦牛输卵管中参与了生殖配子在输卵管内的运行以及早期胚胎发育过程,这可能和RBM3调控细胞生长、分化的生理学功能有关。
子宫作为胎生动物胚胎附植以及生长发育的场所,其生长发育和活动由复杂的内分泌系统所控制,并且具有重要的生理功能。在本研究中,RBM3在子宫的主要表达位置为子宫内膜和子宫腺,而且在子宫卵泡期的表达显著高于黄体期和妊娠期。子宫内膜作为子宫的壁层具有促进发情周期循环、孕育胎儿、内分泌等作用;而子宫腺则可以分泌多种妊娠识别、胚胎着床、促进胎儿发育的多种物质。在卵泡期阶段,子宫内膜以增殖、生长以及细胞内代谢为主,子宫腺的分泌水平也在这一时期较高。子宫内膜可以通过腺体产生的大量因子,而子宫腺通过感受激素刺激可以分泌相关因子,还可以通过分泌相关生长因子控制自身的正常生理周期。这一结果可能与子宫肌的运动对生殖机能的影响、提供胎儿生长发育的环境、构成胎儿发育的环境等生理功能存在联系。因此推测,RBM3参与了雌性牦牛的子宫生理周期的调控以及妊娠识别等过程。
4 结 论
本研究成功克隆了牦牛3基因,登录号为:MF142258.1,3基因ORF全长为483 bp,编码18种、160个氨基酸。预测RBM3蛋白存在17个磷酸化位点,是稳定的非跨膜蛋白。RBM3蛋白及基因在雌性牦牛发情周期不同阶段的卵巢、子宫、输卵管中均有表达,但存在差异。本研究结果提示,RBM3参与了牦牛发情周期以及妊娠过程的调控。
