从江及周边地区稻田鲤的遗传结构及系统发育关系
2022-08-04余科麻智芳安苗罗均黄胜梁志强邵俭潘秋芝任芳龙金梅
余科 麻智芳 安苗 罗均 黄胜 梁志强 邵俭 潘秋芝任芳 龙金梅
(1.贵州大学动物科学学院,高原山地动物遗传育种与繁殖教育部重点实验室,贵州省动物遗传育种与繁殖重点实验室,贵阳 550025;2.贵州省黔南州农业科学研究院畜牧水产所,都匀 558000;3.贵州省从江县农业农村局,从江 557400;4.湖南省水产科学研究所,长沙 410153;5.贵州省锦屏县农业农村局,锦屏 556700)
鲤(Cyprinus carpioLinnaeus)是一种有着几千年的文化史和驯养史的鱼类,在全球100多个国家进行养殖,产量居世界的第四位和中国养殖鱼类的第四位[1,2]。鲤亚种的分类及系统发育关系多有争议,Kirpichnikov[3]基于形态差异提出外高加索鲤C.c.carpio、中亚鲤C.c.aralensis、远东鲤C.c.haematopterus和东南亚鲤C.c.viridiviolaceus四个亚种,而后外高加索鲤和中亚鲤被证实有共同祖先[4],合用C.c.carpio一个亚种名[5]。东南亚鲤的有效性同样受到质疑[6],Steffens[7]认为它可能来自中国。Balon[8]基于形态和生理特征只认可欧洲鲤和东亚鲤2个野生亚种,伍献文[9]基于形态和地理分布差异,提出外来的西鲤、远东鲤和华南鲤3个亚种,并成为我国鲤鱼资源调查中关于形态和分类的主要依据,分子遗传学研究也支持华南鲤是远东鲤分离的一个亚种[10—12]。由于地理隔离、适应性突变积累、自然和人类的选择压力综合作用,鲤已经发展出许多种群和品系[13],我国拥有世界上数量最多的鲤天然种群、驯化品系和人工品种[14],其中从江田鱼C.c.congjiangensis[15]、湖南呆鲤[16]、广西金边鲤C.c.var.jinbian[17]、晒江鲤[17]和桂林禾花鲤[18]等地方性稻田鲤备受人们关注,并在传统稻鱼共生系统中发挥着重要作用。
从江县隶属贵州省黔东南苗族侗族自治州,地处都柳江中下游,周边与榕江、黎平、广西融水和三江等县接壤,东北部与锦屏、湖南的通道和靖州等县比邻,是我国南方湘黔桂苗、侗族聚居的核心区,少数民族风情独特,稻田鲤驯养已有上千年[19,20]。根据外形和体色有乌鲤、荷包鲤和火鲤等当地俗称品种[19,21],它们具有性情温顺、抗病力强、活动力弱和不善跳跃等特点,在稻田浅水环境不易逃遁,统称“呆鲤”或“田鱼”,又因以稻田禾花为食而称“禾花鱼”,现今许多村寨现仍延续着自繁自养自留种的繁养体系。本地鲤与糯禾构成了一种稻鱼共生和鱼米同收的生态农业模式,2011年从江侗乡稻鱼鸭复合系统获联合国“全球重要农业文化遗产”授牌,融水田鲤和从江田鱼等先后获得全国农产品地理标志,极大地促进了稻田鲤的养殖利用。然而,随着现代农业技术的应用和大量外来鲤品种涌入[17,22,23],稻田本地鲤的生存受到严重威胁[24,25],种质面临混杂风险。
目前,从江及周边苗侗族地区稻田鲤的种质特性研究步伐不断加快,袁振兴[26]研究了都柳江野鲤的遗传多样性,郭梁等[15]将从江田鱼与青田田鱼C.c.qingtianensis进行了形态学对比分析,两种“田鱼”在形态上明显区别于其他鲤鱼种群或品系,广西稻田鲤的遗传结构的研究也取得了一定成果[17,18,27,28]。然而从江及周边苗侗族地区稻田鲤的遗传背景我们还知之甚少,分类地位及其系统发育关系仍不确定,严重阻碍了其种质保护和育种利用。系统发育分析可阐明鱼类种群之间系统发育关系,揭示种群和品系鲜为人知的混杂史,有助于遗传育种和种质保护工作[14],mtDNA是系统发育研究中最常用的遗传工具,细胞色素b(Cytb)是系统发育分析中最常用的基因区[11]。为此,本文基于线粒体Cytb基因标记技术,对从江及邻近6个县稻田鲤的遗传组成及系统关系开展研究,揭示传统稻鱼复合生态系统中鲤的种质特征,旨在为鉴定与评价这一独特的农业文化遗产提供理论依据。
1 材料与方法
1.1 实验材料
稻田鲤分别采自贵州、广西和湖南3省的从江、榕江、黎平、锦屏、三江、靖州和通道等7个县22个乡镇28个村寨(表1和图 1)。其中,从江县全境设置12个乡镇16个村寨;榕江、黎平和锦屏3县设置6个乡镇7个村寨;广西三江县设置2个乡镇3个村寨;湖南的靖州和通道2县设置2个乡镇2个村。每个村寨尽量选取有自繁自育自留种习惯的1家农户,放低稻田水位徒手捉鱼,根据捕获鲤鱼多寡从中随机挑选尾数不等的活鱼样品。现场测量体长和体重等常规生物学指标,取背部肌肉3—5 g浸泡于95%乙醇溶液试管中,置于保鲜冰盒回校后放入-80℃冰箱中保存备用。

图1 采样点Fig.1 Sampling Site

表1 样品采集信息Tab.1 Sample collection information
1.2 实验方法
提取DNA 利用DNA提取试剂盒(北京天根)提取肌肉样品基因组DNA,通过1.0%琼脂糖凝胶电泳检验其完整性,再利用紫外分光光度计(Thermo Nano Drop 2000C)测定其吸光值。最后置于-20℃保存备用。
引物的筛选与PCR的扩增Cytb扩增引物的序列为L14724:5′-GACTTGAAAAACCACCGTTG-3′,H15915:5′-CTCCGATCTCCGGATTACAAGAC-3′[29],送至生工生物工程(上海)股份有限公司合成。
PCR反应体系为25 μL:模板DNA 2.5 μL,2×TaqPCR Master Mix 12.5 μL,上、下游引物分别为1.25 μL,剩余体积用双蒸水补齐。反应程序:94℃预变性3min,94℃变性40s,52.5℃退火40s,72℃延伸50s,重复35个循环,最后再72℃终延伸7min,4℃保存备用。通过1.0%琼脂糖凝胶电泳检验,选择特异性强的样品送至生工生物工程(上海)股份有限公司进行双向测序。
1.3 数据处理与分析
测得的上下游序列利用DNAStar5.0软件包中的SeqMan进行拼接并手工校对。从NCBI上下载广西禾花鲤(MK088487)完整Cytb基因序列,比对保留相同序列长度。运用Mega6.06软件[30]对不同序列间的碱基组成进行统计,基于K2-P模型计算群体间的遗传距离[31]评价遗传差异,收集NCBI基因库中国内外鲤的Cytb基因序列与本研究单倍型序列,基于K2-P模型构建不同单倍型间的NJ(Neighbor-Joining)和MP(Maximum parsimony)系统进化树,同时运用MrBayes 3.2.7[32]构建贝叶斯系统树(Bayesian inference,BI),循环运算设置10000000代,每隔1000代进行样本抽样,运算停止后用TreeGraph 2.15.0[33]绘制系统发育图。利用Network5.0软件[34],以中介邻接网络算法(MJ,Median-joining networks),按照单倍型频率绘制神经网络图,展示单倍型之间的衍生和进化关系。用DnaSP 5.10软件计算单倍型数(Number of haplotypes,H)、单倍型多样性(Haplotype diversity,Hd)、核苷酸多样性(Nucleotide diversity,π)、多态位点数目(Number of polymorphic sites,S)和平均核苷酸差异(Mean number of pairwise differences,K)等多样性评价参数。运用Arlequin3.5软件[35]进行变异分析(Analysis of Molecular Variance,AMOVA),检测遗传变异来源,并计算群体间的分化系数。
2 结果
2.1 稻田鲤Cyt b基因的碱基组成和单倍型分布
447条长度为1141 bp的Cytb基因同源序列,共检测出54个变异位点,无碱基的插入和缺失,有5个颠换和49个转换,转换与颠换比为11.23,转换主要发生在碱基G与A之间,占总变异的48.1%,略高于T与C转换比的42.6%。碱基T(U)、C、A、G的平均含量分别是26.3%、29.9%、29.9%、14.0%,A+T的平均含量为56.2%大于 C+G 的平均含量43.9%。共界定了29种单倍型(MW683933—MW 683961),单倍型的分布见表 2,其中独有的单倍型18个,共享的11个。从江群体拥有最多的单倍型(19)和独享单倍型(8),与周边5个稻田群体有11个共享,Hap1、hap3-hap5和hap8是其优势单倍型。榕江群体无独有单倍型,广西三江有1个,黎平有4个,锦屏2个,湖南有3个。Hap3、hap5、hap8、hap11、hap15和hap16在6个稻田鲤群体间广泛分布,代表296尾个体,占总样本数的66.22%。
2.2 稻田鲤的系统发育关系及单倍型神经网络图
以鲫Carassius auratus auratus和三角鲤C.multitaeniata为外群,29种单倍型与欧亚鲤的39条Cytb基因序列构建NJ、MP和BI系统发育树,聚类结果基本一致,其中MP系统树见图 2。
68条鲤Cytb基因序列(1124 bp)以100%置信度聚在一起,严格划分为2大谱系(Clades)。Clade A由4个单倍型和欧洲鲤的4条序列以98%置信度聚类,下分2个亚支subclades(a1和a2),hap22及其衍生的3个单倍型组成a2,它们隶属欧洲鲤亚种。Clade B由剩余的25个单倍型和亚洲的35条鲤序列以64%置信度构成,其下分为7个亚支,其中b1、b3、b4和b5亚支中含17个单倍型,它们分别与珠江野鲤、元江野鲤、都柳江野鲤、红河的华南鲤、广东大肚鲤Big-belly carp、金边鲤和桂林禾花鲤等聚类,分类上属华南鲤亚种;b2、b6和b7亚支中8个单倍型与日本鲤霞浦湖鲤、长江野鲤、兴国红鲤C.c.var.xingguoensis、荷包红鲤C.c.var.wuyuanen-sis、日本锦鲤C.c.Koi、青田田鱼和俄罗斯散鳞镜鲤Russian scattered scaled mirror carp聚类,属远东鲤亚种。
结合表 2统计,欧洲鲤亚种4个单倍型鱼的33尾样品分布于除锦屏外的5个群体,三江(15)和从江(10)的数量最多;远东鲤亚种8个单倍型鱼的44尾样品在6个群体都有分布,从江(26)和湖南(9)的数量最多;华南鲤亚种17个单倍型鱼的370尾样品在6个群体也有分布,且都是其优势的类群。

表2 单倍型分布Tab.2 Haplotype distribution
29个单倍型的神经网络图(图3)划分为三个区(Ⅰ、Ⅱ和Ⅲ):Ⅰ区与Clade A对应,与其他2个区之间的突变步骤较长,湖南通道的hap22进化出另外3个单倍型,在从江、榕江、黎平和三江扩散开来。Ⅱ区对应Clade B的b1亚支,由从江和黎平各自2个独有单倍型组成,因3个未知的单倍型缺失而在该区相互独立分布。Ⅲ区是一个庞大的家族,Hap5和hap18与其余单倍型之间因存在1—2个缺失的单倍型,而略显独立,与图 2中Clade B的聚类情况一致,剩余的单倍型都以从江的Hap7为网络中心向外辐射发展进化而来,显示其祖先的等级。
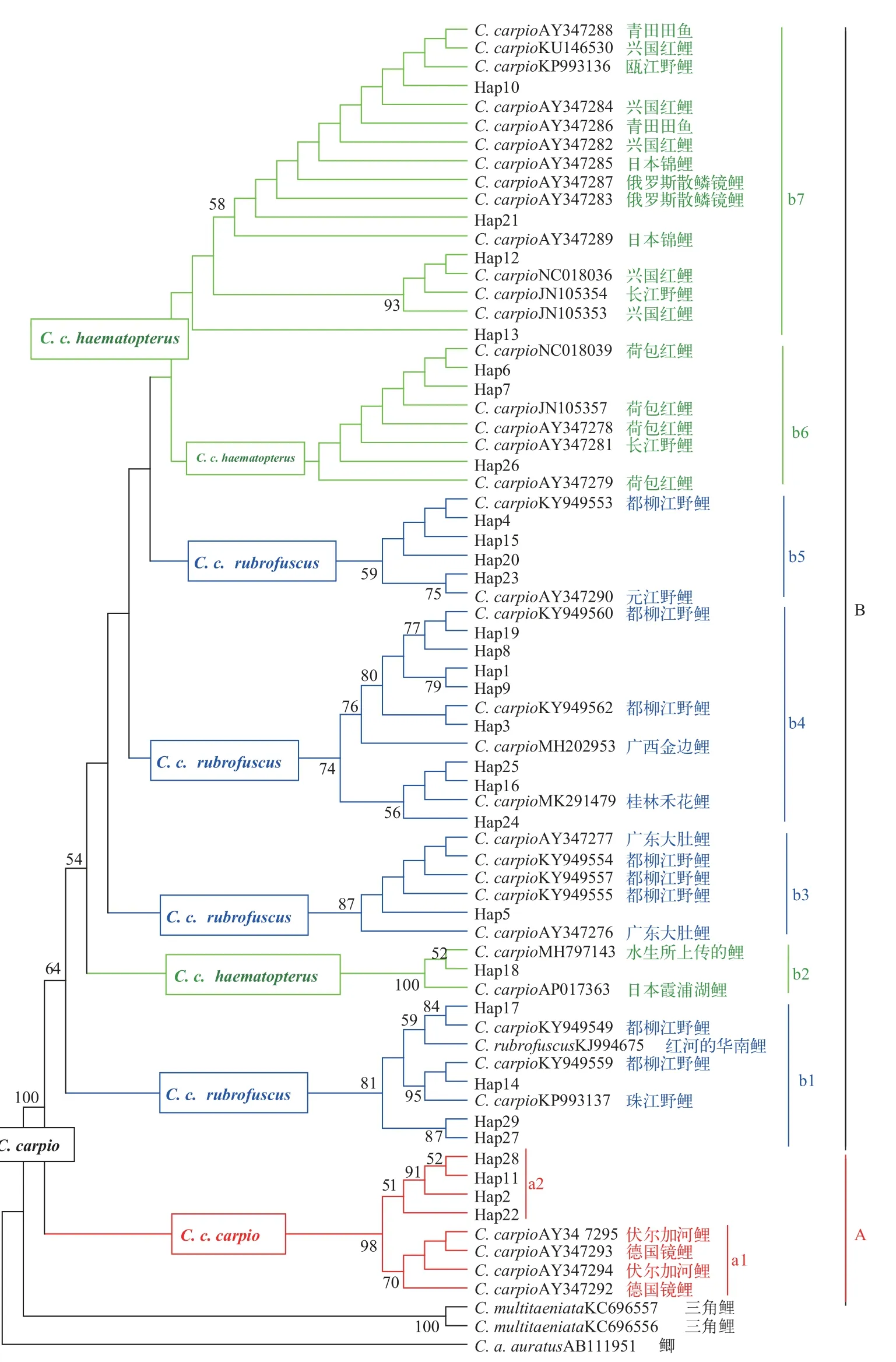
图2 鲤mtDNA Cyt b基因单倍型的MP系统发育树Fig.2 MP Phylogenetic tree of mtDNA Cyt b gene haplotypes in C.carpio
2.3 稻田鲤的遗传多样性
如表 3所示,欧洲鲤为低Hd和低π,其遗传多样性值远低于亚洲的鲤鱼类群,亚洲的2个鲤亚种群体以华南鲤遗传多样性最高。在华南鲤群体中,从江具有最高的多态位点数、单倍型数和单倍型多样性,黎平具有最高的核苷酸多样性,锦屏的Hd和π都最低。
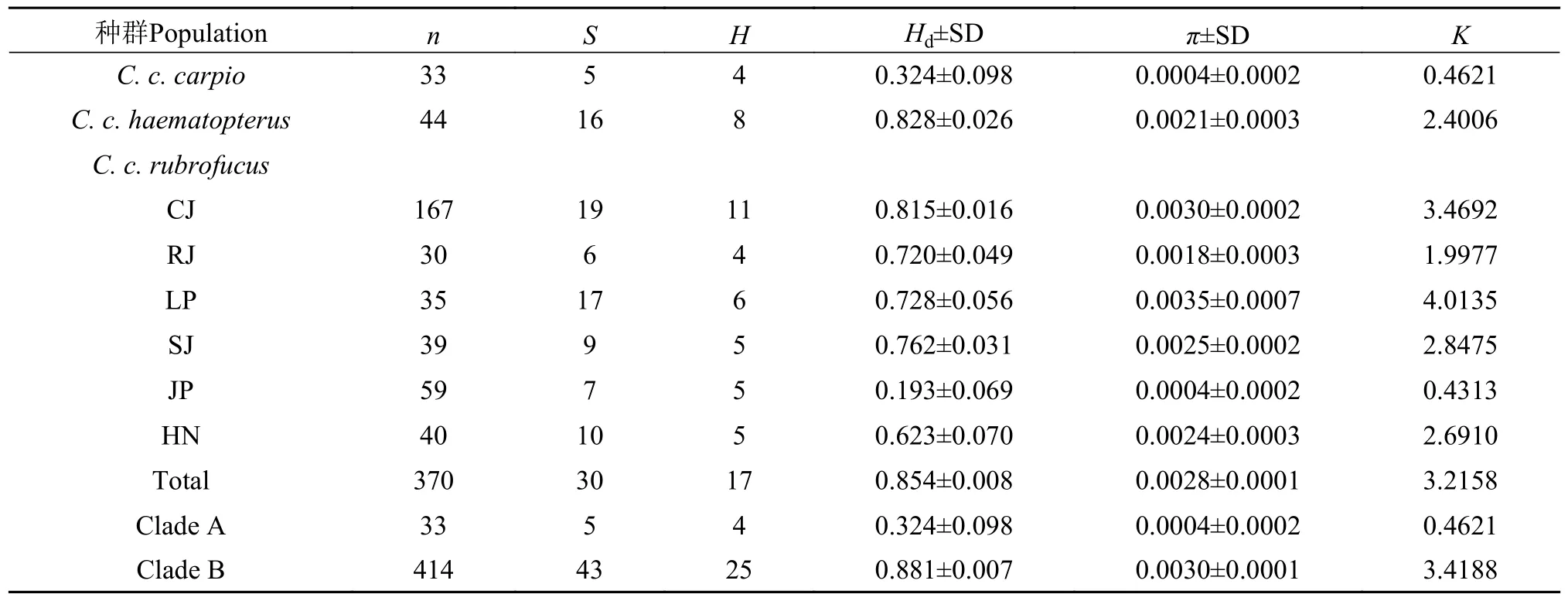
表3 稻田鲤群体的遗传多样性Tab.3 Genetic diversity of rice field carp populations
2.4 稻田鲤群体间的遗传差异及遗传分化
如表 4所示,6个华南鲤稻田群体间的遗传距离为0.0016—0.0036,以黎平与从江和三江之间差异最大,其次是三江与从江和湖南,湖南与从江和黎平以及榕江与锦屏之间差异最小。遗传分化指数在0.0345—0.4233,锦屏与其他群体之间的遗传分化最大,其次是三江群体,其他群体间的数值较小。2个谱系间的遗传距离为0.0129,亚种间为0.0128、0.0129和0.0038,说明谱系间或亚种间的遗传差异水平高于6个华南鲤群体。谱系间或欧洲鲤与远东鲤和华南鲤之间分化值最高,远东鲤与华南鲤之间最低。

表4 稻田华南鲤群体间的遗传距离和遗传分化指数Tab.4 Genetic distance and genetic differentiation index among the rice field carp populations (C.c.rubrofucus)
6个华南鲤群体按省划分3组:第1组为贵州省的从江、榕江、黎平和锦屏4个群体,第2组为湖南群体,第3组为广西三江群体,其AMOVA结果见表 5。组间的变异贡献率为2.17%,群体间为16.69%,群体内为81.14%。这说明稻田华南鲤的遗传变异主要来源于群体内个体间,其次才是地理群体间。遗传分化指数Fst=0.167,并且差异极显著(P<0.0001)。

表5 基于mtDNA Cyt b序列的分子方差分析Tab.5 Molecular analysis of variance (AMOVA) based on mtDNA Cyt b sequence (AMOVA)
3 讨论
3.1 稻田鲤的亚种地位及其母系血统来源追踪
我国3个鲤亚种主要是分布于新疆额尔齐斯河流域的西鲤、黑龙江至闽江的远东鲤及珠江、元江、海南岛等地的华南鲤[9]。系统发育树显示,本研究29种单倍型严格划分欧洲和亚洲2个地理谱系,Clade A中4个单倍型与欧洲鲤聚类,blast比对也发现,hap22与希腊鲤(EU689059和DQ868871)和伏尔加河鲤(AY347295)的相似度为99.91%,另外3个单倍型与其相似度为99.56%—99.74%,表明该4种单倍型隶属欧洲鲤类型。我国自20世纪50年代起陆续引进散鳞镜鲤C.c.var.specularis amurensis和德国镜鲤C.c.var.specularis germanensis等欧洲驯养鲤[36,37],以它们为母本获得芙蓉鲤C.c.“Furong”和颖鲤C.c.“Ying hybrid”等人工品种,本地区也有引种养殖的历史[22,23,36]。4种单倍型鱼33尾样品在锦屏外其他5个稻田群体都有分布,揭示从江及邻近地区稻田鲤受到欧洲驯养鲤母系基因的遗传渗析。Clade B划分7个亚支,b2、b6和b7中的8个单倍型鱼与长江野鲤、青田田鱼、兴国红鲤、荷包红鲤、日本锦鲤和俄罗斯散鳞镜鲤等远东鲤的类群或品系[11]聚类,表明其远东鲤亚种地位。Hap18与日本霞浦湖鲤和水生所上传鲤的序列相似性达100%,Mabuchi等[38]认为霞浦湖鲤系引进的欧亚品系,该单倍型鱼只有1个个体,可能为输入型。Hap6、hap7和hap26分别与荷包红鲤的序列相似度达99.82%—100%,3个单倍型显示是荷包红鲤的母系起源。Hap10、hap12和hap21与长江野鲤和兴国红鲤相似度为99.91%—100%,hap13与荷包红鲤相似度为99.74%。8个单倍型鱼与兴国红鲤和荷包红鲤的同源性较高,预示其相同的母系来源,该地区有兴国红鲤和荷包红鲤及其杂交品种(如丰鲤、荷元鲤、岳鲤和建鲤等)的推广养殖历史[22,39]。8个单倍型鱼的44尾样品在6个稻田鲤群体中都有发现,说明本地区稻田鲤同样遭受国内鲤养殖品种的母系遗传物质渗入。
b1、b3、b4和b5显示是华南鲤亚种地位及其地理种群或地方品系的母系血统。b1亚支中4个单倍型在神经网络图中(图3)处于独立的Ⅱ区,系统树中也相对独立,与其他亚支关系较远,但它们与珠江、都柳江和红河的鲤鱼以81%置信度聚类在一支,归属华南鲤亚种是比较合适的,Hap14和hap17分别与珠江野鲤(KP993137)和都柳江野鲤(KY949559和KY949549)的相似性达100%,Hap27和Hap29与其相似度最高只有99.47%而独立为小支,4个单倍型鱼与都柳江野鲤高度同源。Hap5与广东大肚鲤和都柳江野鲤序列相似度达100%,Zhou等[10]指出广东大肚鲤和元江野鲤都属华南鲤亚种,Hap5代表的32尾鱼以从江的21尾最多,其次是榕江和湖南通道各4尾,广西三江2尾和湖南靖州1尾最少,本支中也含都柳江野鲤的3个单倍型鱼,它是否为本地区所有或来自广东还不能确定。b4中含有多达8个单倍型,Hap16、hap24和hap25分别与桂林禾花鲤相似度最高(100.00%、99.91%和99.74%),hap3和hap8与都柳江野鲤(KY949560和KY949562)的相似性最高(100%),与金边鲤为99.91%和99.82%,另外3个单倍型与都柳江野鲤的序列相似性最高为99.91%、99.91%和99.82%,其次才是与金边鲤(99.82%、99.74%和99.74%)。潘贤辉等[18]研究指出桂林禾花鲤和金边鲤群体关系较近,推测由珠江水系右江的野生鲤演变而来,程磊等[28]指出广西禾花鲤母系起源上属华南鲤。8个单倍型鱼与桂林禾花鲤、都柳江野鲤和融水金边鲤有相同母系血统来源,3地鲤鱼分布区彼此邻近,民族同根同源,文化、习俗和稻作方式相同,有稻田鲤的苗种交流与养殖推广报道[17,19,27,40],前3个单倍型鱼(92尾样品)分布在三江外的5个稻田群体中,主要分布于桂林禾花鲤分布区全州县邻近的锦屏(55/60)、黎平(16/43)和湖南 (11/50),后5个(有185尾样品)广泛分布3省7个县的稻田中,主要集中在都柳江流域的从江(115/203)、三江(13/55)、榕江(18/36)和黎平(13/43)四地,说明b4亚支是本区域稻田中分布最集中的华南鲤类群。b5亚支中的hap4、hap15、hap20和hap23与都柳江野鲤和元江鲤聚类,相似度为99.82%—100.00%,四个单倍型鱼也属华南鲤亚种,51尾样品鱼主要分布于从江(23尾)和三江(22尾)。

图3 单倍型的神经网络图Fig.3 Neural Network Diagram of haplotypes
3.2 稻田鲤群体的遗传结构
稻田鲤遗传多样性统计表明,分布于稻田中欧洲驯养鲤群体为低Hd低π,多样性匮乏。虽然本研究样本数有限,但前期关于欧洲鲤mtDNA多个基因区分析也同样表明其遗传变异程度极低[4,41,42],推测欧洲养殖和驯养鲤的早期存在瓶颈效应或奠基者效应[12],导致遗传变异的局部丢失[8]。稻田中远东鲤和华南鲤2个亚种群体都为高Hd低π,Thai等[42]研究也指出中国鲤与欧洲鲤相比,其进化历史更长,表现出相当高的单倍型多样性和差异性,本研究稻田鲤的2个亚种群体也有同样特点。稻田中最大的华南鲤群体也显示是多个母系血统混合体呈高Hd(0.854),说明多个母系血统来源的鲤品系混养和缺乏人工定向选育,能较大程度保持现有稻田鲤的基因型多样性[28],然而锦屏群体的低Hd低π有点例外,统计发现59尾样本中有53尾归属hap16(启蒙镇便幌村21尾、启蒙镇岑根村14尾和敦寨镇螺丝田村18尾),启蒙镇便幌村是一个典型自繁自育自留种的稻田鲤养殖点,样品鱼遗传同质化,提示为创始者事件和瓶颈效应的结果,Perdices等[43]指出广泛的采样可能会发现更多的线粒体谱系,因此锦屏稻田鲤群体遗传多样性正确评价有待今后扩大采样范围。从江群体具有广泛采样和大样本量,其单倍型数最多,Hd也最高,但其π仍偏低,5个地理群体高Hd低π特点属鱼类多样性第二种类型 (h>0.5,π<0.005)[44],遗传多样性水平与该物种的进化速率正相关[45],Zhou等[11]提出华南鲤与远东鲤的分歧时间在中更新世,由于进化历史相对较短,加之经历过较严重的遗传瓶颈[46]和种群扩张[47],说明华南鲤在经历瓶颈效应后,进化快速,积累了丰富的单倍型多样性,但核苷酸突变积累略显不足。
本研究稻田鲤的谱系或亚种间遗传分化水平存在明显差异,Clade A与Clade B间的遗传距离(0.0129)和遗传分化指数(Fst=0.7934,P<0.001)都显示出较大的遗传差异和显著的分化,欧洲鲤与亚洲的2个亚种鲤间同样有较高遗传差异(0.0128和0.0129)和分化水平(0.8902和0.8058),这符合欧洲和亚洲2个鲤群体古老分离的观点[10]。亚洲的2个鲤亚种间遗传距离(0.0038)和遗传分化(0.3258)明显偏低,显示在系统树中2个亚种未能独立成单系群,Zhou等[11]认为这种低水平遗传差异能与其分歧时间较短有关。本研究6个稻田华南鲤群体间的遗传分化指数为0.0345—0.4233,其中锦屏与另外5个群体间处于高度-中度分化等级[48],三江与剩余4个群体也处于高度-中度分化等级,其余群体间为低度或无分化等级。华南鲤有17种单倍型,虽然共享单倍型有8种,且样品数占比较高(342/370),但共享单倍型数和样品数都集中分布在邻近的2—3个地理种群间(如从江多达160尾),说明共享单倍型鱼在6个群体间的传播是有限的,而独有单倍型比例高达半数以上(9/17)。AMOVA分析显示,群体间遗传变异贡献率为16.69%,处于中等遗传变异,说明群体间及其内部均各自具有相互隔离的遗传结构[27]。这可能与各地理群体存在不同的创始群体、不同的选择压力和地理隔离等限制其基因流动的结果[49]。
3.3 稻田鲤遗传资源的保护和研究
程磊等[22]基于D-loop区和Cytb基因在广西禾花鲤中观察到少量欧洲鲤和远东鲤的入侵,本研究基于mtDNA Cytb基因的分子系统发育结果同样发现从江及邻近苗侗族地区稻田鲤受到欧洲鲤和远东鲤驯养品系的入侵,样品数占总样本量的17.23%(77/447),说明本地区稻田鲤遭受国内外鲤驯养品系混杂的程度较为严重。因此,加强外来人工品种在稻田推广养殖的监管和追踪十分必要。
另一方面,线粒体基因组中不同的基因和区段具有不同的进化速率,在系统发育分析中有时会产生显著的差异[14],当联合mtDNA多个基因序列时,系统发育关系的分辨率和准确性会更高[50]。本研究借助Cytb基因的遗传分析,初步阐明了从江及邻近苗侗族地区稻田鲤的系统关系、遗传组成及种群结构,获得了第一手宝贵资料,但是否遭受国内外鲤品系父系基因遗传渗析仍不得而知,需进一步联合多个核内外基因的检测和分析技术,综合评价该区域稻田鲤的种质特征。
