珍稀蝴蝶的亚种分类问题及保护意义:以喙凤蝶属为例
2021-12-08曾菊平姜梦娜周善义张江涛
邹 武,曾菊平,姜梦娜,王 渌,周善义,张江涛,3
(1.江西农业大学林学院,鄱阳湖流域森林生态系统保护与修复国家林业和草原局重点实验室,南昌 330045;2.广西师范大学生命科学学院,珍稀濒危动植物生态与环境保护教育部重点实验室,广西桂林 541006;3.江西九连山森林生态系统定位观测研究站,江西龙南 3441700)
“物种(species)是客观存在的”已被人们广泛接受,这也是生命科学研究的热点议题(Darwin,1859;Mayr,1963,2001;陈世骧,1983)。而与之相反,亚种(subspecies)地位一直备受质疑(Wilson and Brown,1953;Mayr,1982),一方面,亚种概念多,说法不一;另一方面,亚种分类更易受主观因素影响。然而,在分类实践中,试图用不同术语代替亚种或完全放弃它的尝试都是不可接受的。在地理变异物种中,尤其是有明显地理隔离的物种,亚种分类仍是一种便利的方法(Mayr,1963)。
尽管,亚种不是一个进化单元(Mayr,1963),但在生物多样性保护中,亚种分类的科学性,会影响到相关保护策略的制定、保护方案实施及其成效(Ryder,1986)。目前,存在两种不同观点:一者认为“亚种”提供了一种近乎真实的种内分化地理格局印象,可能对保护努力(程度、方向)产生误导(Zink,2004);另者则认为“亚种”为评估种内遗传多样性模式提供了一个有效的捷径,可将其作为一种用于进化差异与保护研究的工具(Phillimore and Owens,2006)。在野外,珍稀蝶类多在特定的狭小范围内发生(周尧,1994;武春生和徐堉峰,2017),地理种群常呈“岛状”分布格局,彼此间隔离明显。这种格局为亚种确立提供了便利,但也可能诱导产生“过度亚种化”现象。尤其是,当形态描述、分子遗传等信息不足时,亚种分类更易受到主观任意性影响(Mayr and Ashlock,1991)。这里,我们以珍稀蝴蝶喙凤蝶属Teinopalpus为例,系统查阅历史资料、文献,从地理隔离、形态差异等分析珍稀蝶类亚种分类问题(如“过度亚种化”等)及其原因,并探析其对珍稀蝶种与多样性保护、管理的潜在影响,从种下分类层面为昆虫多样性保护研究提供参考。
1 亚种概念与分类
物种是指个体成员间可自由繁殖的生物群体,亚种是物种的一个次级阶元,是某个物种的表型上相似种群的集群,栖息在该物种分布范围内的次级地理区(Patten,2010;van Holstein and Foley,2020),亚种是物种之下的分类阶元,也是《国际动物全名法规》管理范围的最低一级单元。
1.1 亚种概念
当前,有关亚种有多种理解(Sylvester-Bradley,1951;Wilson and Brown,1953;Mayr,1982;Ryder,1986;Mayr and Ashlock,1991;Agapowetal.,2004;Patten,2010;van Holstein and Foley,2020),形成如下多个概念:
(1)地理亚种:指种内占据不同地理分布区或宿主、互不重叠、生殖隔离不完善且具有一定形态差异的生物类群。或者说,指的是与同种内其他居群在地理分布上界线明显、形态特征上有一定差异的居群(Wilson and Brown,1953;Mayr,1982;Mayr and Ashlock,1991)。
(2)年代亚种:在古生物学上,由于时代分布不同而使同一种内存在的在形态特征上具有一定差异的居群,称为年代亚种(Sylvester-Bradley,1951)。
(3)地方性种群:指一个分布广泛的种包括许多地方性种群,它们在基因频率、数量性状的平均值上有微小不同,其差异达到一定程度时可称为亚种。也就是说,亚种是地方性种群在地理上划分出的集合体,在分类上与本种的其他集合体互有不同(Wilson and Brown,1953;Mayr,1982)。
(4)地理宗与生态宗:亚种是在地理上局限分布的,相当于一个地理宗。与此同时,任何两地的地理环境都不一致,因而一个亚种也相当于一个生态宗。所以,地理宗与生态宗代表着亚种的两个不同方面(Wilson and Brown,1953;Mayr,1982;徐炳声,1998;张德兴等,2003)。
以上概念涵盖地理空间、进化时间、生态环境、形态等分类学特征与方面,它们有不同侧重。但是,除年代亚种外,地理亚种、地方性种群或地理宗概念都强调亚种地理分布的有限性或地方性,认为亚种间在地理空间分布上不同或相互隔离。此外,这些概念都认为亚种间在形态等方面应存在差异,尽管差异程度可能微小。
1.2 亚种特征
事实上,亚种包括以下主要特征:(1)亚种形成主要由地理隔离引起,如异域分布(allopatric distribution)(O′Brien and Mayr,1991),即在空间上不重叠;(2)亚种间有可鉴别特征,如呈现表型差异;(3)亚种间无生殖隔离,或隔离不完善,相互可交配、繁殖出可育后代;(4)在两亚种分布交错区可能存在中间过渡类型;(5)亚种自身也是一个集合阶元,由多个当地种群组成,它们在基因型、表型上也呈现轻微差异等(Wilson and Brown,1953;Mayr,1982;Mayr and Ashlock,1991;Agapowetal.,2004;Patten,2010)。然而,除了考虑以上特征外,亚种分类过程也可能受物种自身种群结构影响。例如,物种具有连续种群结构模式,分布中心常被一系列相邻种群占据,种群间因相互扩散而紧密连接,基因交流频繁,变异特征呈渐变性。此时,若比较位于物种连续体的对立两极种群,表型差异明显时,可考虑将两极种群分别确立为亚种。而当物种具有不连续种群结构模式,如因历史事件形成的岛屿种群结构或“岛状”分布结构,岛屿间基因交流少,此时可将多个具有表型差异的隔离种群确立为亚种(Wilson and Brown,1953)。
1.3 亚种分类、界定
根据亚种概念、特征,亚种分类时应参考两个原则:(1)地理隔离原则;(2)表型差异原则。地理隔离意味着进化的相对独立性,孕育着分化的可能性。表型差异则表示亚种间已呈现出一定的变异性。地理隔离是亚种区别于种阶元的重要特征,即亚种为异域分布(Wilson and Brown,1953;Mayr,1982),而种则为同域或异域分布(Wilson and Brown,1953;O′Brien and Mayr,1991)。然而,值得注意的是,地理隔离充分的种群间,其表型差异也可能很小。例如,一些亚种仅有一个单独鉴别性状(如颜色、大小、生长型等)差异,而其他性状相同或相似,不受地理隔离影响,呈现出异地同型亚种(polytopic subspecies)格局(Mayr and Ashlock,1991)。通常,在两个亚种分布交错区可能存在两者的中间类型,它们由混合个体组成,成为居间种群。此时,要判定居间种群的亚种归属则充满挑战。
然而,75%法则(Amadon,1949)或许能帮助应对。所谓的75%法则,就是要求一个种群在被赋予亚种地位之前,需确认在地理变异性性状上组成该种群的75%的个体能与其他种群的99%的个体相互区分,而其总体性状特征则可从现有样本(标本)直接推断出来(Amadon,1949)。所以,当居间种群中的混合组成的个体不满足75%法则,可对其亚种地位做出否定判断。
2 喙凤蝶属亚种分类与问题
喙凤蝶属目前确认有2个蝶种(姊妹种)(周尧,1994;武春生和徐堉峰,2017),即金斑喙凤蝶T.aureusMell,1923与金带喙凤蝶T.imperialisHope,1843。两姊妹种生物地理上均来自东洋区,呈现同域分布特征。其发生范围主要沿我国武夷山、南岭向西延伸至喜马拉雅山脉周缘、向南延至中南半岛(含越南、老挝、缅甸、泰国等国),为亚洲特有分布属。发生区覆盖热带、亚热带林区,但主要栖息地位于中山地带的阔叶林区。当前,喙凤蝶在野外通常个体罕见,成虫只在少数地段、山头活动,生境选择性明显(Igarashi,1987,2001;曾菊平,2005;曾菊平等,2008,2012,2014)。例如,成虫活动对海拔高度有要求,山顶行为常在海拔>1 000 m的山脊及附近区域发生(Igarashi,1987,2001;曾菊平等,2012,2014),属典型的山地蝶种。在蝴蝶自身生境选择作用下,以及低海拔地带人类活动与土地利用影响下,喙凤蝶两姊妹种地理种群被限制在少数山区,相互隔离、孤立,呈“岛状”分布。这种发生、分布格局符合地理隔离原则,有利于亚种分类与界定,如金斑喙凤蝶武夷亚种T.aureuswuyiensisLee(李传隆和朱宝云,1992)主要基于武夷山脉地理种群确立,而海南亚种T.aureushainaniLee(周尧,1994)则包含了海南岛上的地理种群。但与此同时,若仅依据地理隔离原则,而忽视表型差异,则易出现“过度亚种化”问题。
1843年,Hope最早根据来自孟加拉国锡尔赫特的雄蝶标本,对喙凤蝶属Teinopalpus(T.imperialis)进行分类记述(Hope,1843)。此后,研究学者相继根据不同来源的标本分别记述喙凤蝶属金带喙凤蝶T.imperialis8个亚种,以及另一姊妹种金斑喙凤蝶T.aureus及其8个亚种。表1显示,除缺失数据外,亚种分类正模包括雄蝶(8头)与雌蝶(5头),副模则均为雄蝶标本。在数量上,除T.a.shinkaii亚种分类基于多年累积的47个副模标本外(Morita,1998),其余亚种分类副模都在4个以内,其中,3个亚种仅1个副模(Hope,1843;Turlin,1991;Collard,2007)。
2.1 基于行政区划的亚种分类应用与不确定性
从文献收集到133条有关喙凤蝶属亚种分类与应用记录,其中,多数应用能明确亚种名称、归属,但也有一定数量的不确定亚种归属情况(图1)。另外,少数记录提出了有争议的亚种分类问题,包括金带喙凤蝶与金斑喙凤蝶都存在这类问题。例如,在行政区划上,金带喙凤蝶在中国四川记录过T.i.imperialis,T.i.himalaicus与T.i.behludinii3个亚种名;老挝桑怒以及泰国清迈和楠府也均记录过2个亚种。同样地,金斑喙凤蝶在中国浙江以及越南永福、大叻和高平也有过2个不同亚种记录。不仅如此,在泰国帕翁坡山(T.i.imperialis与T.i.bhumipholi)、越南Pia Oac (T.a.aureus与T.a.shinkaii)2个发生点,也分别有过金带喙凤蝶与金斑喙凤蝶的2种亚种记录(图1)。

图1 喙凤蝶属Teinopalpus部分亚种分类应用文献记录(1843-2014年)特征Fig.1 Classification features of some subspecies of Teinopalpus recorded in literatures (1843-2014)A:亚种分类文献记录(n=66)的确定/不确定性Confirmed or uncertain records (n=66)in subspecies classification;B:T. aureus亚种分布的同域记录 (n=72)Sympatric records of subspecies of T. aureus (n=72);C:T. imperialis亚种分布的同域记录 (n=133)Sympatric records of subspecies of T. imperialis (n=133).
同一发生点上出现不同亚种名称的应用,可能与亚种分类时间早晚有关,后期应用中容易得到校正。但在同一行政区内,不同亚种名称的应用可能是亚种分类混乱或区内“过度亚种化”的信号反馈,需引起注意。例如,需进一步采集分类证据,包括形态、分子等证据,对现有亚种分类进行修订。
2.2 基于生态区的地理隔离分析
尝试从生物地理与生态区上来分析喙凤蝶属各亚种地理隔离,结果显示:金带喙凤蝶8个亚种分布范围覆盖着3个植被群落(biome)(Olsonetal.,2001),包括位于温带阔叶混交林(temperate broadleaf and mixed forests,TBMF)的T.i.himalaicus与T.i.miecoae亚种,位于温带针叶林(temperate conifer forests,TCF)的T.i.behludinii,以及位于热带亚热带湿润阔叶林(tropical and subtropical moist broadleaf forests,TSMBF)的T.i.imperialis,T.i.imperatrix,T.i.bhumipholi,T.i.gillesi和T.i.gerritesi5个亚种(表1;图2)。然而,记述的金斑喙凤蝶8个亚种则均在TSMBF植被群落发生(表1)。而进一步参考Olson等(2001),在更低生态区(ecoregion)层面比较,可知金斑喙凤蝶亚种归入4个生态区(图2),其中,T.a.shinkaii与T.a.laotiana同区,T.a.aureus,T.a.wuyiensis,T.a.guangxiensis与T.a.nagaoi同区,而T.a.eminens与T.a.hainani则各占一个生态区。图2显示各亚种所在植被群落或生态区与其地理位置密切相关,分布越近处于相同植被群落或生态区的概率更高,因而可从亚种所在植被群落或生态区层面,判定是否遵从地理隔离原则。显然,金带喙凤蝶8个亚种地理隔离相对金斑喙凤蝶的亚种更明显,但从生态区层面,T.i.imperatrix与T.i.bhumipholi较其他亚种更为相似,可考虑合并,且根据优先律,统一用T.i.imperatrix亚种名。同样地,对于金斑喙凤蝶而言,按生态区相似法与优先律,也可考虑将T.a.eminens与T.a.laotiana合并,统一用T.a.eminens亚种名,而将T.a.wuyiensis,T.a.guangxiensis与T.a.nagaoi合并到指名亚种T.a.aureus。
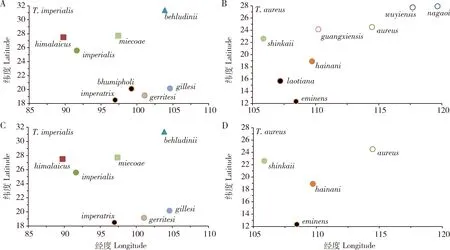
图2 金带喙凤蝶(A,C)和金斑喙凤蝶(B,D)亚种修订前后的经纬度分布及其植被群落和生态区Fig.2 Longitude and latitude distributions of subspecies of Teinopalpus imperialis (A,C)and T. aureus (B,D) and related biomes and ecoregionsA,B:修订前Before revision;C,D:修订后After revision.符号相同表示植被群落一致,生填充色相同表示生态区一致(详见表1),植物群落与生态区划分参考Olson等(2001)。Identical symbols indicate the same biomes,and identical filling-colors indicate the same ecoregions before or after revision (see Table 1 for details).The division of biomes and eco-regions referred to Olson et al.(2001).

表1 金带喙凤蝶和金斑喙凤蝶亚种分类文献及其正/副模来源与所属生态区Table 1 Taxonomic notes on subspecies and holotype/paratype specimens’localities and eco-regions in the subspecies of Teinopalpus imperialis and T. aureus
因此,根据以上地理隔离分析,金带喙凤蝶亚种分类建议修订为T.i.imperialis,T.i.himalaicus,T.i.miecoae,T.i.behludinii,T.i.imperatrix(含T.i.bhumipholi),T.i.gillesi和T.i.gerritesi;金斑喙凤蝶亚种分类则修订为T.a.aureus(含T.a.wuyiensis,T.a.guangxiensis与T.a.nagaoi),T.a.eminens(含T.a.laotiana),T.a.shinkaii与T.a.hainani。
2.3 亚种分类与表型差异
据优先律原则,新亚种确立应与已知的指名亚种进行形态比对。然而,表2和3显示,除与指名亚种形态比对外,喙凤蝶属亚种分类也通过与周边其他亚种比对来确立亚种地位。而可用标本数量(表1)会影响形态比对结果,尤其喙凤蝶属亚种形态比对大多局限在单种性别上(雄或雌性)(表2和3)。显然,75%法则因为数量限制而未能用到喙凤蝶属亚种分类上。当前,其亚种形态比对主要针对前翅背面(upper forewing,Upf)、前翅腹面(under forewing,Unf)、后翅背面(upper hindwing,Uph)等的特定区域[如中区或盘区(median/discal area)、亚缘区(submarginal)、中后区或盘后区(post-medial/post-discal area)、翅脉间等]的颜色、带形、斑纹及其变化,以及翅展等进行,通过寻找形态上的微小差异来确定亚种分类方向(如新亚种确立、同物异名判定等,见表2和3)。

表2 金带喙凤蝶部分亚种形态比对原文描述Table 2 Morphological descriptions and comparisons between subspecies in Teinopalpus imperialis
续表2 Table 2 continued
例如,金带喙凤蝶亚种T.i.miecoae与T.i.imperialis雄虫形态差异描述为:T.i.miecoae后翅背面黄斑较T.i.imperialis宽,T.i.miecoae臀角区颜色最暗,臀角区黄斑小等,而T.i.miecoaee与T.i.bhumipholi雄虫形态差异描述为T.i.miecoae后翅背面黄斑较T.i.bhumipholi窄,T.i.miecoae臀角区颜色最暗,臀角区黄斑小等(表2),意味着雄虫后翅背面(Uph)金黄色斑块(yellow section)在大小上存在连续而微小的变异。又如,金斑喙凤蝶亚种T.a.shinkaii与T.a.guangxiensis形态差异主要为前者雄虫翅展大于后者,而与亚种T.a.eminens形态差异也主要为前者雄虫翅展小于后者(表3)。同样地,说明金斑喙凤蝶雄蝶翅展在3个亚种间存在连续而微小的变异。然而,由于差异微小,辨识度低,不同作者在亚种分类处理上并不一致,如T.i.himalaicus与T.i.imperialis的亚种区分主要在于雌虫形态差异为“后翅背面为灰色,盘区颜色常较imperialis浅,向后更趋于黄色,盘区黑色半月斑更窄,第5和6翅脉间有少量黑色鳞粉(Soibam,2016)”,但Monastyrskii (2007)认为两者差异甚微,应归为同物异名(表2)。事实上,亚种分类时,低辨识度的形态描述常难以避免,但其不确定性显然会影响后续应用,甚至产生争议。比如关于金带喙凤蝶亚种T.i.imperatrix,“Bryk(1934)将该亚种视为himalaicus,但即使一些雌虫个体也较himalaicus更接近bhumipholi(Pen,1937),以及关于亚种T.i.gerritesi“来自泰国东北的新亚种更近似而不是来自泰国北部的bhumipholi与来自老挝北部的gillesi(Nakano,1995)等。

表3 金斑喙凤蝶部分亚种分类的形态比对原文描述Table 3 Morphological descriptions and comparisons between subspecies in Teinopalpus aureus
3 珍稀蝶种的亚种分类与保护意义
与蝴蝶广布种(generalist)相比,蝴蝶特有种或专性种(specialist)在各种干扰(人类活动、生境丧失、气候变化)作用下表现更脆弱(如对气候变化反应慢等)(Warren,2001),因而常作为珍稀种类列入生物多样性保护名录,如IUCN红色名录(IUCN Standards and Petitions Committee,2019),以引起更多关注。然而,在生境丧失、破碎化影响下,以及物种自身对特定生境要求下,蝴蝶特有种的地理种群常被周边不适生境包围,发生范围明显受限。因而,其种群分布无论在区域或地方尺度,都更类似于“岛状”结构,易满足亚种分类的地理隔离原则,尤其是喙凤蝶属、虎凤蝶属Luehdorfia等山地蝶类(曾菊平等,2012;苏杰等,2019),所以种下分类时易出现“过度亚种化”。如中华虎凤蝶陕南亚种L.chinensisshoui与李氏亚种L.chinensisleei(姚肖永等,2008;苏杰等,2019)和本次的金带喙凤蝶亚种T.i.imperatrix与T.i.bhumipholi等可归为“过度亚种化”问题。因为,在结合考虑生态位等性状后,发现中华虎凤蝶陕南亚种L.chinensisshoui与李氏亚种L.chinensisleei(苏杰等,2019)、金带喙凤蝶亚种T.i.imperatrix与T.i.bhumipholi实际上处于相同的植被群落与生态区,生态位相似,地理隔离并不显著,所以不建议独立为亚种。
珍稀蝶类亚种分类中,应遵从表型差异原则。然而,因野外个体罕见等原因,珍稀蝶类的表型差异比对时容易面临标本量偏少问题。例如,喙凤蝶属亚种分类的副模都在4个以内,包括3个仅1个副模的情况(Hope,1843;Turlin,1991;Collard,2007)。标本量少,一方面使得亚种分类75%原则方法无法应用;另一方面,可能出现单性(雄或雌性)副模,即形态描述有性别缺失。信息缺失显然将影响到后续的亚种分类与应用,例如,喙凤蝶亚种分类正模为雄蝶或雌蝶,但副模均为雄蝶,形态描述出现雌性缺失。所以,在喙凤蝶后续亚种分类中,由于雌蝶分类信息缺失,一些亚种形态差异只能在雄性间比对(尤其是金斑喙凤蝶亚种,见表3),这样虽能获得分类性状信息,但不完整,可信度低,增加了应用中的不确定性。例如,喙凤蝶两姊妹种在同一行政区(如中国四川、泰国清迈等)出现多个亚种名,就由不确定性引起。
在珍稀蝴蝶保护中,“过度亚种化”或亚种分类信息的不确定性,都可能影响到保护方案的有效实施。事实上,昆虫保护工作常受属、种鉴定困难影响,亚种分类信息的不确定性则会进一步增加管理的难度。所以,昆虫多样性保护管理者常被建议选择更确定的分类信息(如种或属级分类单元)来设计、规划保护方案,而避免将种下分类单元(如亚种)列入保护名录(袁德成,2001)。这一方面是本着“最有利于动物保护”的预防原则(袁德成,2001);另一方面,也利于正确预算物种保护的投入成本(Agapowetal.,2004),以及做好优先保护区、优先保护类群等的筛选或取舍。
为此,本次参考地理隔离原则与表型差异原则,并根据分类优先律,提出对珍稀喙凤蝶属亚种进行适当修订,如:将亚种T.i.bhumipholi作为T.i.imperatrix的新异名,将亚种T.a.laotiana作为T.a.eminens的新异名,而将T.a.wuyiensis,T.a.guangxiensis与T.a.nagaoi均作为指名亚种T.a.aureus的新异名(这与地理亲缘关系分析结果一致)(黄超斌,2016)等,可为该珍稀蝴蝶保护管理工作提供更清晰的亚种分类信息,并为后续有关喙凤蝶的保护遗传学研究、以及生境保护网络建设等提供重要参考。
4 小结与展望
亚种是《国际动物命名法规》认可的最低等分类阶元。达尔文认为具有更多物种的动物属也应包含有更多的品种(包含现代亚种概念)(Darwin,1859),并进一步推测后者可能在长期进化动力学与物种未来进化中发挥重要作用。而事实上,亚种是形成新种的必由过渡阶段,因为地理隔离是构成新种的前提,而亚种可能由于进一步的地理隔离导致生殖隔离而发展成为新的物种。相对于特有种或专性种而言,广布种更易形成亚种,其广布特征使得地理种群间更倾向于隔离,处于不同的生态环境(如气候类型不同),单独演化成支系,完成亚种形成过程,并进而进入特化阶段,走向新种形成阶段。因此,广布种也被认为是特有种进化过程的过渡阶段(Wahlberg,2001),对于蝴蝶类群而言,前者的数量明显少于后者(Dennisetal.,2011)。
值得注意的是,一个物种的各地理种群间都可能存在微小差异,因而并非都必须冠以亚种名,不然易陷入“过度亚种化”,导致亚种分类应用中名称混乱。对于珍稀蝶类而言,“岛状”种群分布格局虽看似满足亚种分类的地理隔离原则,但可能是由于长期的生境丧失或破碎化的结果,亚种分类需结合比对其他性状(如生态位分化等),以降低“过度亚种化”可能,减少不确定性,提高亚种信息在物种保护、管理与规划中的有效利用(Zink,2004)。表型差异也是亚种分类的原则,但珍稀蝶类可能因标本数量偏少,而出现亚种确立时分类信息不完整的情况(如单性描述、单性副模等)。这种情况需提起注意,在更多的确定信息未采集之前,不建议进行亚种分类。当然,地理种群间倘若符合地理隔离与表型差异原则,尤其在分类学上遵从75%法则,或具备遗传分化证据,就应赋予新的亚种名。因为,亚种作为生物多样性的重要组成部分,具有一定独特性(O′Brien and Mayr,1991),也属于自然进化过程的一部分。
致谢感谢Helmholtz Centre for Environmental Research (UFZ)的Josef Settele教授与Senckenberg Deutsches Entomologisches Institute的Thomas Schmitt教授在蝴蝶文献查阅时提供帮助。
