eEF1A2敲除对小鼠骨骼肌组成的影响*
2022-08-03宋子岱李长龙杜小燕陈振文
宋子岱 郭 萌 裴 朗 王 妍 李长龙 杜小燕 陈振文
(首都医科大学基础医学院,北京 100069)
骨骼肌是人体最大器官,约占人体体重的40%[1],其功能主要包括收缩、产热、代谢和内分泌等。骨骼肌含量和结构组成异常,可导致机体运动障碍、累及器官受损,甚至危及生命[2]。骨骼肌纤维分为快肌纤维和慢肌纤维,快肌纤维以肌球蛋白重链4(myosin heavy chain 4,Myh4)基因编码的2B型纤维为主,其纤维直径更粗、收缩力量大且以糖酵解代谢为主;慢肌纤维以肌球蛋白重链7(myosin heavy chain 7,Myh7)基因编码1型纤维为主,其线粒体数量更多、毛细血管网更丰富、抗疲劳能力更强[3]。研究表明,衰老会引起快肌纤维向慢肌纤维的转换,致使老年人运动能力和活动范围下降,容易出现代谢异常并增加糖尿病、高血压等疾病风险,生活质量严重下降[4],但引起骨骼肌质量下降和纤维类型转换的分子机制目前仍不完全清楚。
真核细胞翻译延长因子1-α(eukaryotic translation elongation factor 1 alpha,eEF1A)作为G蛋白家族成员,是构成真核延长因子1的4个亚基之一[5]。eEF1A在哺乳动物中存在2种亚型,eEF1A1在哺乳动物组织广泛表达,而eEF1A2仅在心脏、骨骼肌以及脑中表达[6]。eEF1A依赖GTP催化氨基酰tRNA与核糖体A位点的结合,将氨基酰化的tRNA在蛋白翻译的延伸阶段募集到核糖体中,以此调控蛋白翻译的延伸阶段[7]。纯合Wasted小鼠(eEF1A2基因启动子和第一外显子缺失),表现出严重的神经退变表型,在出生23 d即死亡[8];临床研究也显示,eEF1A2突变会造成儿童脑发育滞后,肌张力低下和心肌病[9]。这些研究表明,eEF1A2在机体发育过程中发挥关键作用。前期研究发现,骨骼肌eEF1A2通过激活蛋白激酶Cβ和内质网应激,加剧了雄性糖尿病动物模型的胰岛素抵抗[10],这说明eEF1A2也参与了骨骼肌葡萄糖代谢过程。尽管eEF1A2在脑和心脏发育以及骨骼肌代谢中发挥重要作用,然而eEF1A2是否参与调控骨骼肌的结构和组成目前尚不清楚。
本研究利用12月龄eEF1A2基因全身敲除小鼠,检测其主要骨骼肌类型的脏器系数、骨骼肌形态以及快慢肌纤维标志物的表达变化,以探讨eEF1A2对小鼠骨骼肌组成和纤维类型的作用。
1 材料和方法
1.1 材料
1.1.1实验动物:Cre-ERT2小鼠由北京生命科学研究所王凤超博士赠送,eef1a2flox/flox(eef1a2fl/fl)小鼠委托北京唯尚立德生物科技有限公司利用CRISPER/Cas9技术制作。实验所需eef1a2fl/fl;CreERT2+小鼠(iHBKO)及其对照eef1a2fl/fl;CreERT2-小鼠(Control)饲养于首都医科大学SPF级动物房【SYXK(京)2018-0003】,自主饮食和饮水,温度(22±4)℃,湿度 55% ± 5%,室内光照明暗周期12 h/12 h。所有实验操作严格遵守动物伦理和动物福利的要求,按照首都医科大学实验动物管理委员会的相关规定进行实验,伦理委员会审批号:AEEI-2017-097。将12月龄的iHBKO小鼠(9只,2雄7雌)和对照Control小鼠(3只,2雄1雌)分别连续3 d腹腔注射30 mg/kg他莫昔芬,观察1周后进行相关实验。小鼠实施安死术后,分离双下肢腓肠肌、胫骨前肌、比目鱼肌以及趾长伸肌,分别放入4%多聚甲醛固定以及液氮冻存。
1.1.2主要试剂:他莫昔芬(Sigma);玉米油(Sigma);TRIzol(诺唯赞);5 × All-In-One RT Master Mix试剂盒(abm);EvaGreen 2 × qPCR MasterMix试剂盒(abm);BCA 蛋白质浓度测定试剂盒(康为世纪);苏木精染液(中杉金桥);伊红染液(中杉金桥)。
1.2 方法
1.2.1实时荧光定量PCR(qRT-PCR):利用TRIzol有机溶剂抽提法提取小鼠骨骼肌mRNA,使用5 × All-In-One RT Master Mix试剂盒将2 μg mRNA反转录为cDNA。qRT-PCR使用Bio-Rad CFX Connect系统和EvaGreen 2 × qPCR MasterMix进行。反应体系设置为cDNA模板1 μL,上游引物1 μL,下游引物1 μL,纯水7 μL以及EvaGreen 2 × qPCR MasterMix 10 μL。反应条件设置为95 ℃ 15 min,95 ℃ 10 s,60 ℃ 30 s,重复 40 个循环,熔解曲线设置为 60 ℃ 5 s。统计结果使用的公式为:mRNA 相对表达量= 2-△△Ct。以Gapdh作为内参基因,引物序列如表 1 ,均由北京天一辉远生物科技有限公司合成。
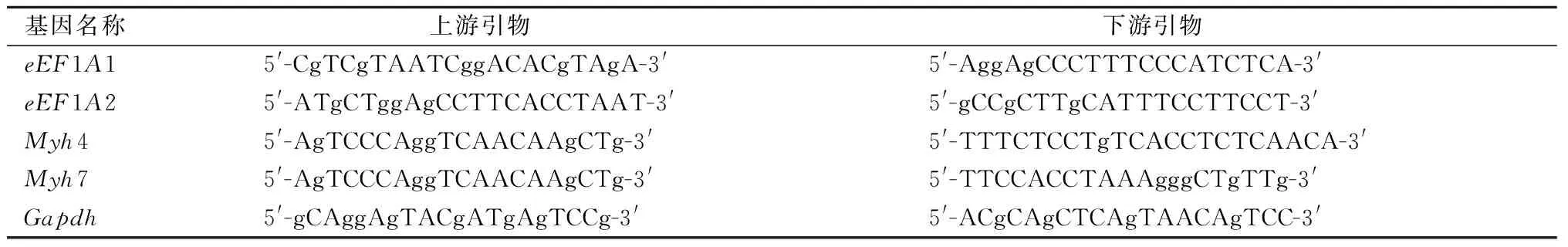
表1 qRT-PCR引物序列Table 1 The primer sequences for real-time quantitative PCR
1.2.2Western Blot:将小鼠骨骼肌放入RIPA∶PMSF=100∶1溶液中并利用匀浆机打碎,在15 000 r/min,15 min条件下获得蛋白上清液,利用BCA 蛋白质浓度测定试剂盒(Thermo Fisher Scientific)对蛋白上清液进行浓度测定并进行定量、变性。取20 μg 蛋白进行 SDS-AGE 凝胶电泳分离,电转蛋白至硝酸纤维素膜。硝酸纤维素膜在5% 脱脂牛奶中封闭1 h。eEF1A2抗体(1∶1 000 稀释,Proteintech)以及GAPDH抗体(1∶1 000 稀释,华安生物)4 ℃孵育过夜,TBST 洗涤后与二抗(HRP)室温孵育 1 h,TBST 洗膜后用化学发光法显色。利用凝胶图像分析系统(Bio-Rad)扫描蛋白质印迹条带灰度。
1.2.3HE染色:将小鼠骨骼肌于4%多聚甲醛中固定24 h后,进行常规组织脱水、石蜡包埋及病理切片的制备。进行染色时将石蜡组织切片按如下顺序进行操作:二甲苯(I) 5 min—二甲苯(Ⅱ) 5 min—二甲苯(Ⅲ) 5 min—100%乙醇(Ⅰ) 2 min—90%乙醇(Ⅱ) 2 min—80%乙醇(Ⅲ)2 min—70%乙醇(Ⅳ) 2 min—流水冲洗5 min—苏木精液染色5 min—流水稍洗去苏木精液1~3 s—19盐酸乙醇1~3 s—流水洗至返蓝—0.5%伊红液染色1~3 min—蒸馏水洗1~2 s—80%乙醇洗1~2 s—95%乙醇(Ⅰ) 2~3 s—100%乙醇(Ⅱ) 3~5 s—二甲苯(Ⅰ) 2 min—二甲苯(Ⅱ) 2 min—二甲苯(Ⅲ) 2 min,最后用中性树胶封固。
1.2.4统计学分析:利用GraphPad Prism完成统计分析,结果表示为平均值±SEM。符合正态分布的计量资料两组间的比较采用非配对的t检验,P<0.05被认为有统计学意义。
2 结果
2.1 他莫昔芬诱导的eEF1A2全身敲除小鼠骨骼肌eEF1A2敲除效率验证
qRT-PCR显示,在快肌腓肠肌(gastrocnemius,GA)(图1A,P<0.01)和趾长伸肌(extensor digitorum longus,EDL)(图1B,P<0.000 1),慢肌比目鱼肌(soleus,SOL)(图1C,P<0.000 1)和胫骨前肌(tibialis anterior,TA)(图1D,P<0.001)中,eEF1A2表达水平与对照组相比均极显著降低;同时,氨基酸序列与eEF1A2高度同源的eEF1A1,其mRNA表达水平在实验组和对照组差异无显著性(图1A—D)。Western Blot显示iHBKO小鼠腓肠肌(GA)(图2A)、趾长伸肌(EDL)(图2B)、比目鱼肌(SOL)(图2C)以及胫骨前肌(TA)(图2D)中eEF1A2蛋白水平均极显著下降(图3,P< 0.001)。以上结果说明,他莫昔芬诱导的iHBKO小鼠在骨骼肌组织中实现了eEF1A2的高效敲除。
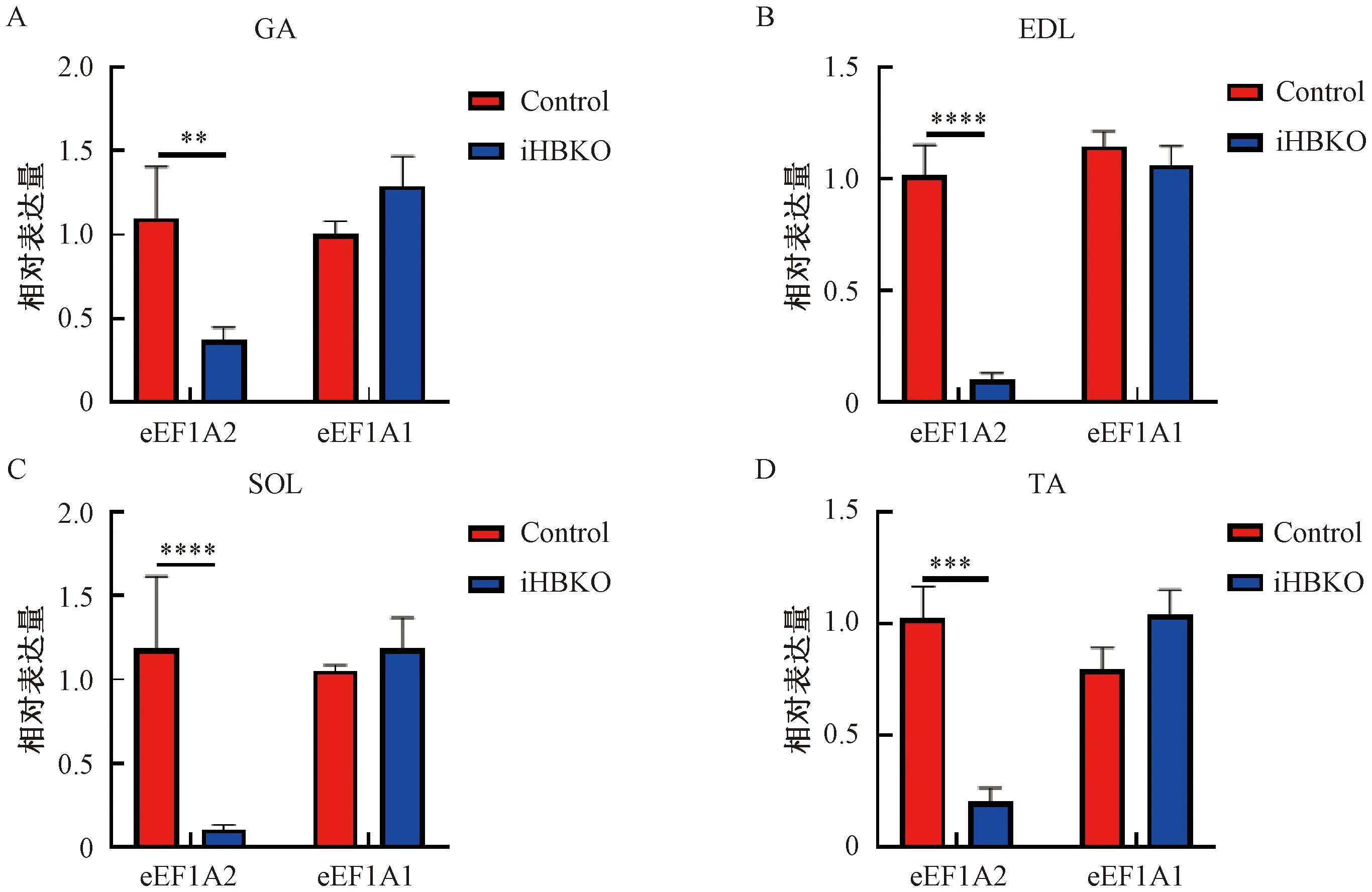
图1 iHBKO小鼠骨骼肌eEF1A2及eEF1A1转录水平检测注:利用qRT-PCR对iHBKO小鼠A(腓肠肌,GA),B(趾长伸肌,EDL),C(比目鱼肌,SOL)以及D(胫骨前肌,TA)的eEF1A2以及eEF1A1转录水平进行检测;**、***和****分别表示P< 0.01、P< 0.001和P< 0.0001的统计学显著性Fig.1 Detection of eEF1A2 and eEF1A1 transcript levels in skeletal muscle of iHBKO miceNote:The eEF1A2 and eEF1A1 transcript levels of A (gastrocnemius,GA), B(extensor digitorum longus,EDL), C(soleus,SOL) andD( tibialis anterior,TA) muscle in iHBKO mice were detected by qRT-PCR. **,*** and **** denoted statistical significance at P< 0.01,P< 0.001 and P< 0.0001, respectively

图2 iHBKO小鼠骨骼肌eEF1A2蛋白水平检测注:利用Western Blot对iHBKO小鼠A(腓肠肌,GA),B(趾长伸肌,EDL),C(比目鱼肌,SOL)以及D(胫骨前肌,TA)的eEF1A2蛋白水平进行检测Fig.2 Detection of eEF1A2 protein levels in skeletal muscle of iHBKO miceNote:The eEF1A2 protein levels of A(gastrocnemius,GA), B(extensor digitorum longus,EDL), C(soleus,SOL) and D(tibialis anterior, TA) muscle in iHBKO mice were detected by Western Blot

图3 iHBKO小鼠骨骼肌eEF1A2 Western Blot统计结果注:对图2中Western Blot结果进行统计;***和****分别表示P< 0.001和P< 0.0001的统计学显著性Fig.3 Western blot analysis for eEF1A2 in skeletal muscle of iHBKO miceNote:Statistical analysis of Western blot results in Figure 2; *** and **** denoted statistical significance at P< 0.001 and P<0.0001, respectively
2.2 eEF1A2敲除导致小鼠腓肠肌含量下降
基因敲除小鼠几种主要骨骼肌的脏器系数结果显示,iHBKO小鼠腓肠肌脏器系数与对照相比显著下降(图4,P<0.05),而两种慢肌(胫骨前肌与比目鱼肌)脏器系数与对照组相比也有下降的趋势。对eEF1A2基因敲除小鼠骨骼肌组织形态学分析,HE染色显示iHBKO小鼠腓肠肌与对照组相比未出现萎缩现象(图5);且腓肠肌横截面积在敲除小鼠(812.0±40.72 μm2)和对照小鼠(902.5±55.83 μm2)中无差异(图6)。分析认为,eEF1A2敲除小鼠腓肠肌含量减少。
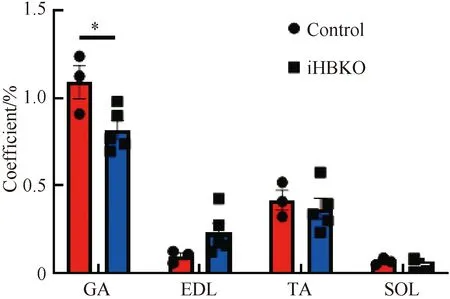
图4 iHBKO小鼠骨骼肌脏器系数统计注:分别记录iHBKO小鼠4种骨骼肌重量,并分别除以小鼠体重,得到4种骨骼肌脏器系数;*表示P<0.05的统计显著性Fig.4 Organ coefficients of skeletal muscle of iHBKO miceNote:Four skeletal muscle weight/body weight in iHBKO mice were recorded;* denoted statistical significance at P<0.05
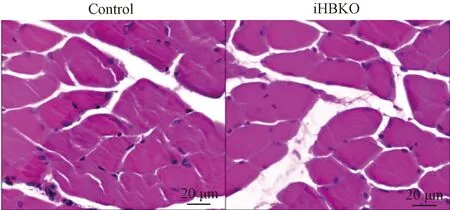
图5 iHBKO小鼠腓肠肌HE染色结果注:对iHBKO小鼠腓肠肌切片进行HE染色,观察腓肠肌形态及横截面积Fig.5 HE staining of gastrocnemius muscle of iHBKO miceNote:HE staining was performed on gastrocnemius muscle sections from iHBKO mice to detect the gastrocnemius muscle morphology and cross-sectional areas
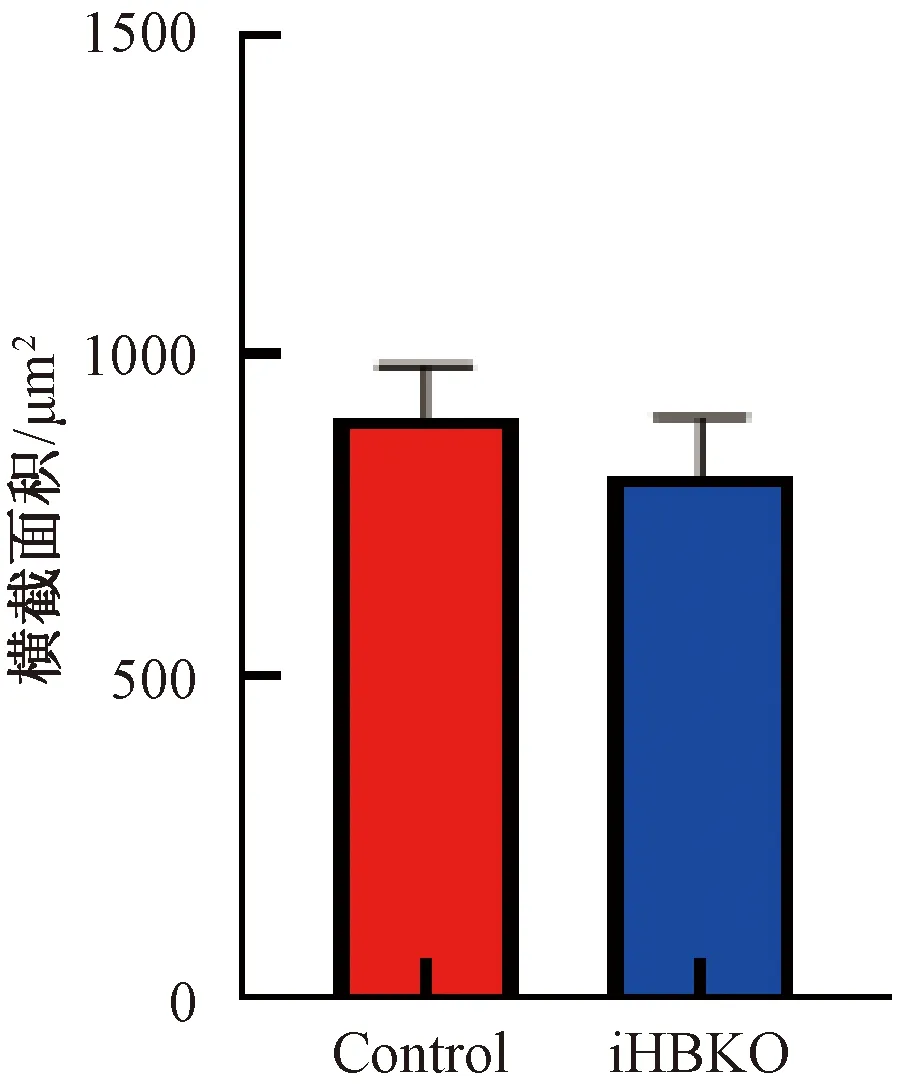
图6 iHBKO小鼠腓肠肌横截面积统计注:利用Image J软件统计图5中腓肠肌横截面积Fig.6 Analysis of iHBKO mice gastrocnemius cross areasNote:Use Image J to analyze the gastrocnemius muscle cross-sectional areas in Figure 5
2.3 eEF1A2敲除导致小鼠快慢肌纤维组成改变
利用qRT-PCR分别检测4种代表性骨骼肌组织中快肌纤维标志物Myh4和慢肌纤维标志物Myh7的表达变化。结果显示,eEF1A2基因敲除小鼠中,在快肌腓肠肌(GA)(图7A,P<0.001)和趾长伸肌(EDL)(图7B,P< 0.000 1)中Myh7表达明显升高,而Myh4则明显降低,但在慢肌比目鱼肌(SOL)(图7C,P<0.01)和胫骨前肌(TA)(图7D,P<0.001)的Myh4和Myh7表达均显著降低。以上结果提示,eEF1A2基因敲除引起小鼠快肌中的快肌纤维向慢肌纤维的转化,并导致慢肌中的两种肌纤维的减少。

图7 快慢肌标志物在iHBKO小鼠骨骼肌中的表达变化注:利用qRT-PCR对iHBKO小鼠A(腓肠肌,GA),B(趾长伸肌,EDL),C(比目鱼肌,SOL)以及D(胫骨前肌,TA)的Myh4以及Myh7转录水平进行检测;**、***和****分别表示P<0.01、P<0.001和P<0.0001的统计学显著性Fig.7 The expression of fast and slow muscle markers in iHBKO miceNote:qRT-PCR was used to detect Myh4 and Myh7 transcript levels in A(gastrocnemius,GA), B(extensor digitorum longus,EDL),C(soleus,SOL), and D(tibialis anterior,TA) muscles in iHBKO mice; **,*** and **** denoted statistical significance at P<0.01,P<0.001 and P<0.0001, respectively
3 讨论
骨骼肌支撑着人体的正常生理活动,同时也是人体最大的产热、代谢和内分泌器官。维持骨骼肌的正常组成和形态是机体活动的必要因素。骨骼肌易受以基因组不稳定性为特点的衰老的影响,这同时也是引起骨骼肌质量下降的最重要原因[11]。在人体老化过程中,骨骼肌肌球蛋白重链2型的合成率下降,导致肌纤维数量的减少[4]。从机制上讲,老龄人体对胰岛素和氨基酸的敏感性降低,二者对哺乳动物雷帕霉素靶蛋白复合物1(mammalian/mechanistic target of rapamycin complex 1,mTORC1)的激活作用降低,导致蛋白合成减少,进而降低肌肉质量[12]。而作为蛋白质翻译延伸因子,有研究表明随线虫衰老,eEF1A2蛋白的丰度和活性逐渐降低[13],但也有研究认为在线虫衰老时eEF1A的表达并不改变[14],因此在衰老过程中eEF1A的表达模式和功能尚无明确定论。本研究中,eEF1A2基因敲除导致了腓肠肌脏器系数的明显下降,这表明eEF1A2在维持腓肠肌含量发挥重要作用,这可能是通过其蛋白翻译延长效率实现的。由此可见,eEF1A2对骨骼肌质量有一定影响。
骨骼肌中快肌与慢肌的比例不是一成不变的,通过耐力、爆发力等训练,快肌和慢肌纤维比例可以互换[15]。在脊髓损伤、长时间卧床、微重力环境等疾病状态下,慢肌纤维会向快肌纤维转化,而在Ⅰ型糖尿病、癌症、衰老等情况快肌纤维会向慢肌纤维转换[15]。这些非生理性的纤维类型改变往往对人体产生严重的影响。如在慢性阻塞性肺病中,膈肌的快肌纤维向慢肌纤维转化,会使患者呼吸负担加重[16];在多种恶性肿瘤患者体内,也出现快肌纤维向慢肌纤维的转换,并影响患者生存期[15]。因此,维持不同肌纤维在一定比例十分重要。本研究发现,eEF1A2的缺失同时造成了快肌以及慢肌中Myh4基因的表达降低,而Myh4编码2B型肌纤维以糖酵解代谢为主,属于快肌纤维。由此推测,eEF1A2可能主要参与快肌纤维的合成和维持,敲除eEF1A2后导致了快肌纤维的减少;同时由于主要肌纤维的减少,为维持骨骼肌的功能,使得快肌中的慢肌纤维代偿性的增加,即编码1型纤维的Myh7的表达增加。综上,eEF1A2的缺失可能与衰老过程中快肌纤维向慢肌纤维的转变有关。
本研究通过建立eEF1A2基因敲除小鼠模型发现,eEF1A2的缺失对骨骼肌的形态无明显影响,但降低了小鼠腓肠肌的脏器系数,同时导致小鼠快肌纤维向慢肌纤维转化。研究结果为进一步探究eEF1A2对小鼠骨骼肌组成和纤维类型的作用和机制研究提供了动物模型,也为衰老过程中骨骼肌的变化研究提供数据参考。
