缺氧环境下子宫内膜腺上皮细胞中微RNA表达谱及细胞凋亡变化
2022-07-30王含必窦帅杰刘思邈张婉玉刘美芝邓成艳
王含必,窦帅杰,刘思邈,张婉玉,刘美芝,邓成艳
中国医学科学院北京协和医院 1妇科内分泌与生殖中心 国家妇产疾病临床研究中心 2疑难重症及罕见病国家重点实验室,北京 100730 3北京致成生物医学科技有限公司,北京 100176
成功妊娠需具备两个必备条件:优质的胚胎和正常的子宫内膜容受性,即除对胚胎质量有严格要求外,在允许胚胎植入的窗口期内,还需子宫内膜保持良好的容受性[1-2]。子宫内膜修复障碍导致的薄型子宫内膜(排卵前子宫内膜厚度<7 mm)可降低子宫内膜的容受性,进而影响妊娠[3-4]。薄型子宫内膜的具体发病机制尚未明确,目前研究认为子宫血供不足可能是影响子宫内膜生长的重要因素之一[5],各种原因引起的子宫血流量降低将导致子宫内膜处于缺氧环境,诱发子宫内膜腺上皮细胞(endometrial glandular epithelial cell,EEC)能量代谢障碍乃至缺氧损伤,最终影响子宫内膜的容受性。
子宫内膜容受性的调控涉及子宫内膜一系列复杂的动态变化,适宜水平的EEC凋亡对于子宫内膜容受性的建立至关重要,凋亡异常将不利于子宫内膜保持良好的容受性[6]。p53是调节细胞DNA修复的重要蛋白,在缺氧、辐射、毒物等导致的基因损伤无法修复时,p53则进入线粒体并活化促凋亡基因,促进凋亡小体复合物形成,启动凋亡。微RNA(micro-RNA,miRNA)是一类具有20~25个核苷酸的非编码RNA[7],其通过与信使RNA互补结合,可影响靶蛋白表达,在细胞增殖、分化、凋亡和血管生成等多方面发挥调控效应[8]。缺氧环境可诱导多种细胞释放大量miRNA[9],且缺氧环境可诱发细胞的代谢活动进行重编程,增强凋亡信号传导[10]。目前尚不明确缺氧环境是否可引起EEC中miRNA表达谱异常,进而影响p53蛋白表达,促进EEC凋亡。本研究探究缺氧环境对EEC中miRNA表达谱的影响及miRNA调控细胞凋亡的机制,为薄型子宫内膜引起子宫内膜容受性降低的治疗提供理论依据。
1 材料与方法
1.1 主要试剂与仪器
本研究主要试剂包括EEC原代细胞及配套培养基(武汉普诺赛公司)、胎牛血清(美国Gibco公司)、0.25% 胰蛋白酶、Trizol RNA提取试剂盒(美国Invitrogen 公司)、兔抗p53抗体(一抗,美国Cell Signaling Technology 公司);HRP-山羊抗鼠IgG抗体(通用型,二抗)、HRP-山羊抗兔IgG抗体(通用型,二抗)、DAB显色剂(丹麦DAKO公司)、Annexin V-FITC细胞凋亡检测试剂盒(上海碧云天生物技术有限公司)、反转录酶试剂(5×Reverse Transcriptase M-MLV)、缓冲液Reverse Transcriptase M-MLV、RNA酶抑制剂(日本TaKaRa公司)、2×Mltra Master Mix、二硫苏糖醇(dithiothreitol,DTT)、不含RNA酶的水和dNTP(中国康为世纪有限公司)、引物(上海生工有限公司)。主要设备包括BX19-HK830倒置相差显微镜(深圳市奥斯微光学仪器有限公司)、Ni-U正置荧光显微镜(日本Nikon公司)、7500 Fast Real-Time PCR System荧光定量PCR仪(美国Applied Biosystems公司)、VeritiProTM梯度PCR仪、Micro17R高速冷冻离心机(美国Thermo Fisher公司)、SC-2546实验室台式低速离心机(安徽中科中佳科学仪器有限公司)以及CLINX ChemiScope6100化学发光成像系统(上海勤翔科技有限公司)。
1.2 方法
1.2.1 EEC培养与分组
取EEC原代细胞培养于配套的专用生长培养基,该培养基含胎牛血清、生长添加剂、青霉素、链霉素,O2∶CO2体积比为95∶5。待EEC密度增长至80%~90%,采用0.25%胰蛋白酶消化,进行传代培养。收集对数生长期EEC,接种至六孔板并随机分为缺氧组和对照组(每组1×105个细胞/孔,每组3个复孔)。缺氧组、对照组分别置于缺氧环境(O2∶N2∶CO2体积比为1∶94∶5)[11]和正常氧环境(O2∶CO2体积比为95∶5)中培养,4 h后进行后续实验。
1.2.2 miRNA表达谱改变
收集两组EEC至EP管中(2×106个细胞/管),加入1 mL Trizol提取EEC中的总RNA。采用高通量测序法进行RNA测序,并绘制miRNA表达谱聚类热图。
1.2.3 miR-7974、miR-7704基因表达检测
将提取的RNA反转录为cDNA。采用实时荧光定量PCR(realtime fluorescence quantitative polymerase chain reaction,RT-PCR)法检测EEC中miR-7974、miR-7704基因表达水平。反应体系:cDNA 1 μL,10 μmol/L规格的上、下游引物各1 μL(表1),Master Mix 10 μL和不含RNA酶的水8 μL。反应条件:95 ℃ 10 min,95 ℃ 15 s,60 ℃ 1 min,95 ℃ 15 s,40个循环;60 ℃ 1 min,95 ℃ 15 s。以sno-RNA202为内参,采用2-△△Ct法计算miR-7974、miR-7704基因相对表达量。
1.2.4 细胞凋亡检测
采用流式细胞术检测EEC凋亡情况。收集两组细胞置于离心机中,常温下,离心半径16 cm,800 r/min离心10 min弃上清液;采用PBS 重悬沉淀,制成单细胞悬液并分装至EP 管中。采用Annexin V-FITC细胞凋亡检测试剂盒标记相应抗体(每组各设1个空白对照),室温避光孵育25 min后置于离心机中。常温下,离心半径16 cm,3000 r/min 离心5 min;加入1 mL PBS去除多余抗体后加入 200 μL多聚甲醛固定细胞,置于流式细胞仪中上机检测,结果以流式细胞图表示。
1.2.5 miR-7704的靶蛋白p53及凋亡相关蛋白检测
通过检索TargetScan、miRDB、miRWalk 3个生物信息学网站,发现miR-7974的靶基因可能为p53(图1A)。本研究通过碱基互补配对,发现miR-7974可与p53的3’非翻译区(3’-untranslated region,3’UTR)连续9个碱基相配对(图1B),因此,推测p53为miR-7974的靶基因。半胱天冬酶3(caspase3)是细胞凋亡过程中重要的执行者,其直接参与可促进凋亡小体形成的细胞内结构分解[12],B细胞淋巴瘤-2(B cell lymphoma-2,Bcl-2)是重要的抗凋亡蛋白,caspase3水平增加、Bcl-2水平降低将导致细胞凋亡增多和DNA损伤[13]。
本研究采用蛋白印迹法(Western blot)测定miR-7704靶蛋白(p53)和凋亡过程中的重要蛋白表达水平。收集两组细胞,4 ℃下加入细胞裂解液(1∶1000),静置30 min后10 000×g离心25 min收集上清液,采用BCA法测定蛋白浓度。取30 μg 蛋白样品,经10% SDS-PAGE凝胶电泳后,将蛋白印记至PVDF膜,并采用含5% 脱脂奶粉的封闭液室温封闭1 h;加入兔单抗p53(Abcam ab26)、兔单抗Cleaved-caspase3(caspase3的活化形式,Abcam ab32042)兔单抗Bcl-2一抗(Abcam ab182858)及鼠单抗β-Tubulin,4 ℃孵育过夜,采用TBST缓冲液 洗膜3次;加入山羊抗兔、山羊抗鼠二抗,室温孵育 1 h,采用 TBST缓冲液洗膜 3 次。采用ChemiScope6100化学发光成像系统观察蛋白表达情况。
1.3 统计学处理
采用 GraphPad Prism 7.00软件绘制图表及统计学分析。miRNA表达谱、miR-7704和miR-7974相对表达量等计数资料的组间比较采用t检验。以P<0.05为差异具有统计学意义。
2 结果
2.1 缺氧引起miRNA表达谱变化
采用高通量测序法对miR-215-5p、miR-7851-3p、miR-369-5p等21种常见miRNA表达水平进行检测。结果显示与对照组比较,缺氧组EEC中16种miRNA表达上调,5种miRNA表达下调(图2)。其中对照组表达丰度较高的miR-7704和miR-7974,在缺氧组表达下降最为显著,差异具有统计学意义(P均<0.05),提示缺氧环境可引起EEC中miRNA表达谱改变。
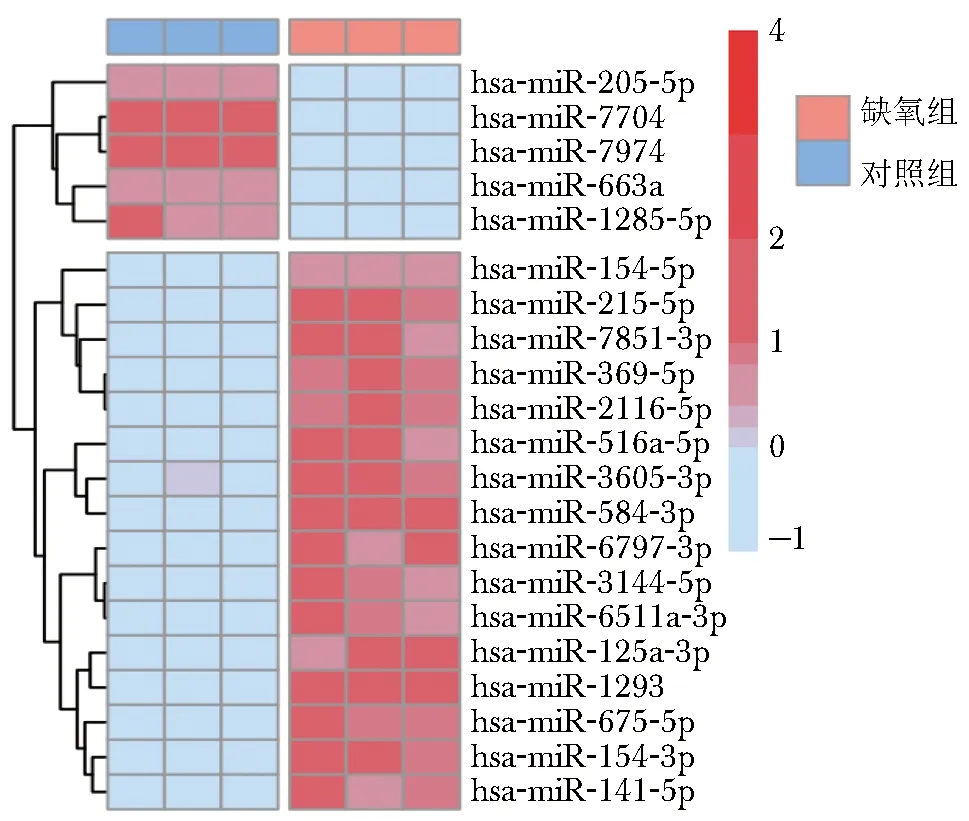
图2 缺氧组和对照组EEC中微RNA表达谱的聚类热图EEC:子宫内膜腺上皮细胞
2.2 缺氧引起EEC中miR- 7974表达显著下调
RT-PCR结果显示,与对照组比较,缺氧组EEC中miR-7704和miR-7974相对表达量分别降低20%、80%(图3),提示miR-7974可能受缺氧环境的影响更大,对EEC生物学功能的影响更显著。
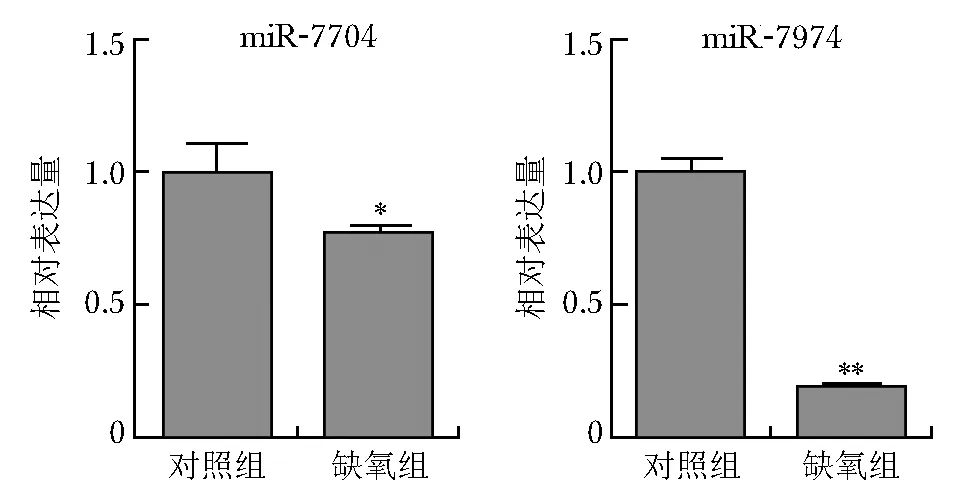
图3 缺氧组和对照组EEC中miR-7704和miR-7974相对表达量的柱状图EEC:同图2;与对照组比较,*P<0.05,**P<0.001
2.3 缺氧诱导EEC凋亡
对照组有少量早期凋亡的EEC,几乎无晚期凋亡细胞。与对照组比较,缺氧组早期凋亡率、晚期凋亡率均明显增高(P均<0.001,图4),提示缺氧环境可诱导EEC发生明显的凋亡。
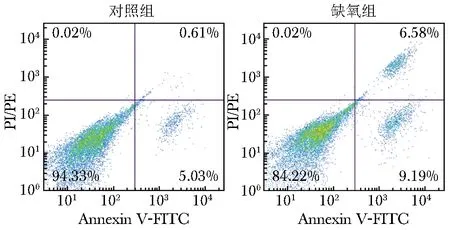
图4 缺氧组和对照组EEC凋亡的流式细胞图EEC:同图2
2.4 缺氧环境诱导miR-7974的靶蛋白p53表达上升
Western blot检测结果显示,与对照组比较,缺氧组EEC中p53蛋白表达升高,伴抗凋亡蛋白Bcl-2表达降低(P均<0.05),Cleaved-caspase3无显著变化(P>0.05),提示缺氧诱发EEC中miR-7974表达下调,可能通过上调其靶蛋白p53水平进而诱导细胞凋亡(图5)。
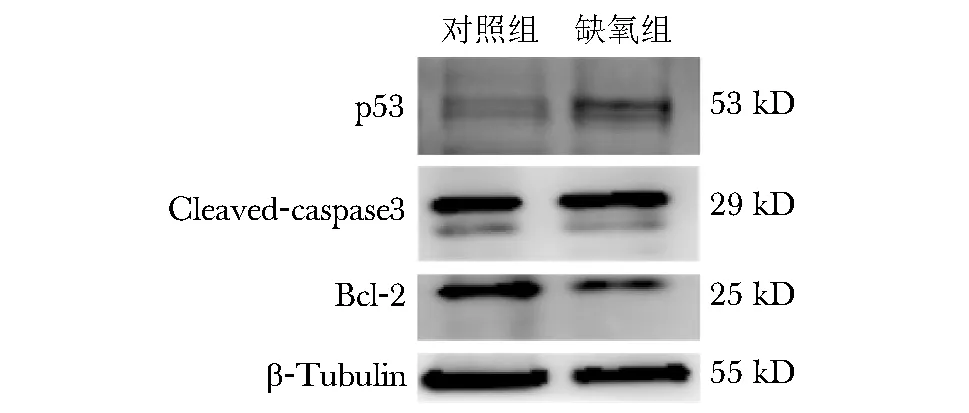
图5 缺氧组和对照组凝胶电泳图Bcl-2:B细胞淋巴瘤-2
3 讨论
子宫内膜良好的容受性与胚胎着床密切相关,EEC是子宫内膜的主要构成成分,EEC损伤将直接影响子宫内膜的功能状态[4]。多种因素可损伤EEC,包括孕期人工流产等妊娠状态的宫腔操作、子宫内膜病变的反复刮宫术、宫腔感染(如结核杆菌感染)等,损伤的病理机制与外界因素引起子宫血管损伤,子宫内膜缺血、缺氧导致的EEC凋亡增多有关。本研究从分子水平探索了EEC的损伤机制,结果显示缺氧环境可引起miRNA表达谱改变,其中以miR-7974对缺氧最为敏感。缺氧导致的miR-7974显著下调可促进其靶蛋白p53表达,继而启动细胞凋亡,导致EEC损伤。
暴露于缺氧环境的生物体首先可通过一系列自身调节,产生多种适应性信号,以促进细胞存活,但持续性缺氧可活化基因调控网络,改变细胞的行为和功能,发生细胞凋亡,最终导致不可逆损伤[14]。缺氧环境下,EEC可发生氧化应激,产生过量的肿瘤坏死因子-α、白细胞介素-6、单核细胞趋化蛋白-1、环氧合酶2、前列腺素E2等炎症介质,诱导细胞损伤和凋亡,但促进凋亡的具体机制尚未阐明[15]。miRNA通过调控靶基因、靶蛋白表达,影响细胞增殖、凋亡等生物学行为,是生命活动的重要调节因子。其调控机制:miRNA以碱基互补的原则与靶基因信使RNA的3’UTR相结合,进而阻断信使RNA翻译或使其降解,从而实现对靶蛋白的调控。目前已明确,miRNA参与肿瘤、心脑血管疾病、糖尿病等疾病进程的调控,是临床研究的热点。缺氧环境是否影响EEC中miRNA表达及其病理机制尚未明确。
本研究团队在缺氧条件下对EEC进行培养,以期诱导细胞损伤,模仿薄型子宫内膜的病理过程。首先通过高通量测序法对EEC中miRNA表达情况进行检测,结果显示21种常见miRNA中,缺氧环境下16种表达上调,5种表达下调,提示缺氧环境可诱导EEC中miRNA表达谱发生改变。进一步对正常氧环境时EEC高表达,缺氧环境中表达降低最为显著的miR-7704和miR-7974进行了RT-PCR检测,结果发现相较于对照组,缺氧组miR-7704和miR-7974相对表达量分别降低20%、80%,提示miR-7974对缺氧更为敏感,可能对EEC生物学功能的影响更显著。
本研究缺氧组EEC早期凋亡率、晚期凋亡率均明显升高,表明细胞凋亡增多可能是导致子宫内膜损伤的机制之一。Western blot检测结果显示,缺氧的EEC中p53表达上调,抗凋亡蛋白Bcl-2表达降低,进一步提示其呈现高凋亡状态。p53基因是重要的抑癌基因,通过凋亡途径促进损伤的细胞死亡,是其修复DNA损伤、发挥抑癌功能的重要方面。正常生理情况下,p53 蛋白以低水平无活性形式存在于机体细胞中,当发生DNA严重破坏或细胞暴露于其他不利环境时,p53蛋白的半衰期明显延长[16],并进入线粒体激活Bcl-2相关X蛋白(Bcl-2 association X protein,Bax)、凋亡蛋白酶激活因子1、Noxa等促凋亡蛋白,抑制Bcl-2家族(Bcl-2、Bcl-X、Beclin-1)等抗凋亡蛋白表达[12],上调凋亡水平。Bcl-2是研究较多的抗凋亡蛋白,通过阻断线粒体细胞色素C释放并与Bax、Bcl-X相互作用抑制细胞凋亡[17-18]。Bcl-2表达降低可间接促进细胞色素C释放、凋亡酶形成和caspase3活化,诱导细胞内结构分解,启动凋亡[19]。因此,推测缺氧环境诱导的miR-7974表达降低可活化p53,引起EEC大量凋亡,引发EEC损伤,最终导致子宫内膜损伤和子宫容受性降低。
本研究局限性:(1)有研究将Bcl-2与Bax的比值作为凋亡的评价指标,判断细胞是否发生凋亡、评判凋亡的趋势[20-21]。本研究仅通过实验发现p53可能为miR-7974的靶蛋白,未评价细胞凋亡趋势。后续实验中可考虑将Bcl-2与Bax比值作为细胞凋亡的评价标准,进一步分析miR-7974对细胞凋亡的调控能力。(2)缺氧仅为薄型子宫内膜细胞内膜形成的因素之一,机械损伤、频繁刮宫等破坏子宫内膜的血供和营养供给,亦在其中发挥重要作用。本研究仅构建缺氧环境,探究其对薄型子宫内膜的影响,未能全面模拟薄型子宫内膜发生的机制。(3)此实验为体外培养,尚缺乏动物模型和临床试验验证。
综上,缺氧环境可诱导EEC中miRNA表达谱改变,其中以miR-7974表达下调为最显著。miR-7974水平降低可促进其靶蛋白p53表达,诱发EEC凋亡。本研究从基因水平揭示了缺氧引起子宫内膜损伤的可能机制,为相关疾病的治疗提供了新的研究线索。
作者贡献:王含必负责研究设计、数据分析、论文撰写;窦帅杰负责实验实施、数据分析、论文撰写;刘思邈、张婉玉、刘美芝负责实验实施;邓成艳为项目总负责人,指导研究设计、论文写作及修改。
利益冲突:所有作者均声明不存在利益冲突
