不同质地葡萄果实果胶含量及基因表达差异分析
2022-07-30吕春晶孙凌俊
马 丽, 乔 军, 吕春晶, 孙凌俊
(1.辽宁省果树科学研究所,辽宁营口 115009; 2.辽宁农业职业技术学院,辽宁营口 115009)
葡萄属于葡萄科(Vitaceae)葡萄属()多年生植物,是世界上最古老的物种之一,其果实可用于鲜食、酿酒和制干,具有很高的经济价值。葡萄在世界果树生产中占有非常重要的位置,据联合国粮食及农业组织(FAO)统计,2018年全世界葡萄栽培面积约为715.8万hm,总产量约为7 912.6万t。我国葡萄栽培面积达到79.8万hm,约占全球总面积的11.1%,产量1 349.5万t,约占全球葡萄总产量的17.1%(http://www.fao.org/faostat/en/#data),是世界葡萄的第一生产大国。就全世界葡萄生产而言,近70%的葡萄用于加工,超30%的葡萄用于鲜食,而我国则以鲜食为主,占据70%以上,鲜食葡萄生产是我国葡萄产业的主体。
葡萄属于典型的非呼吸跃变型果实,果实软化发生时间比呼吸跃变型果实要早,且由于其为浆果,果实皮薄,汁多,在食用、运输贮存过程中易发生破裂、腐烂等问题,所以就其鲜食品质而言,果实质地已经成为判断鲜食葡萄果实新鲜程度的一个重要指标。不同葡萄品种间果实质地存在较大差异,脆肉型品种一直备受消费者关注,现已成为重要的育种材料及目标品种,然而造成不同品种间质地差异的机制还不是很清楚。因此,了解葡萄果实成熟软化及不同质地形成因素,对于葡萄综合品质改良及葡萄栽培有着非常重要的理论意义。
1 材料与方法
1.1 试验材料
试验选用脆肉型葡萄品种红地球(L.)和软肉型葡萄品种玫瑰香(L.)为试材,2个品种均定植于沈阳农业大学葡萄试验园内(41°50′24″N,123°24′41″E),管理水平一致。鉴于2个葡萄品种在辽宁省沈阳地区物候期接近,2018年2个品种分别于盛花后56 d(转色前)开始采样,每隔7 d采样1次,直到果实成熟。采样时间均定在08:00进行,采集方式:每个品种随机选取长势较为一致的6株树,在植株的不同位置选取20个果粒作为1个重复,每个时期采集3个重复,用装有冰袋的保温箱迅速带回实验室。在实验室内,每个重复样品随机选取5个果粒,将种子剔除,果实磨成细末状,用锡纸包好,经液氮冷冻1 h后置于-80 ℃超低温冰箱保存备用。其他果实放入4 ℃冰箱用于果实质地及内含物测定。
1.2 试验方法
1.2.1 果实质地相关指标测定 葡萄果实质地测定采用整果穿刺测试法。穿刺试验使用的是英国Stable Micro Systems 公司生产的TA.XT Plus质构仪,穿刺试验选择2 mm直径的P/2针状探头,测试在室温下进行,测试时将被测试的浆果放置在质构仪载物台的中心位置,用手稍稍固定,力量以不压迫果实为宜,样品被穿刺部位为葡萄浆果中部偏上区域。质构仪主要参数设定为测前速度为 1 mm/s,贯穿速度为1 mm/s,测后速度为10 mm/s,穿刺深度设为7 mm,负载触发力为5 g。参数设定时需要注意,穿刺既要保证一定的穿刺深度,又要避免在穿刺过程中探头接触到葡萄种子,影响测试结果。质地相关指标包括:果皮硬度(PPH)、果皮破裂距离(PB)、果肉硬度(SF),果皮硬度是从探头接触果皮开始,到探头穿刺果皮瞬间所用的力(g);果皮破裂距离是从接触到刺破果皮瞬间探头下行位移(mm);果肉硬度是探头穿刺果肉到测试结束所测感应力的平均值(g)。
1.2.2 总糖和总酸含量测定 葡萄果实中总糖和总酸含量利用高效液相色谱仪Agilent1260(美国安捷伦公司)进行测定。吸取挤压好并混和均匀的葡萄汁,加入超纯水定容至10 mL,吸取定容后果汁 2 mL 置于离心管中,12 000 r/min离心10 min。然后吸取上清液和pH值=2.6的磷酸二氢钾盐溶液各500 μL至新的2 mL离心管中进行总酸的提取,加入等体积的乙腈进行总糖的提取,后使用水系微孔滤膜进行过滤,过滤后的溶液用于上机检测。
总糖含量测定使用的是Agilent Zorbax Carbohydrate色谱柱(4.6×250 mm,5 μm)和示差折光(RID)检测器,柱温为40 ℃,运行时间为15 min;流动相体积比为75 ∶25的乙腈和纯净水,流速为 1.0 mL/min,进样量为10 μL。
总酸含量测定使用的是Agilent SB-AQ色谱柱(4.6×250 mm,5 μm),G1314B可变波长紫外检测器(VWD),参数为柱温25 ℃,波长210 nm,运行时间15 min。流动相为体积比为95 ∶5的10 mmol/L磷酸二氢钾和甲醇溶液,流速0.8 mL/min,进样量为10 μL。
1.2.3 果胶含量测定 原果胶和可溶性果胶采用比色法测定,略有改动。具体方法如下:称取 1.0 g 带果皮的果肉组织,在液氮保护下研成匀浆,转到事先装好25 mL的95%乙醇溶液的50 mL离心管中,沸水加热30 min,在煮沸的过程中要及时补加95%乙醇溶液,冷却至室温后,需8 000 r/min离心 15 min,去除上清液,再加入95%乙醇溶液,沸水浴加热,如此反复3次。然后将沉淀置于20 mL蒸馏水中,50 ℃保温30 min后,溶解果胶。取出冷却至室温,8 000 r/min 离心15 min,再将上清液用蒸馏水定容至100 mL,该液为水溶性果胶;在沉淀中加入25 mL 0.5 mol/L的硫酸溶液,沸水加热1 h,取出冷却至室温,于8 000 r/min离心15 min,上清液用蒸馏水定容至100 mL,该液为原果胶。测定时吸取1 mL提取液,加入6 mL浓硫酸,沸水加热 20 min,冷却室温后,加入0.2 mL 1.5 g/L咔唑-乙醇溶液,暗处放置30 min后,测定反应液在530 nm处的吸光度,重复3次。根据溶液吸光度,在标准曲线上查出相应的半乳糖质量,计算果胶含量,以生成的半乳糖醛酸的质量百分数表示果胶含量。
1.2.4 荧光定量PCR分析 将2个品种花后56、63、70、84、98 d的果实样品进行总RNA提取,反转录合成后的cDNA稀释5倍作为候选基因差异表达分析的模板。实时荧光定量反应在ABI QuantStudio 6 Flex System(Applied Biosystems,Foster City,CA,美国)384 孔仪器上进行,使用SYBR® PremixExⅡ(Takara,Madison,WI,美国)制造反应混合液。以作为内参基因,使用2-ΔΔ计算差异表达值,所有样品进行3次生物学重复,RT-PCR引物采用Primer3设计,相关引物信息见表1。

表1 相关基因RT-PCR引物
2 结果与分析
2.1 果实发育过程中果实质地差异变化
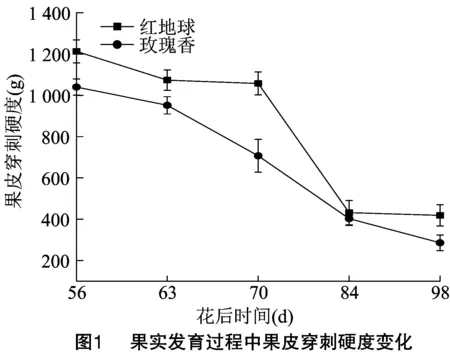
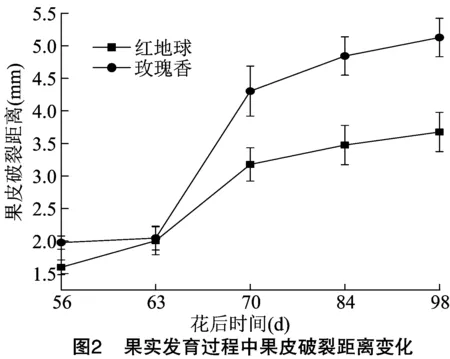
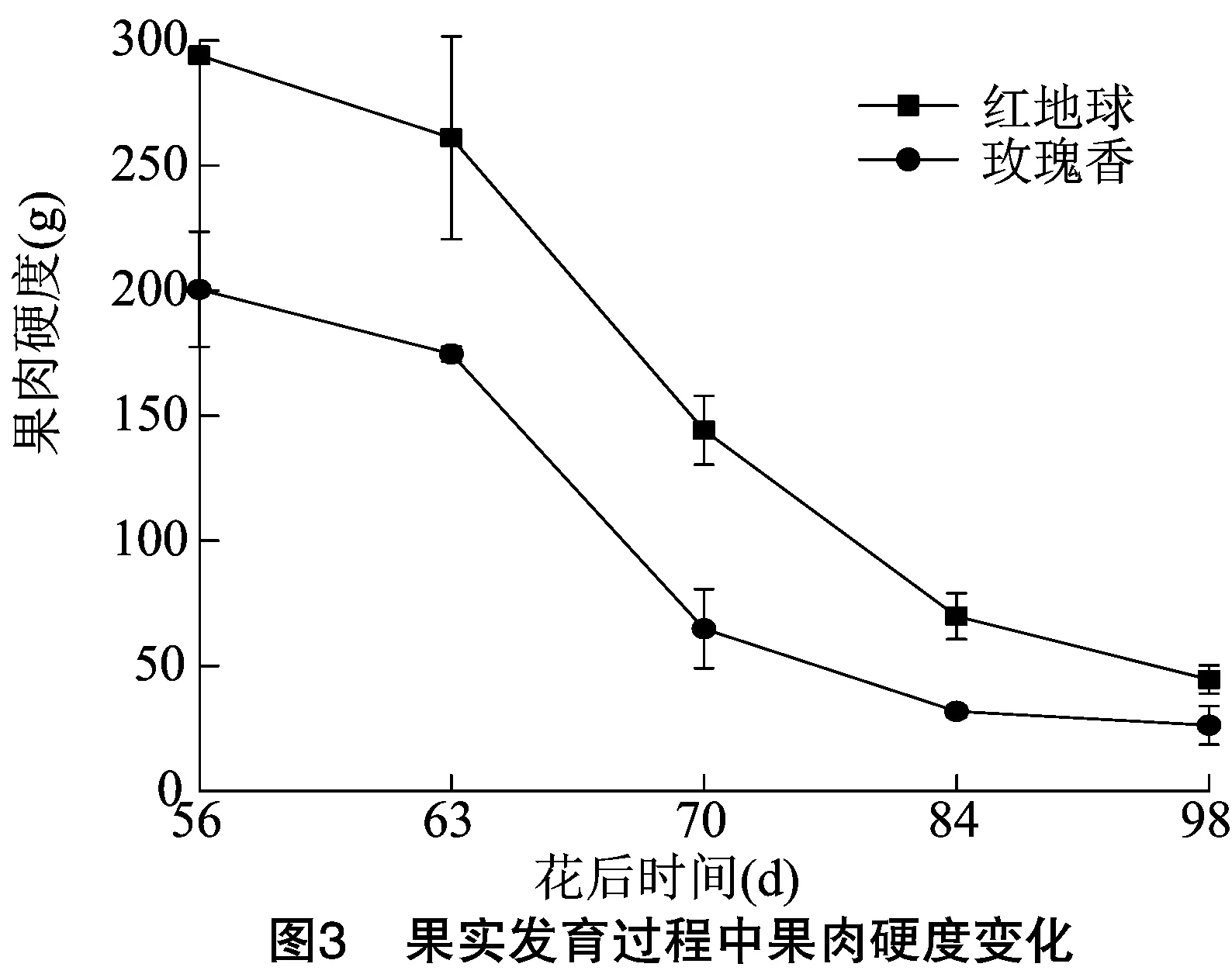
葡萄果实质地是评价果实成熟软化的一项重要依据,对红地球和玫瑰香2个品种各发育阶段果实质地进行测定,由图1分析发现,红地球和玫瑰香2个品种果皮硬度在转色期(花后 63 d)之前均保持在较高水平,随着发育时期递进,软肉玫瑰香在转色期后,果皮硬度迅速下降,直到成熟期达到最低值,而脆肉红地球果皮硬度下降时期较玫瑰香晚,在花后70 d才开始下降。虽然在花后84 d时,2个品种果皮硬度接近,但成熟时红地球果皮硬度还是远高于玫瑰香的果皮硬度。果皮破裂距离可以反映果皮被刺穿时浆果形变的程度。由图2可知,2个品种果皮破裂距离呈相似的增长趋势,转色前期,2个品种果皮破裂距离都处于较低水平,转色期红地球和玫瑰香果皮破裂距离比较接近,分别为2.00、2.05 mm,而后随着果实成熟,果皮破裂距离迅速增大,成熟期相差较大,分别为3.68、5.13 mm,整个发育过程红地球的果皮破裂距离都明显低于玫瑰香的果皮破裂距离。由图3可知,果肉硬度更多反映的是果肉的机械性能,果实整个发育过程中,2个品种果肉硬度变化趋势与果皮硬度的变化趋势相似,即随着果实的生长发育果肉硬度逐渐降低,转色后下降速度迅速增快,近成熟期果肉硬度变化较为平缓,成熟期时红地球和玫瑰香果肉硬度分别降到了44.6、26.28 g,并且在每个发育阶段红地球的果肉硬度都高于玫瑰香的果肉硬度。
2.2 果实发育过程中果实糖酸含量变化
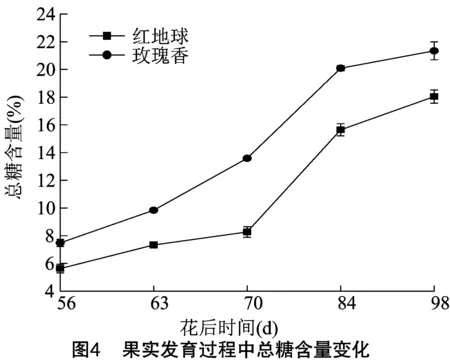
伴随着果实的发育、成熟软化,与成熟软化相关的内含物也发生了明显的变化,总糖含量增加,总酸含量降低。由图4可知,转色前期葡萄果实内总糖含量较低,随着果实转色,迅速增加,在果实成熟时达到最高,而后基本保持不变。红地球在转色后1周总糖含量增加较慢,出现了一个平台期,而后迅速增加,成熟期达到最高。玫瑰香总糖含量增加比红地球快,且玫瑰香各个时期的总糖含量均明显高于红地球。由图5可知,果实内总酸含量随着果实的成熟逐渐降低,红地球是个退酸较快的葡萄品种,转色时总酸含量急剧下降,而后变化较为平缓,但总酸水平也逐渐降低,直到成熟。玫瑰香果实内的总酸含量也随着果实成熟软化逐渐下降,但各时期含量都高于红地球。
2.3 果实发育过程中果胶含量变化
随着葡萄果实的成熟软化,果实细胞壁发生降解,果实质地发生了显著变化,果胶作为细胞壁基质中含量最高的一类大分子物质,其组分和含量也发生了变化。由图6可知,随着葡萄果实的生长发育,2个品种原果胶含量逐渐下降,但在整个发育阶段,红地球果实内的原果胶含量一直高于玫瑰香。随着果实的成熟软化,原果胶在相关酶的作用下分解成可溶性果胶含量,可溶性果胶呈逐渐增加趋势,且在转色后增加速度较快,与原果胶相反,红地球果实内的可溶性果胶含量一直低于玫瑰香(图7)。

2.4 果实发育过程中果胶降解酶基因的表达
由图8可知,-表达量在软肉玫瑰香和脆肉红地球中均呈先升高后降低的趋势。在转色前期,2个品种的表达量接近,转色期二者表达量均迅速增加,此后-在2个品种中的表达模式发生变化。脆肉红地球中-的表达量在转色期达到最高值后逐渐下降,在成熟期达到最低值,而软肉玫瑰香中的表达量在转色后继续升高,花后 84 d 略有降低,但都处于较高水平,而在成熟期表达量才下降到较低水平。在葡萄果实转色后的各个时期,软肉玫瑰香中-的表达量一直极显著高于脆肉红地球中的表达量(除花后98 d)。结果表明,-基因可能在转色后期调控了葡萄不同质地形成。

表达量在软肉葡萄玫瑰香转色期即迅速升高,而后持续较高水平,在花后84 d开始下降,而在脆肉葡萄红地球中的表达量在果实发育前期一直处于较低水平,花后70 d才开始逐渐升高,花后84 d达到最高值后又略有下降(图9)。比较2个品种各个时期的表达量,发现在转色前后表达量差异极显著,随着果实成熟软化表达量差异变小,但仍然是软肉葡萄玫瑰香中的表达量明显高于脆肉葡萄红地球中的表达量。结果说明可能对葡萄成熟软化及不同质地形成起到一定作用。

表达量在2个品种中的变化趋势明显不同,在整个发育阶段,软肉葡萄玫瑰香中的表达量一直处于较高水平,而在脆肉葡萄红地球中表达量呈现出先升高后降低再升高的趋势,且各个时期,在软肉葡萄玫瑰香中的表达量一直极显著高于在脆肉葡萄红地球中的表达量(图10)。结果表明,很可能调控了葡萄果实的成熟软化及质地的形成。
3 结论与讨论
3.1 果胶变化与不同质地形成的关系
果胶是果实细胞壁中非常重要的大分子物质,其组分和含量对果实质地都会产生一定的影响。Dong等以2个欧洲冬梨品种为试材,对贮存5个月后可溶性果胶(WSP)含量和果胶降解酶活性进行测定,得出欧洲梨果实质地与PG、PL、PME、-GAL、-ARF等果胶降解酶高度相关,WSP含量越高,果实硬度越低,认为果实中WSP可以通过减少细胞黏附,使果实更容易软化。Yang等发现沉默基因的番茄果实中可溶性果胶含量降低,果实硬度增加,认为可溶性果胶含量与番茄果实硬度密切相关。管雪强等对红地球葡萄进行喷钙处理,结果发现外源钙可以增加果实内原果胶含量、总果胶含量以及果实硬度,且发现果实硬度与果胶含量呈极显著正相关。本研究发现脆肉葡萄红地球中原果胶含量在各时期都显著高于软肉葡萄玫瑰香中的含量,且果皮硬度和果肉硬度与原果胶含量呈极显著正相关,这与管雪强等的研究结果相似,与可溶性果胶含量呈极显著负相关,与Dong等的结论相似。
3.2 果胶降解酶基因与葡萄不同质地形成的关系
和基因可以降解去甲酯化的果胶,参与果实成熟软化过程。Villarreal等研究发现,可编码PG酶活性的基因在脆肉草莓中低表达,而在软肉草莓中高表达,而-(缺少PG 5′端 85 bp 而导致编码无活性的PG酶)基因表达模式正好相反,在脆肉草莓中高表达,在软肉草莓中低表达。Fabi等研究得出了基因在木瓜果实成熟软化中的关键作用。Atkinson等发现基因可以通过影响细胞壁降解和降低细胞间黏稠度来调控苹果果实硬度。也有研究得出不同PG活性造成了葡萄果实质地的差异。基因被认为是调节果实成熟软化的一种重要的候选基因。有研究者认为草莓果实成熟软化与基因有着直接的关系。Nunan等发现基因高表达可以促进葡萄果实软化。本研究发现不同质地葡萄果实的、基因表达模式相近,随着果实成熟表达量增加,但比较二者表达量发现软肉葡萄玫瑰香中的表达量高于脆肉葡萄红地球中的表达量,这可能是造成不同质地果实形成的关键因素,该结论与前人的研究结果相似。
Smith等研究得出-Gal在草莓果实成熟软化过程中起到了关键作用。Paniagua等研究发现,基因在草莓成熟过程中,参与了共价果胶的溶解过程,降低了果实硬度,得出的表达与草莓果实硬度密切相关。本研究发现在葡萄果实成熟软化早期,软肉型玫瑰香中-的表达量迅速升高,且在整个成熟过程中,软肉型玫瑰香-的表达量远远高于脆肉红地球中的表达量,结果表明-对葡萄果实质地差异形成起到一定作用,与前人的研究结果相似。
