高产高效水稻冠层光能利用特征研究进展
2022-07-28肖治林顾汉柱张耗
肖治林 顾汉柱 张耗
(扬州大学/江苏省作物遗传生理重点实验室/江苏省作物栽培生理重点实验室/江苏省粮食作物现代产业技术协同创新中心,江苏 扬州 225009;第一作者:1825754758@qq.com;*Corresponding author: haozhang@yzu.edu.cn)
水稻是我国的主要粮食作物。不断提高水稻产量仍然是一种刚性需求,关乎我国乃至世界粮食安全和人民生活水平。我国水稻单产从1950 年的2.1 t/hm2增加至2020 年的7.0 t/hm2,在世界主要产稻国中名列前茅[1-2],但同时也形成了水肥的高投入、产量的高回报、环境的高污染、低经济效益的生产模式,这给社会、经济和环境带来了巨大压力。近年来的研究表明,水稻产量与水肥资源利用效率能够实现协同提高[3-4]。光是绿色植物光合作用的能源,同时光还可以为氮代谢提供能量[5]。光能利用率(Radiation Use Efficiency,RUE)是指在一定时期内植物光合作用积累的同化物所含能量与同期地面辐射的能量之比,是水稻高产高效的决定因素之一[6]。作物进行光合作用的场所是冠层,因此冠层结构就影响着光在冠层中的分布,是决定作物群体光能高效利用与光合氮素利用效率的重要因素[7]。一般来说,在高光照条件下冠层上部光辐射能力强,叶片处于“光饱和”,光合效率高;下部叶片光辐射能力弱,叶片处于“光饥饿”,光合效率低[8]。如何构建健康的水稻冠层,提高光能利用率,进而提高冠层光合同化能力,对实现水稻高产高效具有重要意义。本文对高产高效水稻冠层光能利用特征进行了探讨并对未来的重点研究方向进行展望。
1 实现水稻高产高效的途径
水稻高产高效是指根据品种的特点和生产条件,在现有栽培技术的基础上,充分利用当地的温度、光照、水、肥、气,使生产潜力得到充分发挥,产量和效率达到更高水平[9]。以下3 点是实现水稻高产高效的主要途径:1)品种改良。通过品种改良能有效改善水稻品种的产量和资源利用潜力。国际上一般认为,当大田作物产量达到作物产量潜力的80%,进一步改善作物生长环境的潜力变小,产量增加的代价增加,难度增大[10]。因此,不断提升水稻产量潜力是实现高产高效的基础和关键。2)满足高产水稻“供需”匹配,实现资源利用高效。以往研究和生产注重于水肥的大量投入,而忽视了对作物生长实际需求的精确调控。稻田水肥管理必须结合土壤、水肥和环境统筹考虑[11-13]。3)优化群体冠层结构,增加花后物质生产是实现水稻高产高效的关键。塑造良好的群体冠层结构主要包括适宜叶面积、合理冠层光分布以及高效的光合作用,三者相互协调从而实现高产高效[14-16]。
2 高产高效水稻的叶面积特征
叶面积通常用叶长和叶宽进行估算,由叶片长和宽引起的冠层结构的不同会引起冠层光截获量与RUE的差异,进而影响水稻的生长发育与产量形成[17-18],增大叶面积可有效增加冠层光截获量,促进光合产物向经济器官的分配。研究表明,功能叶的光合产物约占水稻经济产量的 70%~80%[19]。SAITOH 等[20]研究表明,大、宽、长剑叶被认为是水稻高产的理想选择,总干物质积累量与冠层上层、中层和总光合有效辐射(Photosynthetically Active Radiation,PAR)截获率均呈显著正相关。马超[21]研究表明,粳稻高产需要叶面积具有以下特征:剑叶 59.52~65.58 cm2、倒 2 叶 45.84~49.88 cm2、倒 3叶36.23-40.44 cm2。姜元华等[22]认为,籼粳杂交稻(甬优 11、甬优12、甬优 13、甬优 15 和甬优 17)高产需要上3 叶的叶长分别为:剑叶61.19 cm、倒2 叶67.26 cm、倒3 叶 64.55 cm,叶宽分别为:剑叶 2.71 cm、倒 2叶 2.24 cm、倒 3 叶 1.96 cm。
如果说叶面积是从二维空间反映叶片性状,那么叶面积指数(Leaf Area Index, LAI)则是从多维空间来反映,是联系个体与群体的重要参数。LAI 是指单位土地面积植物叶片总面积的倍数。一般来说,在一定范围内,LAI 值越大产量越高[23-25]。叶面积指数对群体光合能力有显著影响,在生长后期保持较高的叶面积指数有利于增加群体冠层对光能辐射的截获。关于调节LAI,近年来有诸多报道。研究表明,LAI 随栽植密度的增加而增加[26-27]。潘广良[28]研究发现,湿润灌溉条件下,LAI 增长速度较快,有利于水稻的生长发育。因此,高产高效叶片结构的构建应当根据水稻品种特性和气候环境来制定有效的栽培技术措施,生育期内保持较大的叶面积,促进LAI 增大,增加PAR 截获量,从而获得高效的RUE。
3 高产高效水稻的冠层光能利用特征
3.1 高效光分布
在生产过程中,叶角、叶倾角、消光系数和叶片分布均会对高效的光分布产生影响,进而影响水稻冠层RUE。
叶角是影响光在水稻冠层分布的主要因素之一。较小的叶角可使作物叶片均匀用光,减小叶片间的相互遮蔽,从而增加RUE,优化冠层结构,获得高产。研究表明,叶角与其临界遮蔽角成反比,上层叶片叶角减小有利于增加冠层中、下部的群体透光率,但叶角增大时,合理的群体密度将会减少,这就使植株无法获得高效的RUE[29]。袁隆平院士[30]曾提出,3 张功能叶的叶角分别为 5°(剑叶)、10°(倒 2 叶)和 15°(倒 3 叶)有利于超高产。在实际生产中,因叶片、叶角等因素均会使叶片相互交错遮蔽,而叶倾角(Leaf Inclination Angle, FIA)的存在则为更多的叶面积捕捉到光辐射提供了可能[31]。研究表明,FIA 对有效叶面积有极显著的影响[32]。上层叶片叶倾角较小,有利于增加剑叶长度,从而改善冠层结构,获得群体光合速率与RUE 的协同提高[33-34]。上层叶大部分倾角为60°~90°是叶片群体最理想的状态。但是,不同品种适宜的叶倾角也有所差异。
消光系数(K 值)是反映叶角和叶面积的垂直分布状况和光辐射在冠层垂直递减状况的一个指标[35]。K值在一定程度上反映了该群落的结构及受光势态,且K 值受多种因素的支配。不同的冠层高度其K 值差异较大,中上层最高,单位叶面积截获的光最多,随着平均叶倾角的增大,K 值逐渐减小[36]。高产群体冠层中下部K 值较大、上部较小,有利于光向着冠层深处透射,增加RUE[37]。刘建丰等[34]研究发现,超高产杂交稻组合培矮64S×9311 单株日产量为 0.339 g/d,其 K 值为0.46;湖南农业大学水稻科学研究所选育的苗头组合98073S×E32 单株日产量为 0.396 g/d,其 K 值为 0.44;大面积推广的三系杂交稻组合汕优63 单株日产量为0.277 g/d,其K 值为0.52,即较小的K 值有利于高产。
叶片分布(Leaf Distribution, LD)以叶片在各方位角内的分布频率表示[38]。水稻冠层光的分布状况会随着叶片分布状况的变化而变化,相继引起冠层光截获量和叶片的受光状况发生改变,直接影响水稻干物质积累量和产量的形成。叶片分布极差较小是冠层光辐射特征合理的充分必要条件。有研究表明,LD 值随生育进程推进和施氮量的增加均呈现增加趋势,随方位角增大呈先增后减的趋势,以180°~270°最大[39]。在栽培过程中可通过适当的栽培措施来增大LD 值,增加叶片受光面积。
3.2 高效光合作用
高效的光合作用易受到叶片光合特征,叶片碳、氮代谢酶活性和光氮是否匹配的影响。
高产高效的叶片光合特征是指水稻等作物的叶片具备同化物的低消耗和同化物的高积累。在众多指标中,叶片净光合速率和光合持续期能最直观地反映养分积累情况。优化冠层结构、改善冠层叶片受光状况和延缓叶片衰老后,提高单位叶面积的光合速率是继续提高产量的有效方式[40]。徐云姬等[41]认为,花后高净光合速率及长稳定持续期是水稻高产的重要生理基础。并且在提高光合速率时,应当注意避免呼吸速率增加。光呼吸会消耗已同化CO2的碳素,故降低光呼吸也就成了水稻养分高积累的必要条件之一[42]。光呼吸强度髙,CO2补偿点也较高,光合作用强度则小,光合效能就较低;反之则光合效能较高。
叶片碳、氮代谢酶是限制水稻光合作用的酶类。报道最多的就是核酮糖-1,5-二磷酸羧化酶(RuBisCo)和硝酸还原酶。RuBisCo 活性的高低,是影响水稻光合作用综合能力的主要因素之一。在光合作用中,RuBisCo催化作用是卡尔文循环中第一个主要的固碳反应,它将大气中的游离二氧化碳转化为生物体中的能量储存分子。EVANS[43]认为,小麦旗叶RuBPCase(二磷酸核酮糖羧化酶)活性与叶片全氮量呈正相关。硝酸还原酶作为氮代谢过程中重要的调节酶和限速酶与作物氮素代谢密切相关。其催化还原硝酸为亚硝酸是植物中硝酸同化的限速步骤[44]。增加光强有利于提高植株的硝酸还原酶活性,使水稻植株获得较高的氮素同化能力,增强碳、氮代谢,而硝酸还原酶活性的提高则有利于水稻吸收氮肥,提高养分利用效率。
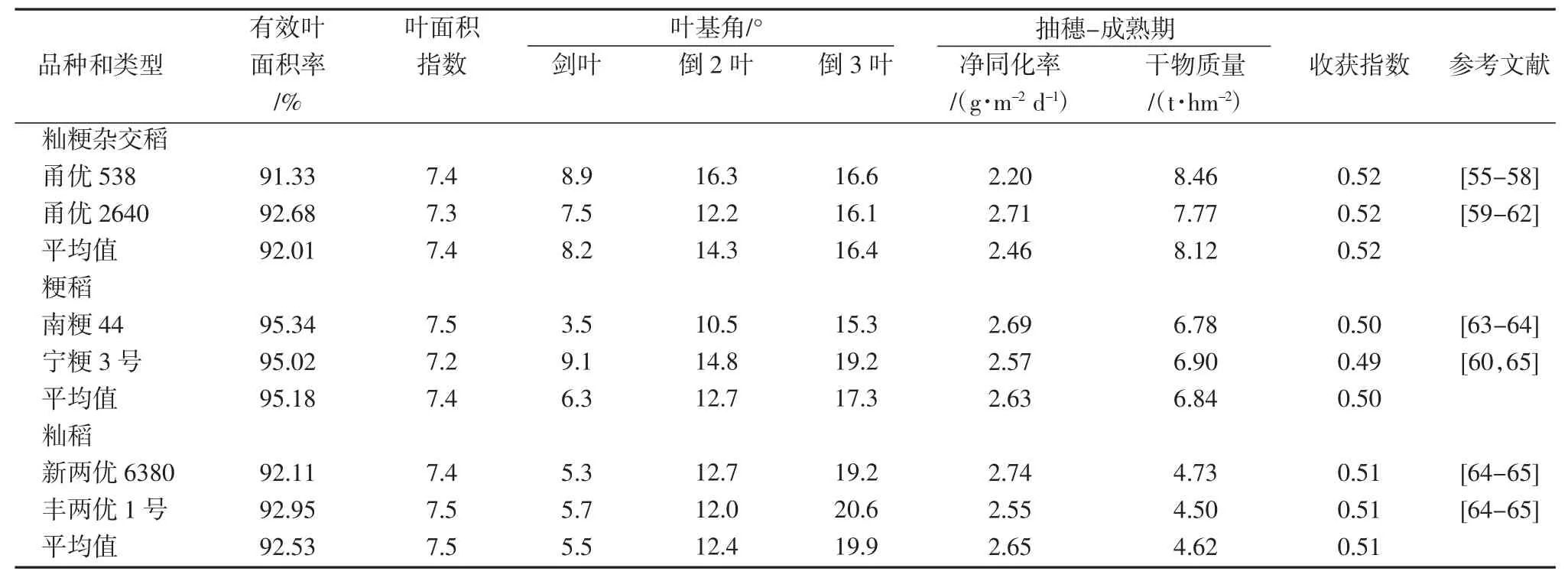
表1 不同品种和类型水稻的高产特征
光照和氮素的协同性直接影响植株体内的养分调控和碳氮代谢。在一定范围内,植株叶片的光合速率随着施氮量的增加而增加。氮素过多会造成碳、氮代谢失调,光合能力下降[45],而氮素过少,会使植株叶片内叶绿素含量降低,叶肉细胞光合活性降低,最终造成RUE下降[46]。光照对植株氮吸收的影响也很大,在一定范围内,光照强度与氮吸收量成正比关系[47]。在水稻生育期内,根据光照强度实时调节氮素供应量是获得氮素利用率与RUE 协同提高的先决条件。
4 高产高效水稻的物质积累与运转
水稻抽穗前拥有高效的叶片特征与冠层光能利用特征即高效的RUE,有利于促进干物质的高积累,而进入抽穗后,高效的RUE 与干物质的高积累共同促进非结构性碳水化合物(non-structural carbohydrate,NSC)的高效转化与运输。韦还和等[48]研究表明,水稻干物质最大积累速率出现在移栽后60~64 d。曹培培等[49]研究表明,水稻茎鞘干质量与茎鞘中NSC 呈显著正相关。以上研究结果表明,拔节孕穗期所形成较高的干物质积累为茎鞘中NSC 的高积累奠定了基础。
抽穗前,茎鞘作为临时的库,接收来自叶片合成的蔗糖。抽穗后,茎鞘由库转化为临时的源,向穗部以蔗糖的形式经过维管束再分配及卸载。在此过程中,蔗糖磷酸合成酶、蔗糖合成酶以及蔗糖转化酶的活性将会显著影响蔗糖的合成与积累,对水稻茎鞘中NSC 的转化与运输起到关键作用[50]。茎鞘NSC 对水稻产量的贡献可达30%,且其贡献率随着库容增加而增加[51]。多项研究表明,在生产中通过对氮肥和水分的正向调控,有利于高效的NSC 转化与运输[52-53]。茎鞘由库转变为源,产量的形成才刚刚开始。抽穗时,茎鞘中NSC 积累量较多;成熟时,茎鞘中残余量较少以及在此过程中相关酶活性较高等,都是高产形成的主要原因。收获指数是反映光合同化物转变为籽粒产量的一个重要指标,是指籽粒产量与干物质积累之比。有研究表明,“库大”和“源足”有利于同化物向库器官中分配比例增加,茎鞘中NSC 滞留量下降,收获指数上升[54]。
5 存在问题与研究展望
水稻冠层在吸收光能、贮藏养分的同时也在消耗养分。协调好水稻光能的高效吸收与养分的低消耗,对于当前水稻的高产高效具有重要意义。然而,目前对水稻高产高效调控机理的研究还不够深入。建议今后系统研究以下3个方面:1)系统研究高产高效水稻冠层光能利用的时空分布特征和水分养分吸收规律;2)从根系形态生理入手揭示水稻高效利用水分和养分的生理机制,特别是激素类的信号传导,对冠层结构和功能的调控研究;3)阐明栽培措施对水稻高产高效形成的调控机理。
