马氏珠母贝混养紫海胆对污损生物防除效果的研究
2022-07-28叶孝飞陆慧琳曾靖于宗赫洪喆聪余祥勇王梅芳
叶孝飞 陆慧琳 曾靖 于宗赫 洪喆聪 余祥勇 王梅芳
(华南农业大学海洋学院,广东广州 510642)
马氏珠母贝(Pinctadamartensi)又称合浦珠母贝,是我国重要的海水养殖贝类和珍珠生产的主要母贝。但是,在马氏珠母贝养殖过程中,其壳表和养殖网笼很快会被污损生物覆盖。污损生物是附着在船底、浮标和一切人工设施上的动、植物和微生物的总称。据估计,水产养殖行业每年因为污损生物而造成的直接经济损失占生产成本的5%~10%[1]。Ventilla[2]研究表明,缠绕盘踞在贝类壳表的污损生物会干扰贝壳的开闭。王春忠等[3]则观察到,贝壳上附着的污损生物在特定情况下会影响贝类的生长。Wallace等[4]发现,污损生物能与贝类竞争养殖区域的溶解氧。Su等[5]则进一步探讨了污损生物附着对马氏珠母贝摄食、耗氧和排泄物的影响。此外,污损生物的某些种类(如凿贝才女虫Polydoraciliata)还能凿穿贝类的贝壳,造成细菌感染,引发贝类病害[6]。使用化学药品处理养殖网具如涂抹防污材料三丁基锡(TBT)、有机铝、有机砷、铜化合物等,以及定期打捞养殖贝笼并用高压水枪冲洗是目前国内外海上筏式养殖普遍使用的方法[7],但是这些方法费时费力,不仅对贝类损伤较大[8],还会污染养殖海域环境。近些年,利用生物法[9],即通过食物链之间的关系选择污损生物的捕食者进行混养来控制污损生物的方法,因投入费用低、操作简单、对环境友好等特点而具有很大的发展潜力。
紫海胆(Anthocidariscrassispina)隶属于棘皮动物门、海胆纲、正形目、长海胆科、紫海胆属,在我国广东省沿海海域有广泛分布[10]。海胆食性杂,其主食藻类,也摄食纤毛虫、水螅虫、端足类、桡足类和有机碎屑,且具有很强的运动能力,几乎能到达养殖网笼全部区域,使污损生物难以附着[11]。利用生物防治法进行污损生物防治已经在实践中得到应用。Lodeiros等[12]利用紫海胆与牡蛎混养,与对照组相比,网笼污损生物附着量减少了74%,贝壳上的污损生物减少了71%。研究表明,南海相较于渤海、黄海更适于污损生物的生长[13]。但关于在南海用生物防治法的研究,如利用海胆去除贝类养殖过程中的污损生物,以及混养紫海胆对马氏珠母贝生长的影响等鲜有报道。基于紫海胆是南海海域常见生物,本试验选用紫海胆与马氏珠母贝在深圳大亚湾海域进行混养试验,通过在固定养殖密度的马氏珠母贝养殖笼中放置不同数量的紫海胆,以马氏珠母贝和紫海胆的生长指标、存活率和养殖笼上污损生物的质量为测量指标,比较不同混养密度对马氏珠母贝及紫海胆生长和存活的影响,以及对笼中污损生物的去除效果,以探究大亚湾海域马氏珠母贝更加合理、高效的养殖模式。
1 材料和方法
1.1 试验地点和材料
本试验在大亚湾自然海域(东经114.52°,北纬22.553°)马氏珠母贝养殖区进行。试验用养殖笼具为直径30 cm、高15 cm的6层珠母贝养殖笼,网笼孔目大小为10 mm×10 mm。养殖网笼每层之间用黑色塑料挡板避光,挡板厚3 mm,板上钻有直径为5 mm的小圆孔,孔间距10~15 mm。将养殖网笼悬挂在大亚湾内湾水深5 m以上的珠母贝筏式养殖区。
试验用马氏珠母贝和紫海胆均取自大亚湾海洋生物综合实验站。紫海胆初始壳径(4.00±0.12)cm,马氏珠母贝初始壳长(4.00±0.37)cm,均选择活力旺盛、健康状况良好的个体。
1.2 试验方法
设置4个试验组,即混养组(PA3、PA6、PA9)和对照组(P),每组在养殖马氏珠母贝30只/层的基础上,分别混养紫海胆3、6、9、0个/层。另外,设置3个紫海胆单养试验组(A3、A6、A9),养殖密度分别为3、6、9个/层。以上每个处理组设置5个平行。
马氏珠母贝和紫海胆的日常管理均参照“大亚湾马氏珠母贝海上养殖工艺”[14],按常规方法进行。试验时间为2020年10月—2021年2月。
1.3 数据处理
为了评估单养组与各混养组紫海胆和马氏珠母贝的生长状况,在试验结束时统计各组紫海胆及马氏珠母贝的存活数量、体质量、壳长,计算各组的存活率、特定生长率、壳长日增长率、肥满度。采用Ross等[15]的方法评估紫海胆对马氏珠母贝养殖网笼污损生物的去除效果。试验前,将养殖网笼完全浸没在养殖海水中12 h,沥干水分5 min后称量,即为干净网笼质量。将养殖区吊养的试验网笼捞上岸,用干净海水冲洗掉淤泥,在日光下晾晒2 h后分别称取质量,此质量减去干净网笼质量即为网笼污损生物的质量。计量贝体附着污损生物的方法为:用刀刮取各组的贝体污损生物后,用干净海水冲洗滤网去除淤泥,然后编号封装,-4 ℃冷冻,带回实验室进行分类鉴定并统计生物量。马氏珠母贝和紫海胆相关指标的计算公式如下。
存活率Rs(%)=(nt/n0)×100
(1)
特定生长率RSG(%/d)=100×(lnwt-lnw0)/t
(2)
壳长日平均增长率RLD(%/d)=100×(Lt-L0)/(L0·t)
(3)

(4)
式(1)~(3)中,nt为第t天时的存活个数,n0为初始存活个数,wt为第t天时的体质量(g),w0为初始体质量(g),Lt为第t天时的壳长(cm),L0为初始壳长(cm),t为试验时长(d)。
1.4 数据分析
采用SPSS 19.0软件对试验数据进行单因素方差分析(one-way ANOVA),设P<0.05为差异显著,采用Duncan’s法进行多重比较,用ORIGIN 2018软件制图。
2 结果
2.1 马氏珠母贝和紫海胆的生长情况
各组马氏珠母贝和紫海胆的生长指标见图1。结果显示,各混养组及P组间马氏珠母贝的存活率没有显著差异(P>0.05),混养组紫海胆的存活率均接近100%,各组间无显著差异(P>0.05)。PA6、PA9组马氏珠母贝的特定生长率显著大于P组(P<0.05),而PA3组与P组间差异不显著(P>0.05)。就马氏珠母贝的壳长日平均增长率而言,PA3、PA9组与P组间没有显著差异(P>0.05),但PA6组显著大于P组和PA3组(P<0.05)。PA6、PA9组马氏珠母贝的肥满度显著大于P组(P<0.05),PA3与P组间无显著差异(P>0.05)。紫海胆的特定生长率、壳长日平均增长率、肥满度在混养与单养的各组间均无显著性差异(P>0.05)。

注:柱状图上方小写字母不同表示马氏珠母贝相关指标组间差异显著(P<0.05),小写字母相同表示差异不显著(P>0.05);柱状图上方大写字母不同表示紫海胆相关指标组间差异显著(P<0.05),大写字母相同表示差异不显著(P>0.05)。
2.2 紫海胆对污损生物的去除效果
由图2可见,紫海胆各混养组养殖网笼污损生物的覆盖面积均小于单养网笼。特别是PA6、PA9组,其养殖网笼相比P组而言,污损生物覆盖面积明显减少,除了最底部隔板等紫海胆无法到达的区域外,网笼笼衣的清洁度都很高,基本没有能造成网孔堵塞的污损生物存在。

注:a图中从左至右依次为P、PA9、PA6、PA3组网笼;b图中从左至右依次为P、PA3、PA6、PA9组网笼隔板。
由图3可见,PA3、PA6、PA9组网笼的附着污损生物量均显著小于P组(P<0.05)。与对照组P相比,混养组PA3、PA6、PA9网笼的污损生物量分别减少了(52.5±12.4)%、(63.3±15.7)%、(70.5±13.1)%。混养组马氏珠母贝壳表附着的污损生物量分别减少了(45.4±9.6)%、(62.9±7.5)%、(68.1±11.9)%,其中PA6、PA9组壳表附着污损生物量显著小于P组(P<0.05)。
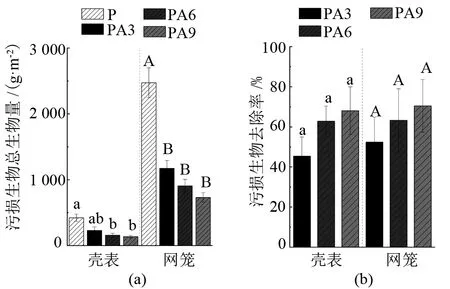
注:小写和大写字母分别代表壳表数据组间差异和网笼数据组间差异情况,字母不同表示差异显著(P<0.05),相同表示差异不显著(P>0.05)。
2.3 贝体壳表附着污损生物优势种及紫海胆对污损生物优势种的去除率
混养组网笼及贝体壳表附着污损生物优势种的去除率见图4~图5。P组网笼附着污损生物优势种为草苔虫(Bugulasp.)、密鳞牡蛎(Ostreadenselamellosa)、网纹藤壶(Amphibalanusreticulates)、巴西埃蜚(Ericthoniusbrasiliensis)、粘附山海绵(Mycaleadhaerens)。马氏珠母贝壳表污损生物优势种为草苔虫、密鳞牡蛎、粘附山海绵、瘤海鞘(Styelasp.)、齿缘牡蛎(Dendostreafolium)、河蜾蠃蜚(Corophiumacherusicum)、巴西地钩虾(Podocerusbrasiliensis)。从图4可以看出,PA3、PA6、PA9组紫海胆对P组网笼的污损生物优势种草苔虫、巴西埃蜚、粘附山海绵均有很好的去除效果,对巴西埃蜚的去除率达到了100%。但PA3、PA6、PA9组优势种网纹藤壶的生物量与P组相比分别增加了(15.7±3.2)%、(63.9±7.2)%、(70.2±9.1)%。紫海胆对马氏珠母贝壳表污损生物优势种草苔虫、巴西地钩虾、河蜾蠃蜚、瘤海鞘、粘附山海绵的去除率均达到50%以上,但是PA3、PA9组密鳞牡蛎的生物量相较P组分别增加了(8.7±1.2)%、(28.6±2.7)%。

图4 紫海胆混养组相比于马氏珠母贝单养组网笼污损生物优势种生物量去除率
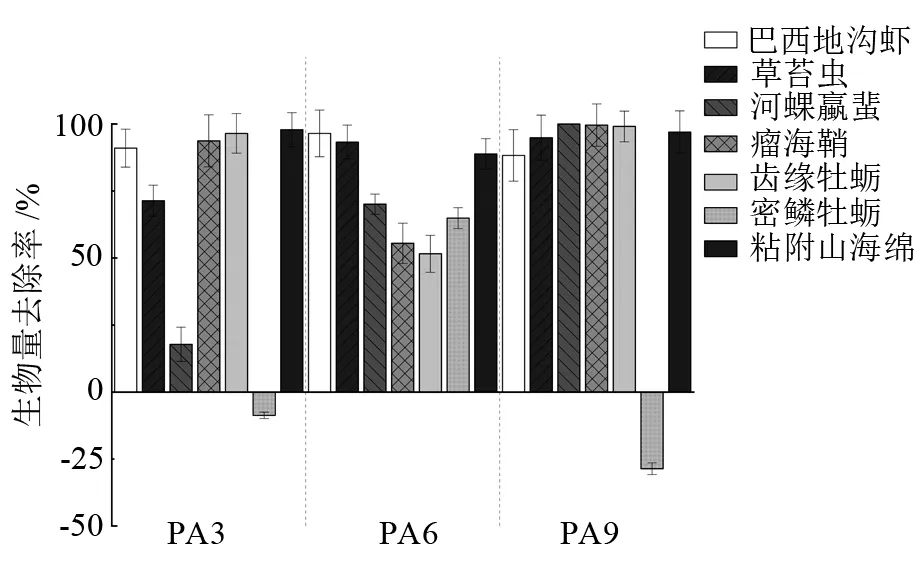
图5 紫海胆混养组相比于马氏珠母贝单养组壳表污损生物优势种生物量去除率
3 讨论
3.1 紫海胆不同混养密度对污损生物的去除情况
大亚湾属于亚热带海域,污损生物常年栖息,通常第1个污损生物高峰期在5—8月,10—12月为第2个暴发期[16]。本试验通过连续4个月的研究观察,发现在污损生物中,草苔虫、密鳞牡蛎、网纹藤壶、巴西埃蜚、粘附山海绵为网笼附着优势种,草苔虫、密鳞牡蛎、粘附山海绵、瘤海鞘、齿缘牡蛎、河蜾蠃蜚、巴西地钩虾为壳表污损生物优势种。其中草苔虫为这个时期马氏珠母贝养殖网笼和贝体附着污损生物绝对优势种,附着严重时甚至可以完全堵塞网眼,阻碍网笼内外的水体交换。郑东强等[17]对大亚湾海水养殖箱、笼上附着污损生物的研究也发现,草苔虫是这个时期珍珠贝养殖的重点防除对象。
海胆通常被认为是草食性动物,但随着人们对其食性的深入研究,发现海胆有着广泛的摄食选择。莫宝霖等[18]利用碳氮稳定同位素技术对大亚湾紫海胆进行食性分析,发现其摄食种类包括颗粒有机物(POM)、有机沉积物(SOM)、裂叶马尾藻、底栖硅藻、浮游动植物等。
本研究将大亚湾自然海域紫海胆与马氏珠母贝混合养殖,结果表明,与马氏珠母贝单养对照组(P组)相比,混养组PA3、PA6、PA9网笼附着污损生物的生物量分别减少了(52.5±12.4)%、(63.3±15.7)%、(70.5±13.1)%,而且混养组紫海胆对网笼内马氏珠母贝壳表污损生物的清除率也达到了(45.4±9.6)%、(62.9±7.5)%、(68.1±11.9)%,PA3、PA6、PA9组网笼及PA6、PA9组贝体污损生物的生物量均显著小于P组(P<0.05)。这说明利用紫海胆能有效防除污损生物在养殖网笼和贝体的附着,且对污损生物的清除率与紫海胆混养密度呈正相关。但是,PA3、PA6、PA9组网笼上的网纹藤壶及PA3、PA9组贝壳表面密鳞牡蛎的生物量与P组相比反而增加了,推测原因,可能是混养组紫海胆将其他污损生物大量去除,导致网纹藤壶和密鳞牡蛎竞争对手减少,生存空间增大。
3.2 混养试验组和对照组养殖马氏珠母贝和紫海胆的生长特性
本试验中,各混养组及P组间马氏珠母贝的存活率无显著差异(P>0.05),而紫海胆的存活率均接近100%,组间无显著差异(P>0.05),表明混养紫海胆并不会对马氏珠母贝和紫海胆的存活造成影响。PA6、PA9组马氏珠母贝的特定生长率、肥满度均显著高于P组(P<0.05),PA6组的壳长日平均增长率也显著高于P组(P<0.05),究其原因,是混养紫海胆后,网笼附着污损生物特别是绝对优势种草苔虫被大量去除,使得通过网笼的水流加大,增加了马氏珠母贝的食物供应(即浮游生物、溶解氧、有机碎屑的含量更高)[19-21],促进了马氏珠母贝的生长。尤其当紫海胆混养密度为6、9个/层时,能显著影响马氏珠母贝的生长(P<0.05)。紫海胆的特定生长率、壳长日平均增长率和肥满度在混养与单养的各组间均无显著性差异(P>0.05),表明混养不会对紫海胆的生长造成影响。综合污损生物去除率以及马氏珠母贝和紫海胆的生长情况,建议紫海胆的混养密度以6个/层为宜。
