构树BpbHLH149转录因子转录酵母的Cd胁迫响应功能1)
2022-07-25何毅张欣谢牧洪陈晓楠赵恬陈钰莹郑欣悦杨桂燕
何毅 张欣 谢牧洪 陈晓楠 赵恬 陈钰莹 郑欣悦 杨桂燕
(西北农林科技大学,杨凌,712100)
bHLH(basic helix-loop-helix)是由约60个氨基酸组成的一种广泛存在于真核生物的转录因子,其名称来源于两个保守但功能不同的结构域-碱性区域(basic)、螺旋-环-螺旋区域(HLH)。15个氨基酸组成的碱性区域位于bHLH结构域的N端,起识别DNA的作用;两个亲水亲脂α-螺旋和连接环组成的HLH区域位于bHLH结构域的C端,通过与不同bHLH转录因子的α-螺旋之间相互作用形成的异源二聚体[1-3],并与靶基因启动子结合形成二聚体,从而调控植物生长发育、应答胁迫、次生代谢[4]。
植物中bHLH转录因子数量众多、功能多样,在植物生长发育过程的关键调控中起重要作用。如赤霉素合成[5]、根毛发育[6]、植物光形态发生、心皮发育[7]等。bHLH家族基因同时也参与植物对各种不良环境胁迫的应答,在拟南芥(Arabidopsisthaliana)、水稻(Oryzasativa)、烟草(Nicotianatabacum)、小麦(Triticumaestivum)和胡杨(Populuseuphratica)等多种植物中发现响应低温、高盐、干旱等多种胁迫的bHLH家族基因[8-12]。拟南芥bHLH122通过与CYP707A3启动子中G-box/E-box顺式元件结合并抑制其表达,从而增强其抗旱、抗盐碱能力[10]。干旱条件下,水稻的bHLH类转录因子OsbHLH148通过茉莉酸信号途径调节植株的耐旱性[11]。烟草NtbHLH123通过结合NtCBF基因启动子中的G-box/E-box元件并正向调节其表达,从而增强转基因烟草植株的耐冻性[12]。小麦转录因子TabHLH39在转基因拟南芥的过表达提升了植物中的可溶性糖和脯氨酸含量,从而改善转基因拟南芥的耐旱、耐盐和耐冻性[9]。胡杨(Populuseuphratica)PebHLH35基因主要存在于细胞核,通过调节叶片气孔密度、光合作用提高植株耐旱性[8]。可见,bHLH是植株应对非生物胁迫的重要转录因子,通过对其进行鉴定,有助于揭示植株的抗性响应机制。
构树(Broussonetiapapyifera(Linn.) L’Hert. ex Vent)属于桑科构属的落叶乔木,具有速生、适应性强、分布广、繁殖能力强等特点,在重金属污染矿区长势优良,被称为矿区植被恢复的先锋树种[13]。近年,构树响应重金属胁迫的分子机制有相关报道,Xu et al.[14]研究发现Cd胁迫下构树多个基因和路径参与了Cd胁迫响应,如AUX/IAA、ARF、bHLH、bZIP、GRAS、NAC、MYB和WRKY家族基因以及ABA信号和钙信号途径。徐正刚[15]研究发现构树BpTT2和BpMYB6基因的过量表达均能有效改善构树的抗Cd能力。以上研究表明,构树响应重金属胁迫是一个复杂的生理生化反应过程,但具体的分子机制有待进一步深入。为充分理解构树在重金属污染土壤修复中的作用,掌握构树重金属响应机制,本研究通过在构树bHLH家族成员挖掘的基础上,选取响应Cd胁迫表达上调的1个bHLH成员(命名为BpbHLH149)进行Cd胁迫下的表达和转基因酵母的抗Cd能力分析,以期为构树用于Cd污染修复提供理论依据。
1 材料与方法
1.1 材料及处理
植物材料:同批次构树组培苗转移至土壤生长至2 a。
处理方法:在相同条件下,用150 μmol·L-1的CdCl2溶液对构树进行胁迫处理,处理后第0、3、12、24、48、72 h等时间分别取其根、叶和茎,用液氮速冻后保存于冰箱-80 ℃备用。每个处理包含5棵植株。
1.2 BpbHLH149基因鉴定与分析
从在CdCl2胁迫下的构树转录组筛选出的上调表达基因中,筛选bHLH家族成员,选择其中1条表达值较高的bHLH基因(命名为BpbHLH149)进行分析。BpbHLH149基因开放读码框(ORF)用ORF finder(http://www.ncbi.nlm.nih.gov/gorf/gorf.html)确定。根据BpbHLH149基因ORF两端序列设计引物BpbHLH149-F和BpbHLH149-R(表1),进行PCR扩增。经PCR及测序确定并利用Expasy Prot-Param(http://web.expasy.org/protparam/)对确认的BpbHLH149基因序列特征进行分析。拟南芥的bHLH家族成员在Tair(The Arabidopsis Information Resource,https://www.arabidopsis.org/)下载;利用MEGA7构建系统进化树。使用CD-Search(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)对BpbHLH149的保守结构域进行分析。利用Swiss Model(https://swissmodel.expasy.org/)推测其蛋白三维结构。
1.3 BpbHLH149基因响应Cd胁迫的表达分析
采用CTAB方法提取各样品总RNA。RNA经DNA消化酶后利用Prime Script TM RT reagent Kit(CWBIO,康为世纪,中国)反转录为cDNA,并经10倍稀释后用作实时荧光定量RT-PCR(qRT-PCR)的模板。qRT-PCR使用SYBR Green Real time PCR Master mix(CWBIO)进行,内参基因为Actin。BpbHLH149定量引物为DL-F和DL-R(表1)。定量反应所用仪器为Applied Biosystems生产的StepOneTMReal Time PCR System,反应程序为:94 ℃ 30 s;94 ℃ 12 s,60 ℃ 45 s,72 ℃ 45 s,45个循环,81 ℃ 1 s,每个样品重复3次。定量结果采用2-△△Ct法[16]进行分析,表示为相对于内参基因相对于对照的表达值。数据使用SPSS软件包分析。样品变异性用标准偏差表示。不同时间点与未胁迫表达量之间的表达差异用T检验分析(p<0.05)。

表1 引物的名称及设计
1.4 酵母表达载体构建及胁迫分析
根据BpbHLH149基因序列特征及pYES2酶切位点特性设计酵母表达载体引物BpbHLH149-JM-F和BpbHLH149-JM-R(表1)。通过PCR反应获得含有酶切位点的BpbHLH149序列,经酶切纯化后与pYES2连接获得重组载体pYES2-BpbHLH149。将pYES2-BpbHLH149转入酵母INVSC1中,经PCR检测和测序确认后的候选菌株提取RNA,通过qRT-PCR检测BpbHLH149的表达水平,选择表达最高的一个株系记为INVSC1(pYES2-BpbHLH149)用于后续分析。转化空pYES2载体的记为INVSC1(pYES2),为对照。本研究所用工具酶均为宝生物(Takara)公司产品。
分别挑取INVSC1(pYES2-BpbHLH149)和INVSC1(pYES2)单克隆置于含2%葡萄糖的SC-Ura液体培养基中,30 ℃震荡培养16 h,收集酵母菌体重悬于含2%半乳糖的SC-Ura液体培养基中,且调整OD600=0.5,30 ℃继续震荡培养至OD600=1.8,分别收集菌体进行胁迫处理。
酵母镉胁迫成活率测定:取相同量的上述INVSC1(pYES2-BpbHLH149)和INVSC1(pYES2)菌体,分别在含有0、0.2%、0.4%、0.6%、0.8%、1.0% CdCl2的SC-Ura液体培养基(含2%葡萄糖)中震荡培养30 h,测定酵母生长活性值(OD600)。数据使用SPSS软件包分析。样品变异性用标准偏差表示。相同处理下INVSC1(pYES2-BpbHLH149)和INVSC1(pYES2)之间的差异显著性用T检验分析(p<0.05)。
2 结果与分析
2.1 BpbHLH149基因基本生物信息
BpbHLH149基因(GeneBank登录号:ON568871)ORF长630 bp,推导其分子质量为22 968.42 u,含有209个氨基酸,理论等电点为11.35。BLAST发现该基因与白梨(Pyrusbretschneideri)转录组中的bHLH149基因(Gene Bank登录号:XP_009337645.1)相同。保守结构域分析发现该基因具有bHLH保守域,表明BpbHLH149蛋白为bHLH蛋白(图1)。经同源搜索并进行进化分析,发现BpbHLH149与拟南芥蛋白AtbHLH147(AT3G17100.1)进化关系较近(图2)。Swiss Model同源建模预测的蛋白三维结构如图3所示。

图1 BpbHLH149蛋白保守结构域

标尺表示蛋白之间的差异性;分支上数值表示同一分支上蛋白的相关性。
2.2 BpbHLH149基因响应镉胁迫的表达
对构树进行CdCl2处理后,BpbHLH149基因在根、茎、叶中均被明显诱导,且不同组织表达水平具有特异性。在根和茎中表达趋势具有相似性,表达水平均随处理时间先升高再降低,但表达水平强弱有差异,根的表达水平远大于茎的表达水平。在根中,BpbHLH149基因在12 h达到最高,为未胁迫时表达量的18.6倍;而在茎中,BpbHLH149基因在24 h达到最高,为未胁迫时表达量的7.1倍。叶中BpbHLH149基因表达趋势与根和茎不同,是随时间延长而逐渐增强,在72 h达到最高,为未胁迫时基因表达量的5.9倍。可见,BpbHLH149基因在镉胁迫下被诱导(图4)。

A.与非冗余PDB结构集比较;B.三维结构模型。
2.3 INVSC1(pYES2-BpbHLH149)和INVSC1(pYES2)在镉胁迫下的生长
将pYES2-BpbHLH149转入INVSC1,挑取9个单克隆用BpbHLH149-F/R进行PCR检测,发现其中7个成功转入BpbHLH149基因,对这7个株系中BpbHLH149基因的表达水平进行检测,发现其中的3号表达水平最高(图5),记为INVSC1(pYES2-BpbHLH149)。对INVSC1(pYES2-BpbHLH149)和INVSC1(pYES2)两种酵母在相同条件下进行不同质量分数的镉胁迫处理,通过比较OD600值分析两种酵母的抗镉能力。结果显示,随着CdCl2质量分数增加,两种酵母的OD600值逐渐变小,但INVSC1(pYES2-BpbHLH149)总是高于INVSC1(pYES2),且CdCl2质量分数为0.6%、0.8%时差异极显著(p<0.01)。表明BpbHLH149基因的表达显著改善了酵母的抗镉胁迫能力(表2)。

图4 BpbHLH149基因在Cd处理下的表达水平

图5 7个转基因酵母株系中BpbHLH149基因的表达水平
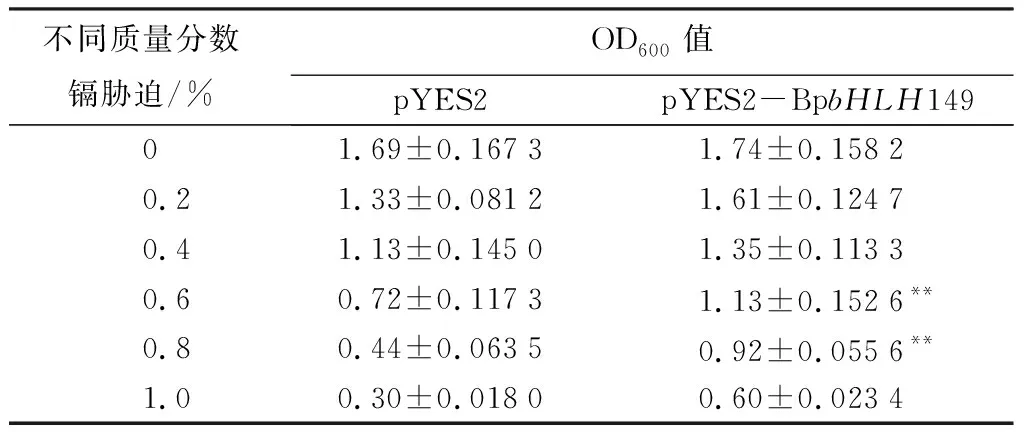
表2 转BpbHLH149基因酵母在镉胁迫下的OD600
3 结论与讨论
土壤生态系统的破坏,不仅导致农作物的减产,而且通过食物链严重危害动物和人体健康,因此必须采取有效方法对土壤重金属污染进行治理。土壤修复技术有物理、化学、生物等修复技术,其中的植物修复技术因具有修复面积广、成本低廉、对土壤扰动小、操作方便、管理简单、无二次污染等优点,备受人们青睐[17]。对重金属富集与超富集能力植物的鉴别与利用,为土壤修复开辟了新的途径。构树适应性强,耐旱、耐瘠,是我国重要的扶贫攻坚树种,因对Sb、Zn、Pb、As、Cd、Cr等重金属的富集能力和转移能力较强,被认为是矿区重金属污染地生态修复与植被恢复较为理想的木本植物[18]。因此,掌握构树的抗逆适应机制,特别是重金属胁迫调控机制对深入了解构树适应性及构树栽培管理具有重要指导作用。由于bHLH是植物中的一类超家族转录因子,成员众多,功能丰富,本研究从构树中鉴定的1条bHLH转录因子基因BpbHLH149,发现其与拟南芥bHLH的众多成员具有较近的亲缘关系(图2)。目前,已有不少研究表明拟南芥bHLH转录因子与多种逆境胁迫调控相关。如,AtbHLH122受高盐、干旱、渗透等多种胁迫诱导,使过表达转基因植株具有更高的抗盐、抗渗透能力[10];AtbHLH20通过多种激素调节途径响应外界脱水胁迫[19];AtbHLH92在高盐、干旱、低温胁迫下的表达量呈现显著上调趋势,且部分依赖ABA信号转导途径[20];AtbHLH38、AtbHLH39通过增加植物根系对镉的螯合作用,提高拟南芥对镉的耐受性[21]。因此,我们初步推测BpbHLH149基因可能参与逆境响应,并发挥重要作用。
为进一步明确BpbHLH149基因是否响应CdCl2胁迫,对构树进行CdCl2处理,分析不同处理时间点该基因在根、茎、叶中的表达。结果显示,BpbHLH149在根、茎、叶中均被明显诱导,且具有一定的组织特异性(图4)。这与其他一些物种的bHLH基因响应重金属胁迫的表达相似。如,芦竹(Arundodonax)bHLH基因被镉显著诱导表达[22];玉米(Zeamays)ZmbHLH105可被锰胁迫诱导[23];鹰嘴豆(Cicerarietinum)bHLH家族蛋白可被铬显著诱导[24]。由于基因表达通常可有效预测基因功能,如,核桃(Juglansregia)JrWRKY2、JrWRKY7发现在干旱等逆境下表达水平显著提高[25],后证实它们的过量表达可以提高转基因植株的抗旱性[26-27]。因此,考虑到本研究中BpbHLH149响应Cd胁迫的表达能力及其他相关基因表达情况与功能间的关系,我们进一步认为BpbHLH149是一个Cd响应基因。
为明确BpbHLH149是否具备Cd响应调控能力,将BpbHLH149基因转入酵母,利用酵母表达系统验证其Cd响应功能。结果发现,转入BpbHLH149基因的酵母INVSC1(pYES2-BpbHLH149)在CdCl2胁迫下的生长活性要显著高于对照INVSC1(pYES2)。由于酵母表达系统繁殖快、操作简单,外源蛋白能够被翻译并修饰,因此被广泛应用于快速检测基因抗逆功能的快速检测。如,通过酵母表达系统证实柽柳(Tamarixchinensis)TheIF1A基因能有效提高转基因酵母的抗干旱、盐碱、氧化、重金属及极端温度的能力,认为TheIF1A可能参与柽柳多种抗逆调控过程[28]。Yang et al.[29]将核桃JrWRKY6和JrWRKY53基因分别转入酵母,通过盐、高渗透和热胁迫环境下的耐受性分析,发现JrWRKY6和JrWRKY53能够积极响应非生物胁迫。本研究Cd胁迫下的酵母活性被BpbHLH149基因的表达而显著提高,这与TheIF1A、JrWRKY6、JrWRKY53等基因在酵母表达系统中改善抗逆能力的表现一致,表明BpbHLH149的表达能有效改善转基因酵母的抗Cd能力,BpbHLH149可作为Cd响应的候选基因。这为进一步研究构树bHLH的抗逆机理奠定了基础。
当前,有关bHLH转录因子调控植株抗逆调控的功能机制研究也取得了一定进展,主要涉及脱落酸(ABA)、赤霉素(GA)等激素信号和活性氧(ROS)代谢及相关蛋白转运途径。如,PEG模拟干旱、盐和ABA诱导了小麦根和叶中TabHLH1表达;与野生型(WT)相比,TabHLH1过表达的烟草在干旱胁迫、盐胁迫和外源ABA介导下的透液积累和细胞内活性氧(ROS)稳态进而有助于提高烟草对干旱和盐胁迫的耐受性得到明显改善[30]。拟南芥中AtbHLH112的转录水平与其耐盐性和耐旱性呈正相关,通过增强P5CS基因的表达和降低P5CDH和ProDH基因的表达以增加其细胞内脯氨酸水平来增强应激耐受;AtbHLH112通过识别E-box和GCG-box增强对POD和SOD基因的表达,以提高ROS清除能力[31]。Cd胁迫下,FIT与AtbHLH38或AtbHLH39共同过表达可激活拟南芥中参与重金属解毒的重金属相关蛋白3(HMA3)、金属耐受蛋白3(MTP3)等转运蛋白基因的表达,从而降低重金属对植株的毒害作用[21]。那么,BpbHLH149在调控Cd等胁迫响应中主要涉及哪些信号途径,其调控机制是什么,是否与上述ABA、GA及ROS相关,这些问题将是后续我们利用植物表达系统对BpbHLH149基因调控Cd胁迫响应机制研究的关键内容。
