4株内生拮抗菌对沙地云杉的促生作用1)
2022-07-25狄佳麟姜海燕丛林白娜娜张旭白慧
狄佳麟 姜海燕 丛林 白娜娜 张旭 白慧
(内蒙古农业大学,呼和浩特,010018)
植物内生真菌(Endophytic fungi)生活在植物各类细胞、组织的内部或间隙中,但不会使寄主出现不良症状[1-2]。大量研究表明,植物内生真菌对宿主植物的生长发育具有重要的促进作用[3]。内生真菌能够促进寄主植物的光合作用[4],提高植物酶活性[5],通过代谢活动产生IAA等生长调节剂,合成植物所需要的有机物,诱导愈伤组织[6],凝聚空气或土壤中游离的N、P、K等营养元素,提高植物利用资源的效率,改善土壤微环境等[7-9],直接或间接促进种子萌发和植株生长[10]。
沙地云杉(Piecamongolica)是松科、云杉属常绿针叶乔木,是内蒙古沙地特有树种,也是中国濒危树种之一[11]。为了促进沙地云杉的生长,提高成活率,筛选具有促生作用的内生菌,本研究选取前期筛选的4株具有强拮抗作用的沙地云杉内生真菌为试验材料,通过发酵液浸种和灌根处理,探究内生真菌对沙地云杉的促生活性。
1 材料与方法
供试沙地云杉内生拮抗菌:SDYS36互隔链格孢菌(Alternariaalternata)、SDYS63三线镰刀菌(Fusariumtricinctum)、SDYS95产黄青霉菌(Penicilliumchrysogenum)、SDYS180聚多曲霉菌(Aspergillussydowii)。4株沙地云杉内生拮抗菌由内蒙古农业大学林学院森林病理实验室提供[12-13]。
供试沙地云杉种子由赤峰市白音敖包自然保护区提供。除杂后挑选种实饱满、色泽正常的健康种子,低温保存备用。种子质量为0.011 4 mg/粒。
主要培养基:PDA固体培养基、PD液体培养基。
1.1 菌株培养
在超净工作台内将沙地云杉内生真菌供试菌株切成0.2 cm×0.2 cm的菌块,接种于已灭菌的PDA平板进行纯培养;无菌条件下,用5 mm打孔器在培养7 d的供试菌株平板边缘打成圆形菌饼,接种于加有100 mL灭菌后的PD液体培养基的250 mL三角瓶中震荡培养5~7 d,28 ℃、180 r/min得到发酵菌液备用。
1.2 沙地云杉内生拮抗菌促种子萌发试验
取沙地云杉种子,经质量分数0.5%高锰酸钾消毒处理后,分别用每种拮抗真菌培养液(试验组)、培养基(PD对照组)和清水(CK对照组)浸泡24 h,备用。
采用纸上萌发试验[14],以无菌滤纸为芽床,将处理后的种子分开放置在不同培养皿中,毎皿100粒,并设置3组重复,放置在光照培养箱中22 ℃培养7 d,并间隔24 h用滴管加入少量无菌水保湿,加入无菌水量以刚好被滤纸吸收为宜。每天观察种子发芽情况,7 d后记录发芽数量,计算萌发率,分析拮抗菌对种子萌发的影响。
1.3 沙地云杉内生拮抗菌促盆栽出苗试验
将V(蛭石)∶V(河沙)∶V(松针土)=1∶1∶1的混合土高压灭菌2 h,以穴盘为育苗容器,采用点播的方式将每种菌液处理后的种子对应均匀种在混合土中,每穴5粒,放至人工气候室(温度(25±2)℃,湿度(40±10)%,16 h光照/8 h黑暗)。待出苗后隔天记录出苗情况,计算出苗率。
1.4 沙地云杉内生拮抗菌促幼苗生长试验
参照侯姣姣等[14]、陈雅琦等[15]、李子英等[16]方法及植物生理学实验指导[17],将带菌种子播种后15 d,对应接入沙地云杉内生拮抗菌发酵液灌根处理,每穴10 mL。通过对比播种后30、45和60 d时,带菌幼苗形态(株高生长量、地径生长量、根长生长量)和生理(可溶性蛋白质量分数、丙二醛质量摩尔浓度、保护酶活性、叶绿素质量分数)的发展变化,分析内生拮抗菌株对沙地云杉幼苗生长的影响。
2 结果与分析
2.1 内生拮抗菌发酵液浸种对种子萌发率的影响
不同内生拮抗菌发酵液处理后,沙地云杉种子萌发率从大到小总体趋势呈现SDYS63(55.33%)、SDYS180(45.23%)、SDYS36(40.55%)、SDYS95(36.27%)、PD(33.10%)、CK(30.11%),4种内生真菌均提高了种子萌发率。其中,SDYS63与其他处理差异性显著(P<0.05),较CK和PD分别提高了25.22%、22.23%;SDYS180作用次之,较CK和PD分别提高了14.12%、11.13%;SDYS36和SDYS95的作用不明显,对种子的萌发率较CK仅提高了10.44%和6.16%,较PD仅提高了7.45%和3.17%(表1)。
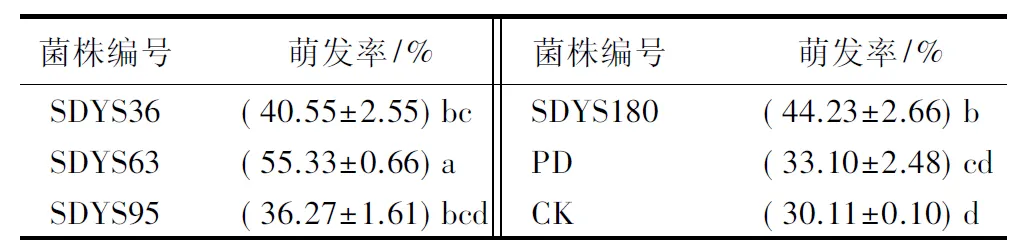
表1 不同内生拮抗菌发酵液浸种后种子萌发率
2.2 内生拮抗菌发酵液浸种对出苗率的影响
不同内生真菌对沙地云杉出苗率均产生了影响。播种5 d时,CK的出苗率为17.5%,SDYS36处理的出苗率高于CK,表现为促进作用,分别为24.0%,SDYS180的出苗率为18.0%,与CK相近,而SDYS63、SDYS95的出苗率低于CK,表现为抑制作用。在播种13 d时,CK的出苗率趋于平稳,为53.5%。SDYS36出苗率最高,为69.5%,较CK高出16.0%;SDYS180和SDYS63的出苗率超过CK,为61.0%和56.5%,高出7.5%和3%;SDYS95较CK低1.0%,仍表现为抑制作用。在播种15 d时,CK出苗率不再大幅上升,所有处理的出苗率均超过CK,表现为促进作用。SDYS36仍出苗率最高,为77.5%,超出CK18.0%;SDYS63次之,为65.0%,超过除SDYS36的其他处理,较CK高5.5%;SDYS180较平稳,不再大幅上升(表2)。
SDYS36在全部过程中的出苗率持续高于CK,表现为明显的促进作用;SDYS63在试验前期虽然低于CK,但斜率偏大,持续上升,在后期超过除SDYS36的其他处理,具有很高的发展潜力;其他处理在后期均超过CK和PD,具有一定发展潜力。
2.3 内生拮抗菌对幼苗株高的影响
播种30 d,幼苗株高由大到小呈现SDYS63、SDYS180、SDYS95、SDYS36、PD、CK,4种内生真菌均对株高生长产生促进作用。其中,SDYS63的促进作用最为明显,较CK提高了13.52%,表现出显著性差异(P<0.05);SDYS36的促进作用最弱,仅3%,较PD和CK未见显著性差异。45 d时,幼苗株高由大到小呈现SDYS36、SDYS63、SDYS180、SDYS95、PD、CK,4种内生真菌均对株高生长产生促进作用,SDYS36和SDYS63表现为显著性差异(P<0.05),较CK分别提升了16.68%和15.63%。60 d时,幼苗株高由大到小呈现SDYS63、SDYS95、SDYS180、SDYS36、PD、CK,4种内生真菌均对株高生长产生促进作用,SDYS63较CK提升了25%,与PD和CK表现出显著性差异(P<0.05)。在3个周期中,SDYS63对沙地云杉幼苗株高生长的促进作用最为突出,且这种作用可以稳定存在,随着时间推移,促进幼苗株高生长的效果会越发明显(表3)。
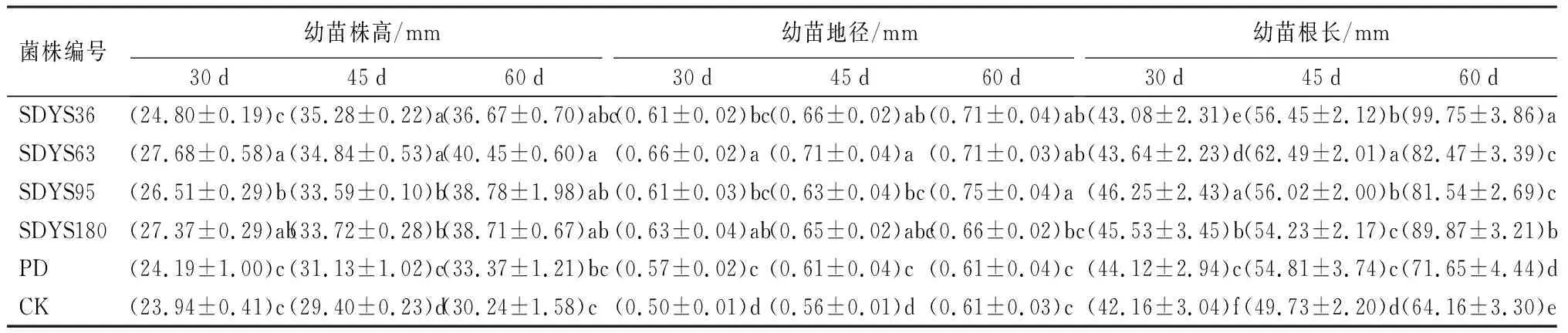
表3 不同内生拮抗菌处理后幼苗株高、地径和根长
2.4 内生拮抗菌对幼苗地径的影响
带菌幼苗的地径均持续高于对照组,但增长幅度在缓慢下降。30 d时,幼苗地径由大到小呈现SDYS63、SDYS180、SDYS95、SDYS36、PD、CK,带菌植株地径较CK分别提升了24.63%、21.74%、18.36%和17.91%。45 d时,由大到小呈现SDYS63、SDYS36、SDYS180、SDYS95、PD、CK,带菌植株分别较CK提升了20.29%、15.53%、13.65%和10.49%,SDYS36超过SDYS180和SDYS95。60 d时,由大到小呈现SDYS95、SDYS63=SDYS36、SDYS180、CK、PD,带菌植株分别较CK提升了18.88%、13.69%、13.69%和10.49%(表3)。
4种内生真菌对幼苗地径的增长均表现为促进作用。在前两个周期,菌株SDYS63对地径增加最多,较PD和CK表现出显著性差异(P<0.05);而在第3周期中,接种SDYS95的幼苗株高超过其他处理,较PD和CK表现出显著性差异(P<0.05)。说明内生真菌的接种对幼苗地径的促生作用在前期作用明显,但并不持续稳定,随着时间推移,内生真菌的影响会逐渐降低。
2.5 内生拮抗菌对幼苗根长的影响
30 d时,幼苗根长由大到小呈现SDYS95、SDYS180、PD、SDYS63、SDYS36、CK,带菌幼苗的根长均高于CK;SDYS95的根长显著高于其他处理(P<0.05),较CK增长了8.86%;SDYS180次之,较CK增长7.42%。45 d时,幼苗根长由大到小呈现SDYS63、SDYS36、SDYS95、PD、SDYS180、CK,带菌幼苗的根长均高于CK;其中SDYS63超过SDYS95,显著高于其他处理(P<0.05),较CK增长了20.42%。60 d时,幼苗根长由大到小呈现SDYS36、SDYS180、SDYS63、SDYS95、PD、CK,带菌幼苗根长均高于对照组,SDYS36显著高于其他处理(P<0.05),较CK增加了35.68%(表3)。
在整个过程中,4株带菌幼苗根长均高于CK。其中SDYS95持续高于PD和CK,对幼苗根生长表现出良好的促生作用。其他带菌幼苗前中期出现低于PD高于CK的表现(如SDYS36),但在后期展现出加速超越对照组的趋势,说明内生真菌对幼苗的根长生长的促进作用表现缓慢,但随时间延长,这种促进作用会逐渐表现并带有不同程度增加的趋势。根据长期发展来看,SDYS36的促生效果最佳。
2.6 内生拮抗菌对幼苗渗透调节的影响
30 d时,带菌幼苗可溶性蛋白质(SP)质量分数均高于对照组,由大到小依次为SDYS63、SDYS95、SDYS36、SDYS180、PD、CK;4种带菌幼苗较CK表现出显著性差异(P<0.05),SDYS63、SDYS95、SDYS36较PD和CK表现出显著性差异(P<0.05),SDYS63中SP质量分数最高,较CK高出12.39%;SDYS95次之,高出10.07%;SDYS36和SDYS180较弱,仅高9.37%和7.73%。45 d时,带菌幼苗SP质量分数均高于对照组,由大到小依次为SDYS63、SDYS180、SDYS95、SDYS36、PD、CK;SDYS63、SDYS95、SDYS180较PD和CK均表现出显著性差异(P<0.05),SDYS63中SP最高,较CK高出8.52%;SDYS180次之,高出5.55%;SDYS95和SDYS36较弱,仅高出5.28%和4.08%。60 d时,带菌幼苗SP质量分数均高于对照组,由大到小依次为SDYS63、SDYS95、SDYS36、SDYS180、PD、CK;带菌幼苗较CK均表现出显著性差异(P<0.05),SDYS63处理SP质量分数最高,较CK高出8.37%;SDYS95次之,高8.08%;SDYS36和SDYS180较弱,仅高出7.22%和6.70%(表4)。

表4 不同内生拮抗菌处理后幼苗SP质量分数
带菌幼苗的SP质量分数在3个周期过程中均高于对照组,其中SDYS63的SP质量分数在3个周期均高于其他试验组,并持续稳步上升;SDYS95和SDYS36处理在第2周期表现出微弱的下降趋势,在后期恢复至高水平,但仍未超过SDYS63处理;说明4种内生真菌对沙地云杉幼苗的要SP质量分数的提高均有促进作用,随着时间的延续,内生真菌表现出不同程度增加的趋势。
2.7 内生拮抗菌对幼苗丙二醛(MDA)质量摩尔浓度的影响
30 d时,丙二醛(MDA)质量摩尔浓度由大到小呈现PD、SDYS63、SDYS95、SDYS36、CK、SDYS180。SDYS63、PD、SDYS95产生MDA质量摩尔浓度较高,比CK提高了16.27%、17.09%和8.70%;SDYS36中MDA质量摩尔浓度与CK相似,仅相差0.91%,SDYS180中质量摩尔浓度偏少,比CK降低了4.17%。45 d时,MDA质量摩尔浓度由大到小呈现CK、SDYS36、SDYS95、SDYS63、SDYS180、PD,带菌幼苗MDA质量摩尔浓度均低于CK;其中,SDYS36和CK几乎相同,仅差0.32%;其他处理较CK减少了11.54%~15.79%。60 d时,MDA质量摩尔浓度由大到小呈现CK、SDYS95、SDYS180、PD、SDYS36、SDYS63,SDYS63的MDA质量摩尔浓度较CK低20.58%(表5)。
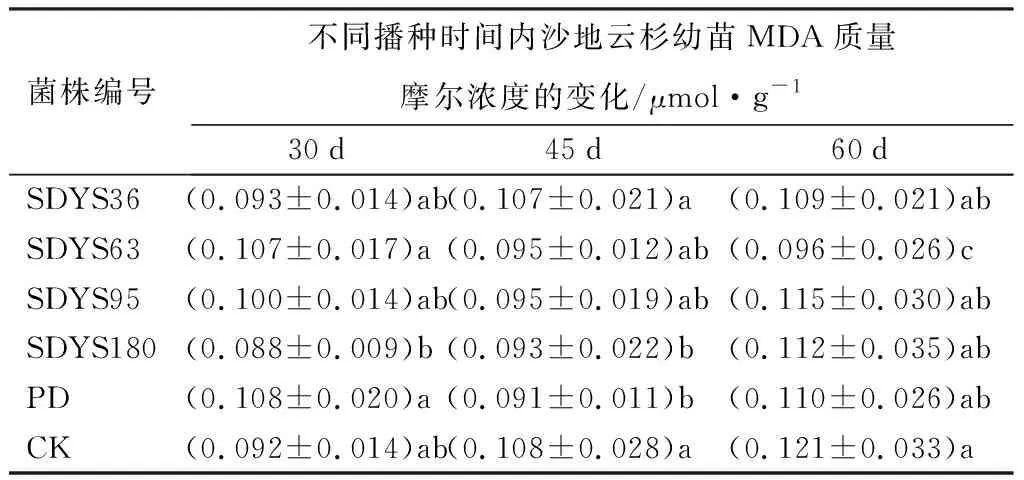
表5 不同内生拮抗菌处理后幼苗MDA质量摩尔浓度
与CK相比,SDYS180在全部周期中表现良好,MDA质量摩尔浓度均低于CK;SDYS63在第1周期中较大,但在中后期表现良好,较大程度上降低了幼苗中MDA的质量摩尔浓度效果明显;其他处理表现出并不稳定的趋势。说明当MDA质量摩尔浓度积累达到一定程度时,内生真菌对幼苗的膜保护机制才会启动,其中菌株SDYS180对MDA最敏感。
2.8 内生拮抗菌对幼苗过氧化物酶(POD)活性的影响
30 d时,带菌幼苗中的过氧化物酶(POD)活性均高于对照组,由大到小呈现SDYS95、SDYS36、SDYS180、SDYS63、CK、PD;SDYS95中POD活性最高,较CK高出79.85%;SDYS36次之,高出62.69%;SDYS180和SDYS63较低,分别高出53.73%和44.03%;45 d时,带菌幼苗的POD活性均高于对照组,由大到小呈现SDYS36、SDYS95、SDYS63、SDYS180、CK、PD;SDYS36和SDYS95较其他处理组表现出显著性差异(P<0.05);SDYS36中POD活性最高,较CK高出302.22%;SDYS95次之,高出231.11%;SDYS180和SDYS63较低,分别高出66.67%和37.78%;60 d时,带菌幼苗的POD活性均高于对照组,依次为SDYS36、SDYS95、SDYS63、SDYS180、CK、PD;SDYS36较其他处理组表现出显著性差异(P<0.05);SDYS36中POD活性最高,较CK高出714.28%;SDYS95次之,高出533.33%;SDYS180和SDYS63较低,分别为190.47%和90.47%(表6)。
3个周期中,带菌幼苗POD活性均明显高于PD和CK对照,其中SDYS36的POD活性相对稳定,虽然前期低于SDYS95,但在中后期,仍保持在较高的水平上,且高于其他组。说明内生真菌可以很好地提高沙地云杉幼苗POD酶活性,但在时间推移中,存在较大差异。
2.9 内生拮抗菌对幼苗超氧化物歧化酶(SOD)活性的影响
30 d时,带菌幼苗中的超氧化物歧化酶(SOD)活性存在很大差异,由大到小呈现SDYS63、SDYS180、PD、CK、SDYS36、SDYS95;SDYS63较其他处理表现出显著性差异(P<0.05);SDYS63中SOD活性最高,较CK高出69.87%;SDYS180次之,高出26.36%;SDYS36和SDYS95的SOD活性低于对照组。45 d时,带菌幼苗的SOD活性均高于对照组,由大到小呈现SDYS180、SDYS36、SDYS95、SDYS63、CK、PD;SDYS180较其他处理组表现出显著性差异(P<0.05);SDYS180中SOD活性最高,较CK高出89.44%;SDYS36次之,高出57.42%;SDYS95和SDYS63较低,分别为45.87%和40.27%。60 d时,带菌幼苗的SOD活性均高于对照组,由大到小呈现SDYS36、SDYS63、SDYS180、SDYS95、PD、CK;SDYS36和SDYS63较其他处理组表现出显著性差异(P<0.05);SDYS36中SOD活性最高,较CK高出152.46%;SDYS63次之,高出147.53%;SDYS180和SDYS63较低,分别高出为122.22%和42.60%(表6)。
带菌幼苗和CK的SOD酶活性均表现为先上升再下降的趋势,其中SDYS63最稳定,在3个周期中始终在较高水平上高于对照组,并且可以保持在基本稳定的水平上,而其他带菌幼苗水平差异较大,具有不确定性。说明内生真菌对沙地云杉幼苗SOD酶活性的提高具有不确定性,其中菌株SDYS63可以明显提高SOD活性。
2.10 内生拮抗菌对幼苗过氧化氢酶(CAT)活性的影响
30 d时,带菌幼苗中的过氧化氢酶(CAT)活性存在很大差异,由大到小呈现SDYS36、SDYS63、SDYS95、PD、CK、SDYS180;SDYS36较其他处理表现出显著性差异(P<0.05);SDYS36中CAT活性最高,较CK高出61.90%;SDYS63次之,高出33.33%;SDYS95高出16.67%;SDYS180的CAT活性低于对照组2.38%。45 d时,幼苗的CAT活性由大到小呈现SDYS36、SDYS95、SDYS63、CK、PD、SDYS180;SDYS36较其他处理组表现出显著性差异(P<0.05);SDYS36中CAT活性最高,较CK高出50.00%;SDYS95次之,高出10.87%;SDYS63高出8.69%;SDYS180的CAT活性低于对照组8.70%。60 d时,幼苗CAT活性由大到小呈现SDYS36、SDYS95、SDYS63、CK、PD、SDYS180;SDYS36较其他处理组表现出显著性差异(P<0.05);SDYS36中CAT活性最高,较CK高出69.44%;SDYS95次之,高出30.56%;SDYS63较低,高出为27.78%;SDYS180的CAT活性低于对照组11.11%(表6)。
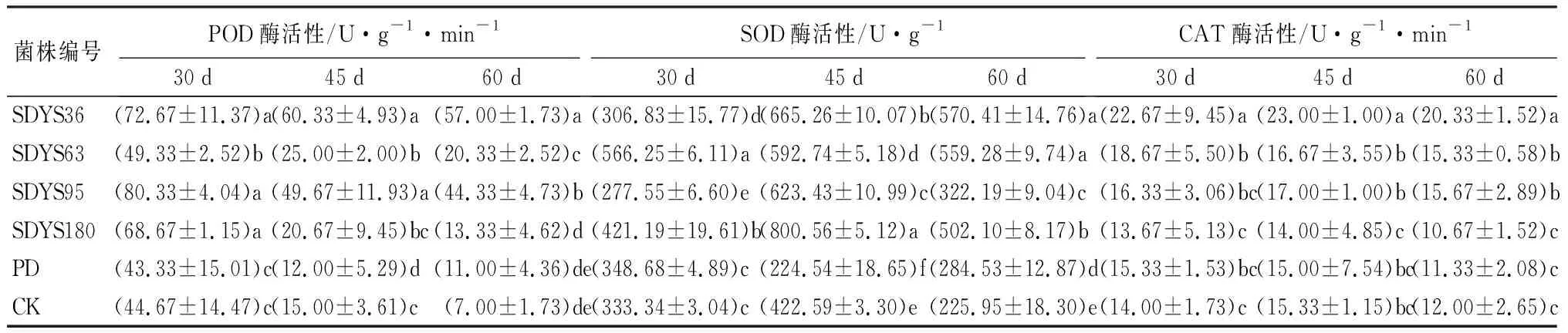
表6 不同内生拮抗菌处理后幼苗酶活性
SDYS36、SDYS63、SDYS95在3个周期中均高于对照组,SDYS36的活性最高,SDYS63和SDYS95低于SDYS36,三者随着时间基本保持相对稳定的水平,说明菌株SDYS36、SDYS63和SDYS95可以提高沙地云杉幼苗CAT酶活性。SDYS180的活性低于对照,说明菌株SDYS180对沙地云杉幼苗CAT酶活性具有抑制作用。
2.11 内生拮抗菌对幼苗叶绿素质量分数的影响
内生拮抗菌对幼苗叶绿素a质量分数的影响:30 d时,带菌幼苗叶绿素a质量分数均高于对照组,由大到小呈现SDYS36、SDYS63、SDYS180、SDYS95、CK、PD;SDYS36较PD和CK均表现出显著性差异(P<0.05);SDYS36中叶绿素a质量分数最高,较CK高出37.47%;SDYS63次之,高出21.62%;SDYS180和SDYS95较弱,仅高出3.43%和2.27%。45 d时,带菌幼苗叶绿素a质量分数均高于对照组,由大到小呈现SDYS36、SDYS95、SDYS63、SDYS180、PD、CK;SDYS36表现出显著性差异(P<0.05),SDYS36中叶绿素a质量分数最高,较CK高出50.07%;SDYS95次之,高出25.18%;SDYS63和SDYS180较弱,仅高出22.81%和22.67%。60 d时,带菌幼苗叶绿素a质量分数均高于对照组,由大到小呈现SDYS63、SDYS180、SDYS95、SDYS36、PD、CK;SDYS63表现出显著性差异(P<0.05),SDYS63中叶绿素a质量分数最高,较CK高出73.23%;SDYS180次之,高出60.18%;SDYS95和SDYS36较弱,仅高出59.24%和56.60%(表7)。
4种带菌幼苗的叶绿素a质量分数在3个周期过程中均高于对照组,其中SDYS36的叶绿素a质量分数在第1、2周期均高于其他试验组,并持续稳步上升,但在第3周期低于其他试验组;SDYS63在第1周期低于SDYS36,但在后期超越其他组并上升至最高;SDYS95和SDYS180稳定上升。说明4种内生真菌对沙地云杉幼苗叶绿素a质量分数的提高均有促进作用,随着时间的延续,内生真菌表现出不同程度增加的趋势;其中菌种SDYS36对提高叶绿素a质量分数表现上升稳定,SDYS63持续上升在后期超过SDYS36,SDYS95和SDYS180促进作用较弱。
内生拮抗菌对幼苗叶绿素b质量分数的影响:30 d时,带菌幼苗叶绿素b质量分数由大到小呈现SDYS36、SDYS63、SDYS95、CK、PD、SDYS180;SDYS36较PD和CK均表现出显著性差异(P<0.05);SDYS36中叶绿素b质量分数最高,较CK高出52.11%;SDYS63次之,高出13.39%;SDYS95较弱,仅高出2.13%;SDYS180中叶绿素b质量分数低于CK9.46%。45 d时,带菌幼苗叶绿素b质量分数均高于对照组,由大到小呈现SDYS36、SDYS63、SDYS95、SDYS180、PD、CK;SDYS36较SDYS95、SDYS180、PD和CK表现出显著性差异(P<0.05),SDYS36中叶绿素b质量分数最高,较CK高出45.43%;SDYS63次之,高出35.72%;SDYS95和SDYS180较弱,仅高出19.99%和16.25%。60 d时,带菌幼苗叶绿素b质量分数均高于对照组,由大到小呈现SDYS36、SDYS63、SDYS180、SDYS95、PD、CK;SDYS36和SDYS63较SDYS95、PD和CK表现出显著性差异(P<0.05),SDYS36中叶绿素b质量分数最高,较CK高出68.88%;SDYS63次之,高出65.43%;SDYS95和SDYS36较弱,高出56.83%和55.08%(表7)。
SDYS36、SDYS63、SDYS95带菌幼苗的叶绿素b质量分数在3个周期过程中均高于对照组,其中SDYS36的叶绿素b质量分数在全部周期均高于其他试验组,并持续上升;SDYS63在全部周期仅次于SDYS36。说明4种内生真菌对沙地云杉幼苗的叶绿素b质量分数的提高有不同程度的影响,其中菌种SDYS36和SDYS63对提高叶绿素b质量分数表现上升稳定。
内生拮抗菌对幼苗总叶绿素质量分数的影响:30 d时,带菌幼苗总叶绿素质量分数由大到小呈现SDYS36、SDYS63、SDYS95、SDYS180、CK、PD,SDYS36表现出显著性差异(P<0.05);SDYS36中总叶绿素质量分数最高,较CK高出40.25%;SDYS63次之,高出20.06%;SDYS95较弱,仅高出2.25%;SDYS180中总叶绿素质量分数与CK相近,仅高出0.99%。45 d时,带菌幼苗总叶绿素质量分数均高于对照组,由大到小呈现SDYS36、SDYS63、SDYS95、SDYS180、PD、CK;SDYS36表现出显著性差异(P<0.05),总叶绿素质量分数最高,较CK高出49.14%;SDYS63次之,高出25.41%;SDYS95和SDYS180稍弱,高出24.14%和21.38%。60 d时,带菌幼苗总叶绿素质量分数均高于对照组,由大到小呈现SDYS63、SDYS36、SDYS180、SDYS95、PD、CK;SDYS63表现出显著性差异(P<0.05),总叶绿素质量分数最高,较CK高出71.36%;SDYS36次之,高出59.53%;SDYS95和SDYS36稍弱,高出58.67%和58.96%(表7)。
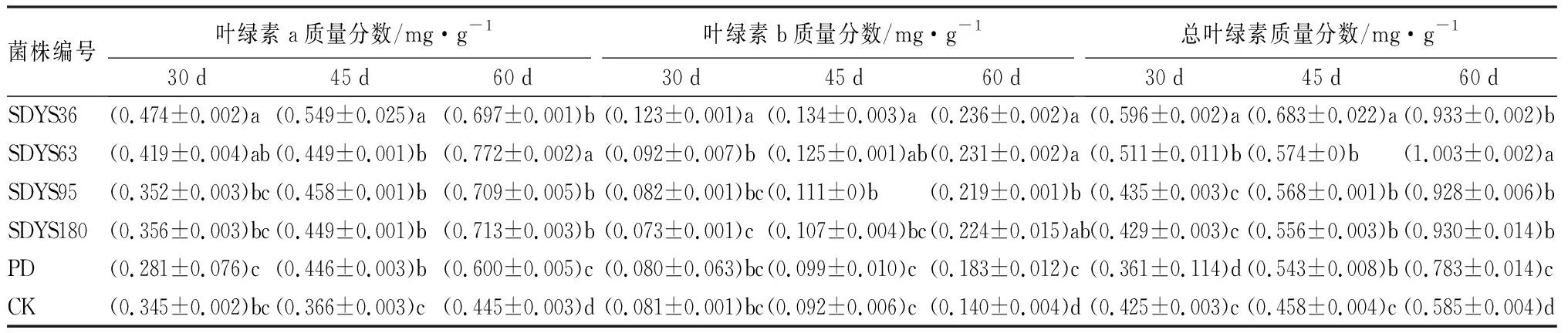
表7 不同内生拮抗菌处理后幼苗叶绿素质量分数
4种带菌幼苗的总叶绿素质量分数在3个周期过程中均高于对照组,其中SDYS36在第1、2周期均高于其他试验组,并持续上升;SDYS63在第1、2周期仅次于SDYS36,在第3周期超越。说明4种内生真菌可以提高沙地云杉幼苗的总叶绿素质量分数,其中菌种SDYS36和SDYS63对提高总叶绿素质量分数表现良好,稳定上升且高于其他内生真菌。
3 结论与讨论
内生真菌除提高寄主植物抗逆性和抗病性,本身及其代谢产物对植物生长同样具有一定的影响。内生真菌自身可以刺激种子萌发和苗木生长,同时也可以产生IAA等生命活性物质促进植物生长。
4株沙地云杉内生拮抗菌发酵液浸种萌发试验中,带菌种子的萌发率均高于CK对照,说明内生真菌能够刺激沙地云杉种子萌发,表现为促进作用;同时这些内生真菌在刺激苗木出苗试验时也表现出促进作用。此结果与傅本重等[18]的研究结果一致。
带菌植株的树高、地径和根长与对照植株比较,带菌植株的形态指标均高于CK对照植株,部分带菌显著高于CK对照植株(P<0.05),说明内生真菌及其代谢产物对沙地云杉幼苗各组织快速生长具有促进作用,且不同内生真菌对幼苗促生作用具有选择性。此结果与杜疏炀等[19]的研究结果一致。
带菌植株的SP质量分数与对照植株比较,带菌幼苗的SP质量分数均持续高于CK对照组,说明4种内生真菌对植株组织细胞的渗透调节均表现出促进作用。此结果与李芳等[20]的研究结果一致。
带菌植株组织中的MDA摩尔质量浓度与对照植株比较,4种内生拮抗菌在较大程度上降低幼苗中MDA的摩尔质量浓度,但这种作用并不稳定。说明当MDA摩尔质量浓度积累达到一定程度时,内生真菌对幼苗的膜保护机制才会启动,不同菌株对MDA的敏感程度不同。此结果与刘鑫铭等[21]的研究结果一致。
带菌植株的保护酶活性与对照植株比较,4种带菌植株之间的酶活力具有很大差异,部分带菌植株表现出较高的酶活力,说明内生真菌对沙地云杉幼苗保护酶活力的提高具有促进作用,但不同内生真菌对保护酶活力的提高具有选择性。此结果与陈玲等[22]的内生真菌促进幼苗酶活力的研究结果一致。但不同处理的抗氧化酶活性出现了不稳定的发展趋势,说明真菌菌液中可能存在一定的细胞毒性[23],且内生真菌通过氧化降解进入植物组织[24-25],具体原因有待进一步研究。
带菌植株的光合色素(叶绿素a、叶绿素b、总叶绿素)质量分数与对照植株比较,发现4种带菌幼苗的叶绿素质量分数均高于CK对照组,说明内生真菌能够促进沙地云杉幼苗产生更多的叶绿素,提高植物的光合效率。此结果与徐欢等[26]的内生真菌促进杉木光合作用的研究结果一致。
4株沙地云杉内生拮抗菌均表现出不同程度的促生活性。内生拮抗菌发酵液浸种处理的沙地云杉种子萌发率较CK提高6.16%~25.22%,菌株SDYS63的促生效果最佳;出苗率较CK提高4.50%~18.00%,SDYS36的促生效果最佳。3个周期健康幼苗的形态生理特性变化,带菌幼苗株高较CK提高17.54%~25.24%,SDYS63的促生效果最佳;地径较CK提高6.81%~18.88%,SDYS63的促生效果最佳;根长较CK提高21.31%~35.68%,3个周期中SDYS95的促生效果最稳定,但在长期发展来看,SDYS36的促生效果最佳;SP质量分数较CK提高6.70%~8.37%,SDYS63促进渗透调节效果最佳;MDA质量摩尔浓度较CK降低7.73%~20.58%,SDYS63的促生效果最佳;POD活性较CK提高90.48%~714.29%,SDYS36的促生效果最佳;SOD活性较CK提高42.60%~152.46%,SDYS63的效果最佳;CAT活性较CK提高27.78%~69.44%,SDYS36的促生效果最佳;叶绿素a质量分数较CK提高56.60%~73.23%,叶绿素b质量分数较CK提高55.08%~68.88%,总叶绿素质量分数较CK提高58.67%~71.36%,SDYS63的促生效果最佳。根据带菌沙地云杉种子和幼苗的生长发育变化,4株内生拮抗菌株中SDYS63对沙地云杉的促生效果最佳。
