伴生东北红豆杉针阔混交林种子雨时空动态
2022-07-20李云红田松岩沃晓棠邵英男刘延坤韩丽冬刁云飞刘玉龙
李云红,田松岩,沃晓棠,邵英男,刘延坤,韩丽冬,陈 瑶,刁云飞,刘玉龙
(1. 黑龙江省生态研究所 森林生态与林业生态工程重点实验室,黑龙江 哈尔滨 150081;2. 黑龙江牡丹江森林生态系统国家定位观测研究站,黑龙江 牡丹江 157500)
种子雨(seed rain,SR)作为植物生活史中不可缺少的部分,连接着植物繁育和后续生活史阶段,其时空动态反映树种自身天然更新策略及树种对环境的适应[1],进而对植物种群的数量、物种配置以及分布格局有着重要影响[2],是研究森林更新过程的关键环节以及植物种群扩散的重要途径[3-4]。
以往对于热带、亚热带以及温带地区种子雨的研究发现,森林群落中物种种子雨时空格局存在变异性。在时间尺度上群落、种群和个体水平上的年际间结实量存在显著差异,即种子雨大小年现象:西双版纳热带雨林中种子雨均有呈现“大小年”交替变化的年际波动,间隔期为1 ~2 a[3];亚热带常绿阔叶林种子雨也显示了很大的年际变异[5-6];小兴安岭温带阔叶红松林中11 个树种均表现出了年际间的差异,一些树种如水曲柳Fraxinus mandshurica、糠椴Tilia mandschurica、红皮云杉Picea koraiensis等在某一年份结实量很小或几乎不结实[7]。种子雨年际变异在不同纬度地区表现不同,有研究发现高纬度森林木本植物种子年际产量变异高于低纬度地区[6,8],也存在高纬度地区种子产量年际变异低于低纬度地区[9],且用以解释这种变异的假说(传粉效率假说、捕食者饱和假说和资源分配理论等)也不尽相同。群落中的生物因素(母树个体结实差异、种子性状、散布能力等)[10]和环境因素(地形、风速风向等)[5,11]均影响种子雨在空间分布上的变异性,大量研究表明种子雨在空间上的变异使种子雨往往呈现聚集分布而不是均匀分布[1,12]。综上,仍需更多研究检验在区域尺度上、不同生境条件下森林群落种子雨时空格局是否存在显著差异。
近年来,随着中国森林生物多样性监测网络的建立,生态学家对我国不同地区不同森林类型的种子雨时空动态进行了大量研究,揭示了一些种子扩散限制和种子结实规律。对东北地区主要森林类型如大兴安岭南段白桦林[13]、小兴安岭阔叶红松原始林[7]、长白山阔叶红松原始林[8]和云冷杉针阔混交林[1]等也相继开展了种子雨时空动态研究,但对东北温带天然次生林,尤其是伴有珍稀濒危树种东北红豆杉Taxus cuspidata的天然次生林的研究较为罕见。本研究依托穆棱东北红豆杉林25 hm2样地,对群落种子雨进行5 a 的定位研究,旨在探讨群落种子雨的组成以及东北红豆杉种子雨特征;群落中主要树种种子雨的时空变异性,变异数据是否支持高纬度地区种子产量年际变化低于低纬度地区的假说;伴生东北红豆杉针阔混交林种子雨的空间分布格局,以期为温带针阔混交林种子生态学大尺度和长时序的研究提供基础数据支持。
1 研究区概况与研究方法
1.1 研究区概况及样地的建立
研究样地位于黑龙江省穆棱市东北红豆杉国家级自然保护区内东北红豆杉集中分布区(130.07°E,43.95°N),参 照CTFS(Center for Tropical Forest Science)样地建设标准,2013 年建立面积25 hm2(500 m×500 m)的监测样地。样地内树种组成丰富,以紫椴Tilia amurensis、色木槭Acer pictum、 臭 冷 杉Abies nephrolepis、 红 松Pinus koraiensis等树种为群落优势种,群落垂直结构明显,是典型的复层异龄林,胸径≥1 cm 的木本植物共有57 种,隶属于22 科38 属,其中东北红豆杉共172株。本区为中纬度北温带大陆性季风气候,年温差较大,气温范围在-44.1 ~35.7℃之间。降水量集中在6—8 月,年均降水量为530 mm,全年无霜期为126 d,年日照时数为2 610 h。
1.2 种子雨动态监测
SR 收集器参照巴拿马[14](BCI)热带雨林50 hm2样地种子收集器的设置方法,于2013 年9月在25 hm2样地内设置161 个种子收集器,收集器高0.8 m,收集框为正方形,面积0.5 m2。
自2014 年1 月开始,每年4 月末收集1 次,5—7 月和11 月每月末收集1 次,8—10 月末每15 d 收集1 次,12 月至次年3 月为雪被期不收集,每年收集11 次数据,共收集5 a(2014—2018 年)。对收集到的种子进行分类鉴定并计数。
1.3 数据处理
1.3.1 种子雨密度
种子雨密度(Seed rain density,DSR,粒·m-2)用每平方米收集到的种子数量表达,即:
DSR=种子数量/收集器面积。
1.3.2 种子雨和群落物种构成的相似性
用SØrenson 相似系数(SI)评价[12],即:
SI=2C/(A+B)。
式中:A、B分别为群落和种子雨中出现的物种数;C为群落和种子雨之间共有的物种数。
1.3.3 种子雨的时空变异系数
用变异系数(CV)计算种子雨数量的时间变异和空间变异[12],即:
CV=σ/μ。
式中:CV 指观测指标的变异系数;μ指观测指标的平均值;σ指观测指标的标准差。
1.3.4 种子雨的空间分布格局
在计算DSR后同时采用插值法[7]和方差均值比率(C0)[15]对种子雨的空间分布格局进行判断,该分布类型假设种子雨空间分布为Poisson 分布,公式如下:
C0=V/m。式中:V为Poisson 分布总体的方差;m为Poisson分布总体的均值。当C0>1 时,种子雨呈现聚集分布;C0=1 时,种子雨呈现随机分布;C0<1 时,种子雨呈现均匀分布。
采用t检验来确定实测值和理论值的偏离程度,公式如下:
t=(C0-1)/(2/(n-1))1/2。
式中:n为SR 收集器个数。当|t|=tn-1,0.05时,表示均匀分布,此时差异不显著;当|t|<tn-1,0.05时,SR 空间分布格局为随机分布;当|t|>tn-1,0.05时,SR 空间分布格局为聚集分布。
SR 年际间差异采用单因素方差分析(one-way ANOVA),用LSD 检验差异显著性(α=0.05)。
使用SPSS 20.0 软件对数据进行统计分析,用SigmaPlot12.0 软件作图。
2 结果与分析
2.1 SR 产量和物种组成
伴生东北红豆杉针阔混交林5 a 种子总数达604 250 粒,DSR达1 501.2 粒·m-2·a-1,成熟种子占总数的87.52%(表1)。种子雨高峰分别出现在2014 年 和2018 年,DSR分 别 为2 656.8 粒·m-2·a-1和3 490.7 粒·m-2·a-1,成 熟 种 子 分 别 占 总 数 的71.69%和98.76%,集中为风力扩散树种的种子。2015、2016 和2017 年的DSR相对较小,尤以2015年最低,DSR仅为155.7 粒·m-2·a-1,成熟种子所占比例为95.80%。
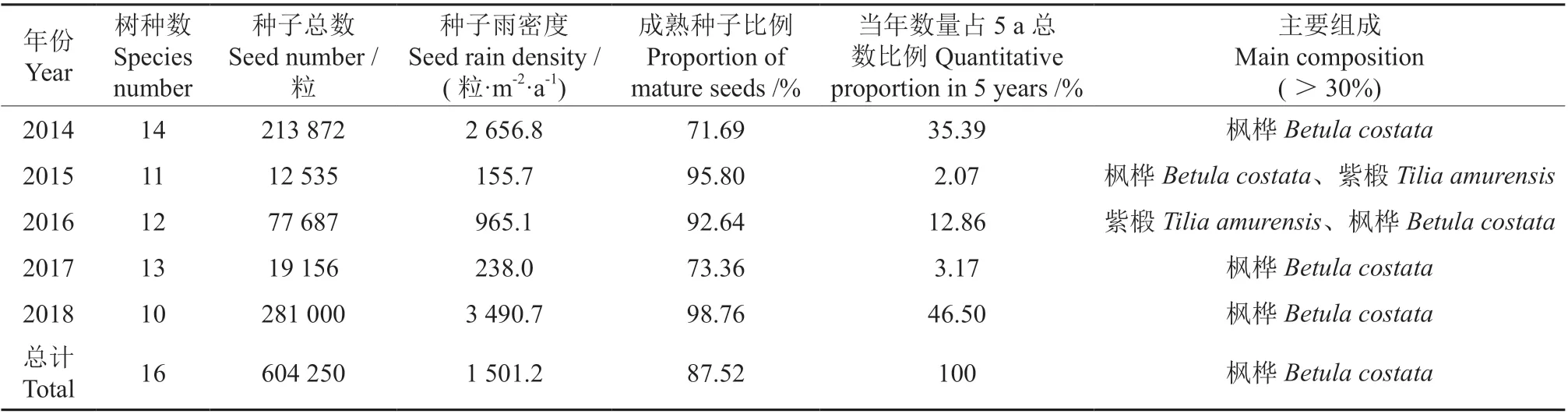
表1 2014—2018 年种子雨数量及主要组成Table 1 Numbers and main compositions of SR in 2014-2018
从组成种子雨的树种上看,在161 个种子雨收集器中5 年间共收集到16 个树种的种子,隶属于9 科11 属。各年间收集到的种类差异不大,从10 个树种到14 个树种。种子雨主要由枫桦Betula costata、紫椴、色木槭、水曲柳等群落的建群树种组成,其中枫桦的种子数量最多,紫椴次之,这两个树种的种子分别占5 a 总数的68.03%和22.00%。共收集到槭树属5 种不同的槭树,色木槭的种子较多,占总数的3.75%;其次为青楷槭Acer tegmentosum和 花 楷 槭Acer ukurunduense,分别占总数的1.66% 和1.61%; 白牛槭Acer mandshuricum和簇毛槭Acer barbinerve种子较少,分别占总数的0.25%和0.20%。针叶树种只收集到了臭冷杉、东北红豆杉和红松的种子,分别占总数的0.38%、0.04%和0.002%。
收集到种子的物种仅占样地木本植物物种总数的28.07%,有41 个树种尚未收集到任何种子,包括胡桃楸Juglans mandshurica、黄菠萝Phellodendron amurense、红皮云杉等主林层树种以及山荆子Malus baccata、山桃稠李Padusmaackii、龙牙楤木Aralia elata等次林层树种和毛榛子Corylus mandshurica、长白忍冬Lonicera ruprechtiana、刺玫蔷薇Rosa davurica、五味子Schisandra chinensis等林下灌木种和藤本植物。种子雨和群落物种构成相似性较低,SI 的5 a 平均值为0.44。
2.2 种子雨的时间动态
从季节来看,伴生东北红豆杉针阔混交林种子雨季节性明显,在收集种子雨的5 个年份均以秋季收集到的种子数量最多(图1)。综合5 a 的种子收集数据,秋季种子产量占全年的65.20%,春季、夏季和冬季分别占1.27%、11.73%和21.80%。
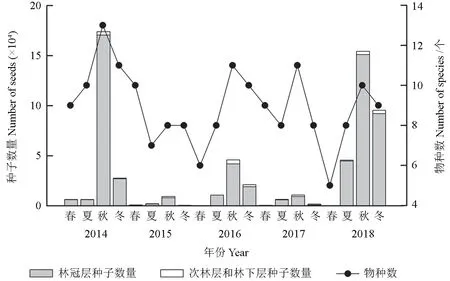
图1 5 年间各季节各林层种子数量和物种数Fig. 1 Number of seeds and species collected in each layer in each season within the 5 years
5 年间收集到的种子大多数来自林冠层,占总数的96.25%,林冠层收集到8 个树种,次林层收集到6 个树种,林下层仅收集到2 个树种。林冠层各年份均在秋季出现种子雨高峰,次林层和林下层各年份仍均在秋季出现种子雨高峰,但数量较少,仅占总数的3.75%。
从2014 年到2018 年,群落DSR呈现先降低后升高的趋势,2014 年和2018 年DSR差异显著(P<0.05),二者显著高于其他年份(P<0.05)。群落中不同物种结实不同步,年际变化格局不同。分析群落中主要树种种子雨的年际动态,可分为3种类型,分别为:Ⅰ,逐年上升型,代表种为色木槭,其2018 年的DSR显著高于其他年份(P<0.05),2017 年和2016 年DSR也显著高于2015 年和2014年(P<0.05);Ⅱ,先下降后上升型,代表种为枫桦,与群落水平DSR的年际变化格局一致,其种子雨高峰出现在2014 年和2018 年,显著高于其他年份(P<0.05);Ⅲ,大小年交替型,此类型物种数最多,主要有紫椴、水曲柳、臭冷杉、花楷槭等。东北红豆杉虽然收集到的种子数量较少,但也表现出了年际间的波动性规律,2 次种子雨高峰间隔2 a,其中2014 年DSR与2017 年差异不显著(P>0.05),但显著高于其他年份(P<0.05)(图2)。

图2 群落和主要优势种DSR 年际动态Fig. 2 Interannual dynamics of the DSR for the community and major dominant species (P <0.05, N=161)
2.3 种子雨空间动态及时空变异
综合分析161 个收集器5 年间收集的种子雨数据,单个收集器收集到的种子数量为715 ~16 209 粒,平均每个收集器收集到3 753 粒种子。单个收集器收集到的树种数为4 ~11 种,平均收集到8 种,多数收集器收集到7 ~9 个树种(图3)。
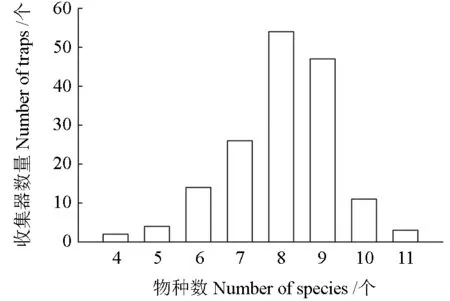
图3 收集器收集到的树种种数Fig. 3 Species numbers collected from all traps
伴生东北红豆杉的针阔混交林在5 年间共收集到16 个树种种子,种子总数≥200 粒的有10个树种,此10 个树种的种子雨空间分布格局均呈现聚集分布状态(表2)。但10 个树种的种子雨在25 hm2样地内的高密度分布区域不同(图4)。枫桦、紫椴、色木槭5 年间收集到种子总数分别为411 025、132 962、22 663 粒,3 个树种种子分布均遍及全样地,但种子雨高密度斑块和次高密度斑块集中分布点不同,依次分别为样地中部偏东南、样地中部偏西南和东北、样地中部偏西北和东南;水曲柳和青楷槭5 年间收集到的种子数超过10 000 粒,分别为12 126 粒和10 025 粒,水曲柳种子雨高密度斑块和次高密度斑块面积较大,主要分布在样地东部,青楷槭则主要分布在样地中部偏南以及样地东北角和西北角;花楷槭种子雨高密度斑块和次高密度斑块主要分布在样地中部、中部偏西北以及样地东南角;臭冷杉种子雨高密度斑块和次高密度斑块主要分布在样地西北角,西南方向也有小面积的分布;白牛槭种子雨高密度斑块和次高密度斑块集中分布在样地东南方向;簇毛槭种子雨高密度斑块和次高密度斑块在样地的中部偏西和偏南、东南和东北均有集中分布区;东北红豆杉5 年间仅在35 个收集器中收集到257 粒种子,SR 高密度斑块和次高密度斑块集中分布在样地中部。

图4 各树种种子雨空间格局(粒·m-2)Fig. 4 SR spatial patterns of trees (Inds·m-2)
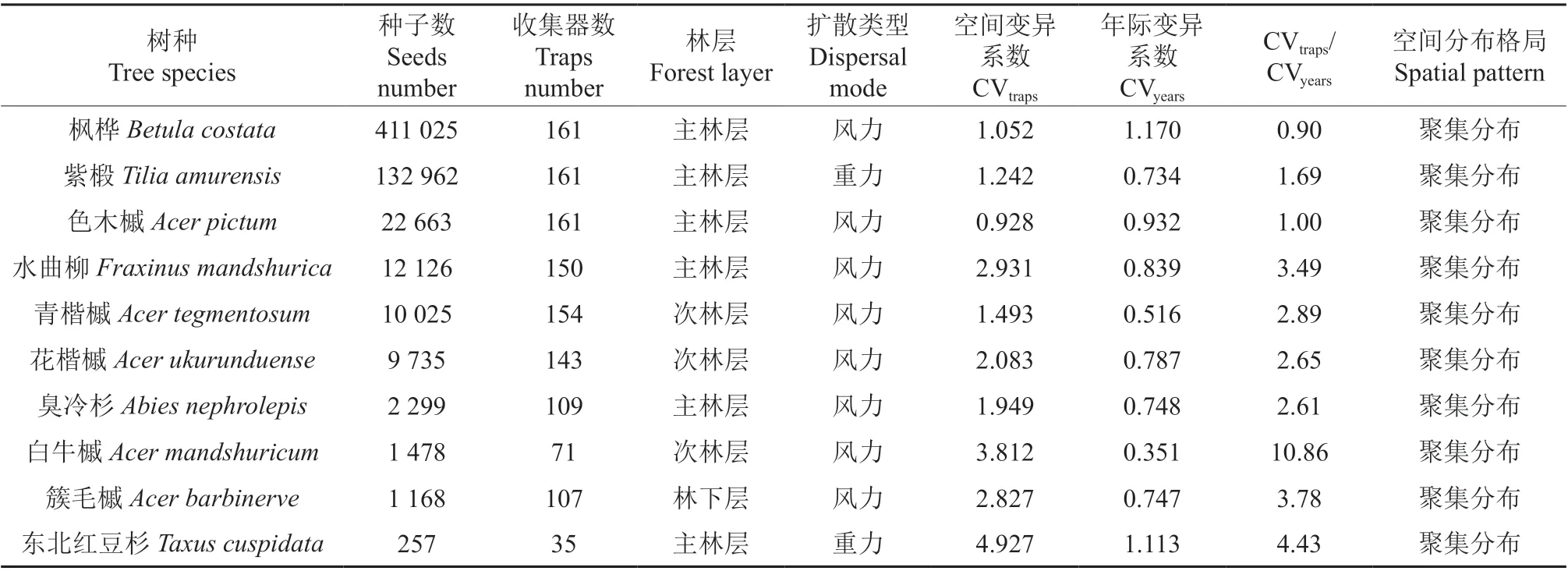
表2 25 hm2 样地各树种相关信息及变异系数†Table 2 Species information and coefficient of variation for trees collected within the 25 hm2 plot
分析此10 个树种的空间变异系数(CVtraps)和年际变异系数(CVyears)发现:SR 的CVtraps在0.928 ~4.927 之间,CVtraps的中位数为2.016;其中,CVtraps大于2 的树种有水曲柳、花楷槭、簇毛槭、白牛槭、东北红豆杉,在1 ~2 之间从高到低依次为臭冷杉、青楷槭、紫椴、枫桦,小于1 的树种为色木槭;种子雨的CVyears在0.351 ~1.170 之间,CVyears的中位数为0.768,东北红豆杉种子雨的CVyears为1.113,枫桦的CVyears为1.170,其余8个树种的CVyears均小于1。10 个树种的CVtraps与CVyears的比值范围为0.90 ~10.86(表2)。
3 结论与讨论
3.1 结 论
伴生东北红豆杉针阔混交林SR 产量在北方温带针阔混交林中处于较高水平,群落优势种对SR贡献占绝对优势,SR 物种组成与群落物种构成相似系数低;10 个树种SR 年际变异系数支持高纬度地区种子产量年际变化低于低纬度地区的假说;10 个树种SR 在空间上均呈聚集分布,但各树种SR 高密度斑块分布位置不同。东北红豆杉为群落的伴生树种,DSR相对较小,存在种源限制。
3.2 讨 论
3.2.1 种子雨产量和物种组成
5 年间,伴生东北红豆杉针阔混交林SR 平均密度为1 501.2 粒·m-2·a-1,高于小兴安岭阔叶红松林(1 235.6 粒·m-2·a-1)[16]、长白山阔叶红松林(728.95 粒·m-2·a-1)[8]和大兴安岭中部针阔混交林(814.1 粒·m-2·a-1)[15]。不同地区的相似林型之间DSR差异较大,可能与各自地区的温度、降水量、地形、土层厚度以及养分条件等有关[17],如肖静等[12]的研究发现,亚热带水热条件和土壤养分条件较好的东部地区常绿阔叶林种子雨产量高于亚热带常绿阔叶林北界,但这与本研究结果不尽一致。本研究样地纬度介于大小兴安岭与长白山之间,但种子雨产量并未明显地表现出随与温度相关的不同纬度变化而变化的趋势,分析原因东北温带针阔混交林DSR可能更受森林树种组成的影响,尤其是基于R对策繁殖的树种,其种子小,数量大,易于传播,本研究中的伴生东北红豆杉针阔混交林与小兴安岭阔叶红松林SR 中贡献最大的树种皆为枫桦,其次为紫椴,二者种子分别占本研究5 a 总数的68.03%和22.00%,可见枫桦和紫椴的种子雨产量决定了本研究区域的种子雨总量。
5 年间共收集到16 种木本植物种子,收集到种子的物种仅占样地木本植物物种总数的28.07%,虽然种子雨和群落物种构成相似性较低,但收集到种子总数≥200 粒的10 个树种中,有7个树种为样地内重要值>3%的物种[18],这在一定程度上反映了群落物种的组成和相对优势度。本研究收集到种子的物种数低于长白山阔叶红松林(51 个物种收集到27 个物种),这有可能是收集时序的原因。绝大多数物种没有收集到的原因可能是这些物种存在种子源限制和散布限制,如胡桃楸、黄菠萝、红皮云杉等乔木树种在群落中重要值<0.5%,存在种子源限制,而毛榛、山荆子、山桃稠李、刺玫蔷薇、五味子等灌木或藤本种子扩散距离有限,导致其种子难以被收集。
3.2.2 种子雨时间动态
种子雨的时间动态主要表现为群落、种群和个体水平上的季节和年际动态变化[19],产生种子雨时间动态的主要因素是温度、降水、日照长度等气候因素和物种自身生态学属性。以往对植物种子雨物候的研究证明了结实存在季节性[20],在热带雨林地区,结实大多在旱季出现高峰[21];在亚热带地区,也有研究发现SR 具有明显的季节动态且均在秋季达到峰值[22]。在温带森林,寒冷的冬季决定了植物结实物候,大多数植物在春季或夏季开花,在同年秋季结实[20]。本研究验证了这一规律,伴生东北红豆杉针阔混交林的主林层、次林层和林下层综合5 a 的种子收集数据,秋季种子产量占全年的65.20%,这与小兴安岭和长白山阔叶红松林[16,23]、长白山不同森林群落中紫椴种群[24]等研究的种子雨季节规律是一致的,都是在秋季达到高峰。这种植物结实的物候规律可能是为种子萌发和幼苗定居提供最佳时机[20]。
种子雨年际变化主要表现为种子产量的大小年差异,反映了不同个体结实的同步性、母树个体间的产量差异[25]以及植物生长发育过程中生殖能量分配的生态适应性。在温带森林,许多树种的种子产量每隔几年出现一次高峰,其他年份少有或没有种子生产,年际产量变异性大。伴生东北红豆杉针阔混交林5 a 种子的年际变化可以证明此现象的存在,群落SR 在2014 年和2018 年达到高峰,在2016 年出现了一次小高峰,交替出现大小年现象。枫桦、紫椴、水曲柳等主要阔叶建群种种子雨也呈现此规律,此外,在观测的5 a 中,色木槭只出现了一次种子雨高峰,东北红豆杉出现了两次高峰,但时间与其他主要建群种不同步。以上现象验证了种子生产丰歉年的资源配置假说[20]和对适宜幼苗定居的有利条件的预测假说[26],种子产量的高低可能代表了植物每年对日照时间、降水量和营养物质等资源的最优化利用,此外,这种不同可使种子错峰萌发,从而减弱各物种在幼苗阶段的资源竞争压力,有利于多物种共存以及生物多样性的维持。
3.2.3 种子雨空间动态及时空变异
本研究中10 个主要树种除枫桦和色木槭外,种子雨的CVtraps均大于CVyears,这与古田山常绿阔叶林[6]和长白山阔叶红松林[8]的研究结论一致。种子雨的空间变异受种源限制和散布限制的影响,这种空间变异使得种子雨在空间上常常呈聚集分布而不是均匀分布[2],本研究的10 个树种的种子雨空间分布格局均呈现聚集分布状态,验证了这一规律。种子空间聚集分布的生态学后果是使同种个体由于受负密度制约而降低种子转化成幼苗的概率,从而有助于其他物种的生存,继而促进物种共存和群落物种多样性的维持[27]。因此,种子需要扩散到远离母树的位置来降低亲缘竞争。而影响种子扩散并最终影响种子雨空间分布格局的因素比较复杂,包括植株高度、种子大小、种子传播方式等生物因素[10]和地形、坡向、风速风向等环境因素[5,11]。一些生态学家认为,传播方式相同的不同物种,小种子比大种子扩散更远[20,28],本研究样地中枫桦的多度和重要值均低于色木槭[18],但二者在161 个收集器中均收集到种子,验证了这一规律。此外,对于槭树科植物和水曲柳等风力传播的物种,树高、风速以及种子的沉降速度也能很好地解释空间分布的差异。
种子年际产量变异大是森林树种普遍的生存策略[29],伴生东北红豆杉针阔混交林5 a 种子的时空动态证明了这一现象的存在。杜彦君等[6]认为,如果CVyears大于0.5,说明变异很大;如果CVyears大于1.0,意味着标准偏差已经大于平均值。本研究的10 个主要树种中,CVyears大于1.0 的有2 个物种,介于0.5 和1.0 之间的有7 个物种,年际变异中位数(0.768)小于巴拿马BCI 样地(1.01)[27]、西双版纳样地[3]和古田山样地(1.79)[6],这与Koenig 等[9]的研究结果一致,即高纬度地区种子产量年际变化低于低纬度地区。但也有研究认为,高纬度地区种子产量年际变化高于低纬度地区[8],因此需要开展更多样地和更为长时序的种子雨研究以获得更加可靠的结论。
由于本研究在5 年间仅收集到16 个树种的种子,可能客观上还不能真正反映伴生东北红豆杉针阔混交林所有树种种子雨动态,此外,本研究还没有对影响格局的生物和非生物因素进行调查分析,在后续的研究中需要增加收集器的数量和更为长时序的观测,并且将影响种子雨分布格局的各种因素考虑在内,对种子散布进行详尽的分析。
