中蜂形态鉴定与分子生物学遗传特性研究进展(2)
2022-07-19徐祖荫
徐祖荫
(贵州徐祖荫养蜂工作室,贵州 贵阳 550001)
(接上期)
3 大量的研究成果印证了第一次全国中蜂资源普查的结果
3.1 研究结果
20 世纪70~80 年代我国第一次全国中蜂资源普查的结果,奠定了分布于我国东方蜜蜂种下分类的基础,其中海南中蜂、阿坝中蜂、滇南中蜂等还曾被一些学者划分为新亚种(杨冠煌、匡邦郁等)。近20 年来,许多学者运用几何形态测量学和分子生物学继续探索,再一次印证了第一次全国中蜂资源普查的结果是基本正确的。
例如,姜玉锁(2007a、b) 分别通过线粒体DNAtRNAleu~COⅡ基因多态性研究、AFLP 分析等手段研究,由于长期海岛隔离、独立进化的结果,海南岛内与国内其他中蜂遗传分化系数Fst均在0.440 3~0.587 4 之间,支持从形态学分析将海南中蜂划为亚种的意见,周姝婧(2012) 通过线粒体DNA 分析,海南主要的单倍型与大陆主要的单倍型不同,大陆以HO2(非编码区Japanl) 为主要单倍型,海南则以HO1 为主要单倍型,占所有单倍型样本的46.15%,是海南岛本土典型的遗传标记,与大陆其他种群存在明显分化,应该是一个独立的种群。
就海南岛内部而言,第一次全国中蜂资源普查时,由于形态特征,尤其是生物学特性上的差别,研究者将分布于海南岛东北部文昌一线的中蜂,划为海南“椰林型” (特点是好分蜂,群势小),而将分布在山区的中蜂,如琼中、乐东、澄迈的中蜂,划分类“山地型” (刘宜钿、梁锦英,1982;杨冠煌等,1984)。近期通过分子生物学检测发现,文昌中蜂单倍型具有独特性,有较高比例的单倍型HO7,且其遗传多样性在岛内检测的样点中最高(周姝婧等, 2012)。微卫星DNA 分析,文昌、屯昌2 样点遗传结构与岛内其他中蜂差异较大,文昌和屯昌样点在AT101 位点以等位基因247 为主,其他样点以等位基因245 为主,与岛内其他样点的遗传分化系数Fst 值大于0.05,达到中度分化水平,聚类分析时(图2),文昌和屯昌样点聚为一支(朱翔杰, 2009;徐新建, 2013),说明将海南中蜂分为2 个生态类型是有遗传学依据的。
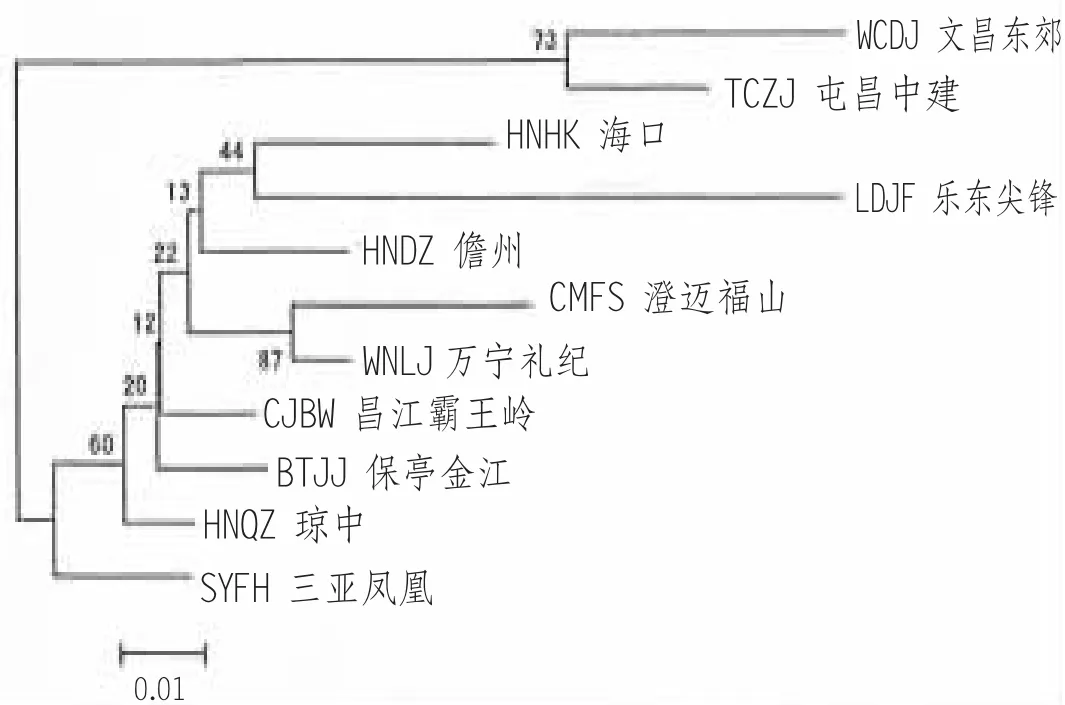
图2 海南中蜂样点间NJ 聚类图(引自徐新建)
长白山中蜂是分布于我国最北端的中蜂种群,那里气候寒冷,冬季极端气温在-20℃以下,蜂群越冬期长,因此生活在那里的中蜂具有独特的生物学特性及遗传特点,长白山中蜂个体较大,体色偏黑,群势大,耐寒力强。许多学者,如谭垦、姜玉锁、王瑞武、薛运波、葛凤晨、于瀛龙等,先后对长白山中蜂开展过研究,发现吉林(安图) 的中蜂,在mtDNA 色素氧化酶Ⅱ(COⅡ) 部分基因编码测序对比发现,其第271 个核苷酸处发生转换;对DNA 进行AFLP 测试,少一个ECOR 1 位点。薛运波通过对长白山中蜂基因组CBS-700(bp) DNA 同源片段克隆及Southern 杂交,提示700 bp 片段可能是长白山中蜂基因组有意义的分子标记。形态分析表明,该型中蜂工蜂前翅外横脉中段常有一个小突起,肘脉指数为6.19,远高于国内其他中蜂3.5~4.17(见表1),这些特征与朝鲜、日本等东方蜜蜂接近而与国内其他东方蜜蜂不同。此外,高鹏飞对我国10 省市mtDNA Cyt b 基因片段扩增测序,聚类分析表明,吉林、海南、云南的中蜂各为一个独立的地理种群。郑秀娟等通过工蜂翅脉进行几何形态测量,同样也证明吉林、海南、陕西中蜂(即长白山、海南和北方中蜂) 各为一个地理种群(图3)。根据许金山等(2020) 对中蜂全基因组SNPs 检测结果,也可将长白山中蜂和分布在我国华北地区的中蜂(TaiV MT,即北方型) 区别开来。

表1 不同样点东方蜜蜂前翅特征比较(引自葛凤晨)
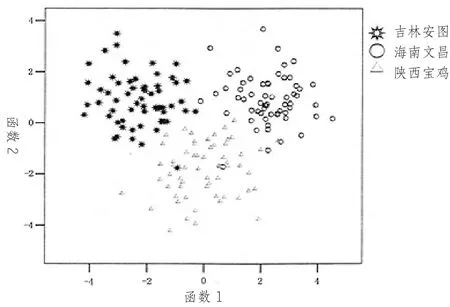
图3 翅脉交点的逐步判别分析3 个样点的散点分布(引自郑秀娟)
在第一次全国中蜂资源普查中,华中型中蜂分布在很大的一片区域内,是所有不同生态类型中中蜂分布地域最广的一个区域,其分布范围位于北纬24°- 34°、东经108°- 109°,即秦岭以南,大巴山、乌蒙山以东,武夷山以西的长江中下游流域的广大地区,主要分布于湖南、湖北、江西、重庆、安徽等省、市及浙江东部、江苏南部、贵州大部、广东、广西北部及四川东北部。这样划分是否正确反映了中蜂种群分布的客观情况呢?郭慧萍等(2016)做了一项工作,即通过8 个微卫星DNA 标记,对17 个样点979 群原始饲养秦巴山区的中蜂开展分析。秦巴山区的地理位置介于北纬30°50′- 34°59′、东经102°54′- 112°40′之间,地跨陕西、甘肃、四川、湖北、重庆、河南等省市(北红英,2014),这个范围与华中型中蜂的分布区域大体相同。秦巴山区植被良好,中蜂数量较多,是研究大面积相似的生境条件下(即种群连续分布、环境均质),中蜂种群遗传多态性和遗传分化的理想场所。
Wright(1978) 提出,当遗传分化系数Fst<0.05 时,表明种群间弱分化或没有分化。基因流参数Nm>1 时表示基因流足以防止遗传分化的发生(Slatkin and Barton, 1989;Wang et al., 2015)。在西方蜜蜂研究中,Nm>2 即表示不存在遗传分化(Nikolova et al., 2015)。而该研究的结果,所有样点间遗传分化系数Fst 值为0.002~0.037,其中,直线距离最远的四川青川和河南陕县之间的距离是650 km,两者间Fst 值为0.021,四川万源与甘肃麦积Fst 值最大,也只有0.037,均小于0.05。聚类分析和遗传结构分析结果也都支持这个观点。该研究基因流参数Nm 为6.51~124.75,Nm 大大高于1~2,足以表示秦巴山区的中蜂未发生遗传分化(郭慧萍等,2016)。分析秦巴山区中蜂未发生明显分化的原因可能是该区域生态条件好,蜜粉资源丰富,环境均质,中蜂的种群数量大(该区域也是我国中蜂的主要产区),又呈连续分布状态,由于中蜂婚飞(10~18 km) 以及蜂群迁徙等原因,种群间基因交流频繁,致在此区间分布的中蜂种群未发生明显分化,其主成分分析紧紧地聚在一起;但秦巴山区(南川、万州、垫江、荣昌) 的中蜂与阿坝中蜂、长白山中蜂遗传分化明显(图4、图5)。于增源等(2013) 等利用线粒体Cyt b 为遗传标准分析重庆地区(南川、万州、垫江、荣昌)的中蜂(同属于秦巴山区) 样本,各地区间遗传距离Da=0.002 4~0.006 0,说明重庆地区的中蜂无遗传分化;许金山等(2020) 对中蜂全基因组SNPs 测定,我国华中地区中蜂各取样点之间的Fst 值在0.001~0.008 之间,都强有力地支持了上述观点。
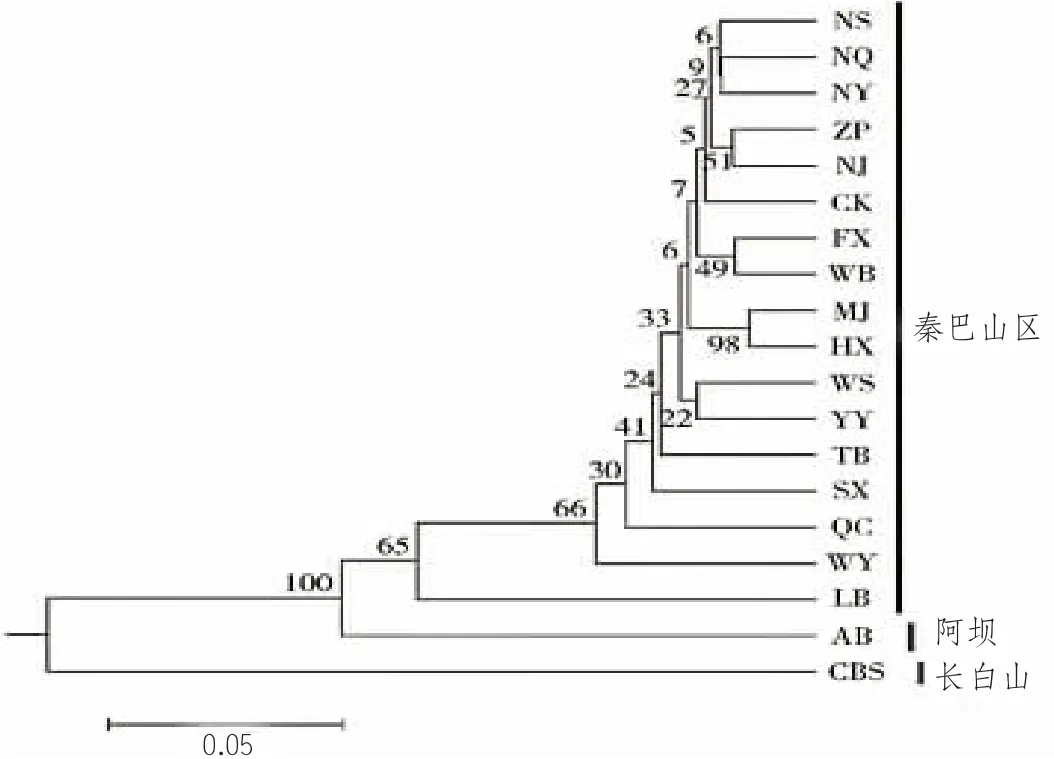
图4 秦巴山区、长白山和阿坝中华蜜蜂UPGMA 聚类图(引自郭慧萍等)
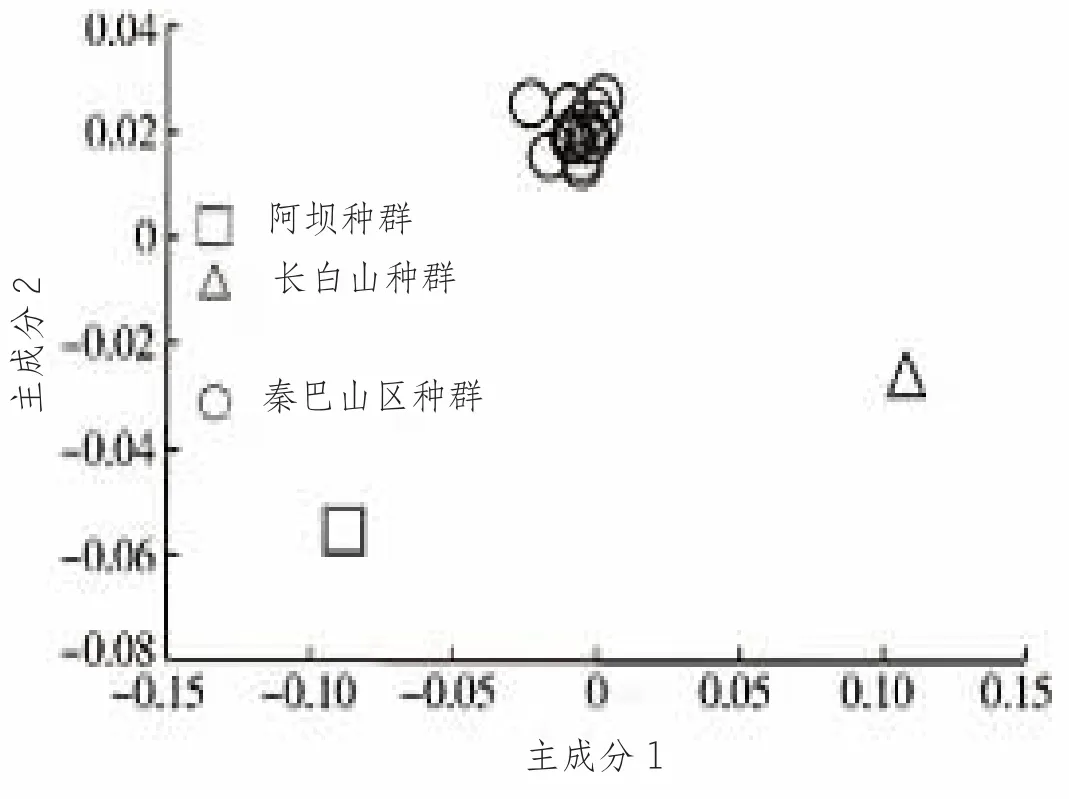
图5 秦巴山区、长白山和阿坝中蜂主要成分分析(引自郭慧萍等)
样本来源及代码:甘肃麦积,MJ;甘肃徽县,HX;陕西凤县,FX;陕西渭滨,WB;陕西太白,TB;陕西留坝,LB;陕西宁陕,NS;陕西宁强,NQ;陕西镇平,ZP;河南陕县,SX;河南南阳,NY;四川万源,WY;四川青川,QC;四川南疆,NJ;重庆巫山,WS;重庆城口,CK;重庆云阳,YY;吉林长白山,CBS;四川阿坝,AB。
此外,陈伟文等(2013) 对从广东、广西和云南采集的中蜂标本,参照鲁特涅的38 个指标测定,并通过计算机所测数据进行聚类分析,证实广东、广西的中蜂为一个地理种群(华南中蜂),而云南中蜂为另外一个地理种群(滇南中蜂)(图6)。张祖芸等通过采自对广西(浦北) 及云南(蒙自等7 县市) 的中蜂标本进行测定分析,也得出同样的结论。
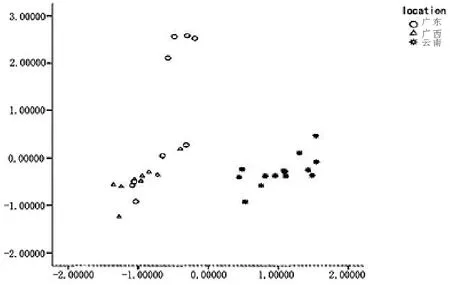
图6 广东、广西、云南中蜂形态指标因素1 至因素2 聚类分析图(引自陈伟文等)
阿坝中蜂的分布范围大致包括四川西北部、青海东部和甘肃东南部,阿坝中蜂的特点是体型大、吻较长、色较深(杨冠煌等,1984),在形态和行为方面都表现出高海拔、低气温的适应性。甘肃珉县、青海民和的中蜂就具有这样的特征(谭垦、祁文忠, 2004;谭垦等, 2005;李华等, 2008)。杨明显等通过甘肃不同地区(珉县、临潭、定西、平凉、奉安) 以及河南、陕西等省的中蜂38个形态指标测定,并聚类分析,将四川九寨沟(阿坝中蜂) 与珉县、临潭聚为一类。赵芳等(2016) 对青海贵德的中蜂进行DNA 测序表明,虽然tRNAlle-ND2、16SrRNA 序列形成特有的单倍型,但基于tRNAlle-ND2、16SrRNA、EFl-α 合并序列的Network 分析,青海贵德的种群与四川九寨沟、马尔康地方种群形成辐射状并与一个mdeian Vector 节点(mvl) 连接,说明他们之间的遗传关系比较接近。许金山等(2020) 对中蜂全基因组SNPs测定表明,分布于海拔3 000 m 的川西北高原中蜂种群(即阿坝中蜂) 和热带滇南种群比较,发现有151 个差异基因,因而影响到两者对不同生境气候的适应性。对这些不同功能类别基因信号通路富集分析,分别富集到HiPPO 信号通路,r 氨基丁酸能通路(基因AcVIAAT) 和海藻糖—磷酸合成酶信号通路。在蜜蜂受到低温胁迫时,其中大多数基因参与了信号传导和能量代谢过程,从而提供了不同蜂种应对寒冷、缺氧环境的能力以及蜂种演化的分子生物学依据。
云南省中蜂分布至少有3 ~4 个类型(云贵高原、滇南、西藏、华中),比较复杂。云南是一个高原、多山省份,境内高山耸崎,河谷深切,东西部、南北部海拔、地貌、植被类型不同。云南大致以云岭山脉及其西段哀牢山为界,将云南分为东西两部分,东部为与贵州省西北角连片形成高原面比较完整的云贵高原,平均海拔4 000 m 左右。西部高黎贡山、碧罗雪山、云岭、怒江、澜沧江、金沙江等相间纵向排列,形成高山纵谷地貌特征。一般北部海拔在4 500~5 000 m,海拔最高处达6 700 m,南部海拔1 500~2 500 m,最低处河口附近只有76.4 m。
由于云南地处低纬度高原,且水平方向上的纬度增加与垂直方向上海拔增高相一致,年均温从5℃(北部德钦,3 000 m) 增至23℃(南部河口,海拔76.4 m),致使云南出现寒、温、热三带皆有的复杂气候,但又由于错综复杂的地形对热量再分配的作用,气温、雨量的分布亦错综复杂。高大的山体可以改变冷空气的方向和速度,翻山后又由于焚风效应,冷空气会变暖变干,因此坡向、坡位、坡度对热量、雨量也起着再分配的作用,更增加了气候的复杂性。气候的复杂性又导致了植被类型的多样性,云南的植被类型大致可分为5 个类型,即南部边缘热带雨林、季雨类型,热带稀树草原旱生类植被类型,南亚热带植被类型,云南高原中亚热带植被类型,滇西北高寒山区植被类型,每一类型又可分为若干亚型。正因为如此,云南素有“动植物王国之称”。由于气候、蜜源植物的不同和变化,也导致生活在其间的中蜂产生了种群分化,中蜂季节性迁徙,夏至高山,冬至河谷,导致南方种群沿河谷北上,北方类群顺山脉南下,其种群分布在一定区域内,出现了明显交错分布的现象。
在20 世纪80 年代全国第一次中蜂资源调查时期,匡邦郁、李有泉等历时8 年,先后在云南省17 个地、州、市,92 个县142个点调查取样、鉴定分析(见表3~11),认为可将云南中蜂分为3 个地理类型,即分布于云南南部1 500 m 以下、气候湿热地区的滇南山地型,以及云贵高原型和滇西北生态型,同时将乌蒙山区中蜂归为云贵高原型。滇东北金沙江河谷的黄色中蜂经与四川畜牧所比对标本,证实与四川盆地为同一类型(即华中生态型)。通过调查鉴定,匡邦郁等认为该省乌蒙山区的中蜂体型较大,吻最长,体色偏黑,能维持大群,采集力强,这与徐祖荫等(1984) 贵州省中蜂资源调查所得结论是一致的(同属乌蒙山区的贵州毕节市、六盘水市的中蜂与其均属云贵高原中蜂,工蜂体色偏黑,吻长5.425 mm;而贵州其余地区华中型中蜂吻长(5.171±0.016) mm,体色偏黄)。张祖芸等(2003) 后来研究证明,云贵两省乌蒙山区的中蜂(云南水富、贵州六盘水) 与同属云南北部的德钦中蜂应归为同一类群,而应与云南南部的中蜂分开(滇南中蜂)。早在第一次全国中蜂资源普查时,云南学者(张正松, 1981;李有泉, 1981;匡邦郁等, 1988) 就已经注意到了云南省中蜂种群间(南部为黄色蜂种,北部为黑色或灰色中蜂) 南北移动、呈现锯齿状交错分布的复杂状况,并认为交汇地带大致在怒江州泸水市的六库附近。而后来谭垦等(2001;2005)、李华等(2012) 运用鲁特涅提出的测定体系,多次调查、测定分析,也肯定了云南境内的中蜂,总体上可分为南北两大类群,一个类群是海拔1 600 m 以下,纬度25°以南的亚热带地区的中蜂,另一个类群是高海拔(1 600 m 以上)、纬度在25°以北地区的中蜂。这与匡邦郁等所提滇南中蜂分布于海拔1 500 m 以下,南北蜂种交汇地点在泸水六库附近(北纬26°以南) 的提法是非常接近的。云南中蜂种群内遗传变异丰富,且在一定区域内(如滇西北地区) 不同种群的中蜂存在交汇混杂的过渡地带,此区域内的蜂种形态呈现出多态性、多型性以及生态地理的多样性,个体大小、体色居于南北两大类群之间(见表2)。正因为如此,所以国内一些学者(谭垦等,2001, 2012;李华等,2012;邱勇等, 2021) 认为云南是亚洲东方蜜蜂的“交混区”“交际区”“分化区”和南北方两大类群的过渡区。
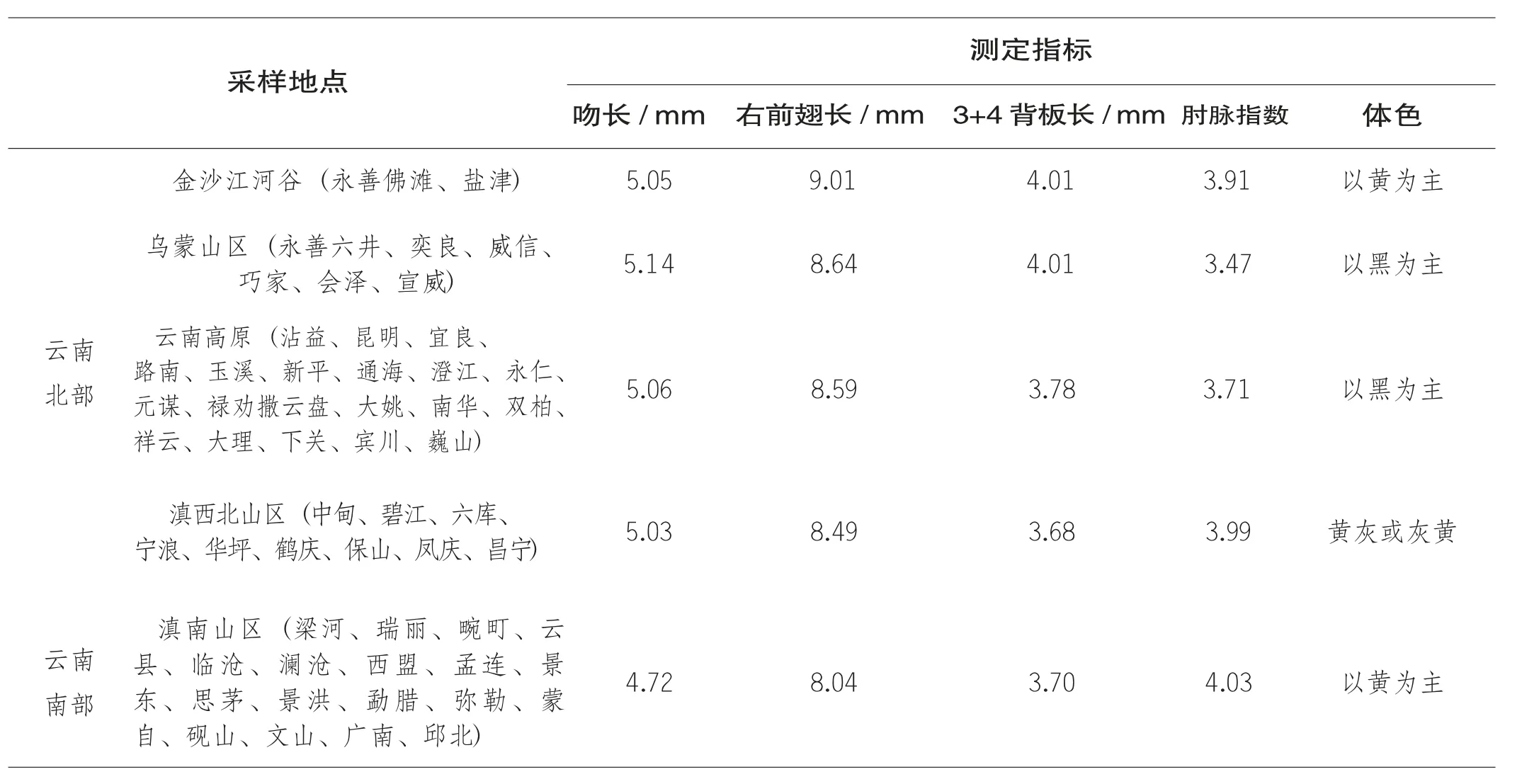
表2 云南省不同区域内中蜂形态指标测定(引自匡邦郁、李有泉等, 1989)
3.2 研究方法的选择
对分布在国内外东方蜜蜂的种下分类工作中,由于使用不同的数理统计方法,例如主成分分析、聚类分析、逐步判别式分析等,常会出现不同的结果。例如在我国东方蜜蜂的形态学分析中,用主成分分析得到4 个类群,用聚类分析得到7 个类群(谭垦等, 2005);对菲律宾东方蜜蜂的研究中,主成分分析中把Palawan 与其他地方分开成独立的一支,而聚类分析中将其归类为北方的Luzon 类群(Tilde et al.,2002)。另有学者用聚类分析法将分布于我国的中蜂分为8 个亚种,其中将分布于我国西北地区的甘肃天水、陕西咸阳、南方的海南、广东(惠莱)、东部的江苏(海门)、中部的河南(波县)、西南地区贵州(锦屏、雷公山) 的中蜂划分为一个类群(华夏亚种),而将贵州毕节、福建南靖、山东泰山、浙江诸暨、四川南江划为毕节亚种(庄德安, 1989),一个类群的分布在一个相当大的地理范围内,呈现出跳跃式、不连续、东西南北皆有的局面,不同的生态类群之间分布严重交叉、混杂,给分类工作造成了严重的矛盾和困惑。一些学者正是因为注意到了上述问题,有针对性地提出了改进的意见和方法,朱翔杰等(2011) 采用已知、公认的地理种群,用不同的方法进行验证,分析统计方法的可靠性和合理性,发现形态鉴定中,逐步判别分析,比较适合作为蜜蜂形态、遗传分析的方法,主成分分析的结果精确度不高,有样点被分入错误类群,且遗传关系也存在偏差。直接采用聚类分析进行蜜蜂形态数据处理的方法不适合蜜蜂种群分化和亲缘关系的鉴定,因为聚类分析的理论背景尚不完备,是一种探索性的统计分析方法,选用不同的计算参数,以及参数数量不同,对于微小指标的差异,都会得到千差万别的结果。因此,在进行聚类分析时,筛选判别能力强的形态变量,寻找合理的距离和分类方法组合等操作(如卡方距离—离差平方和法),才可能得出比较合理的结果。另外,每一种测定方法都有其不足之处及局限性,为了避免产生与实际状况偏差较大的误差,应该将不同的研究、测定方法结合起来,如形态鉴定与分子生物学结合起来,相互参照和补充,这样才能得出客观和合理的鉴定结果。在分析这些结果时,应充分考虑到其生物学意义,符合物种进化演变的规律,否则就会将鉴定的结果,变成一场脱离物种本身真实情况、难以指导生产实际的纯粹数学游戏。
(未完待续)
