环斑猛猎蝽对草地贪夜蛾低龄幼虫的捕食功能分析
2022-07-14胡宗伟冯万祖张浩然李建平吴雨欣于永浩夏鹏亮张建民
胡宗伟,冯万祖,张浩然,曾 曼,陈 黔,李建平,吴雨欣,于永浩,夏鹏亮,张建民*
(1. 长江大学农学院,湖北荆州 434000;2. 广西农业科学院植物保护研究所,广西作物病虫害生物学重点实验室,南宁 530007;3. 湖北省烟草公司恩施州公司,湖北恩施 445000)
草地贪夜蛾SpodopterafrugiperdaSmith属鳞翅目Lepidoptera夜蛾科Noctuidae灰翅夜蛾属Spodoptera昆虫,最早发现于美洲热带区和亚热带区(Sparks, 1979),于2019年1月开始,陆续侵入我国云南、四川、广西、湖南、湖北、安徽等省份,严重威胁了我国的农业生产(刘杰等, 2019a)。草地贪夜蛾具有寄主范围广,取食量大,繁殖力高,适应力强和易地迁飞等特点,因此被联合国粮农组织(Food and Agriculture Organization of the United Nations,FAO)列为了全球预警的重大迁飞性害虫(Deboraetal., 2018; Earlyetal., 2018; Dietal., 2021)。草地贪夜蛾幼虫可取食玉米ZeamaysL.、水稻OryzasativaL.、高粱Sorghumbicolor(L.) Moench、小麦TriticumaestivumL.、棉花Gossypiumspp.等80余种作物,低龄幼虫主要取食植物幼嫩组织,4龄后进入暴食期,可啃食叶片,形成缺刻,或将完整叶片取食殆尽,影响植株正常发育,导致农作物大幅度减产(孔琳, 2020)。我国玉米的种植布局呈现出从南至北依次推移的分布特点,这也从一定程度上促进了草地贪夜蛾在我国的传播(杨普云等, 2019)。目前,我国对于草地贪夜蛾的防控措施以监测预防、农业防治、物理防治、生物防治和化学防治为主(刘杰等, 2019b)。化学防治是目前防控草地贪夜蛾最常用的方法,但是由于化学农药的大量使用,研究人员已经在巴西等南美国家发现了耐受有机磷类、氨基甲酸酯类和拟除虫菊酯类杀虫剂的草地贪夜蛾抗性种群(Yu, 1991; 赵胜园等, 2019)。生物防治主要利用天敌昆虫和生防微生物来控制草地贪夜蛾的种群数量,符合我国“长短结合、标本兼治、分区治理”的防治理念,因而受到众多研究者的普遍重视(Harrisonetal., 2019)。目前,已报道的草地贪夜蛾捕食性天敌有46种,其中捕食性蝽类占总数的41%。猎蝽成虫、若虫皆为捕食性天敌且捕食能力强,因而在草地贪夜蛾的绿色防控方面具有巨大的开发潜力(唐艺婷等, 2019a)。
环斑猛猎蝽SphedanolestesimpressicollisStål属半翅目Hemiptera猎蝽科Reduviidae猛猎蝽属Sphedanolestes昆虫,在国内外均有分布,在国内主要分布于辽宁、山东、陕西、江苏、浙江、湖南、江西、湖北等地。环斑猛猎蝽猎食范围极为广泛,尤其喜食鳞翅目害虫,如玉米螟OstriniafurnacalisHübern、斜纹夜蛾SpodopteralituraFabricius、菜青虫PierisrapaeLinnaeus、杨扇舟蛾ClasteraanachoretaDenis & Schiffermuller、舞毒蛾LymantriadisparLinnaeus、黄刺蛾CnidocampaflavescensWalker等(姚德富等, 1993)。目前,国内仅有3篇文献报道了环斑猛猎蝽的捕食能力,分别为环斑猛猎蝽4龄若虫对烟蚜MyzuspersicaeSulzer的捕食作用、环斑猛猎蝽雌成虫对玉米螟的捕食作用以及雌成虫对玉米螟和蚜虫混合种群的捕食选择性(田静, 2007; 李梦钗等, 2009; 冯万祖等, 2021),国外则无相关报道。环斑猛猎蝽是一种在自然界广泛存在的捕食性天敌,理应可以取食多种农业害虫。因此,进一步开展环斑猛猎蝽对各种害虫的捕食能力研究,对环斑猛猎蝽的合理利用,以及害虫的绿色防控工作具有重要意义。
草地贪夜蛾入侵我国,已对我国农作物生产造成严重影响。而环斑猛猎蝽4龄若虫龄期长达200多天,可在草地贪夜蛾的防控中发挥重要作用。为明确环斑猛猎蝽对草地贪夜蛾的防控潜力,本研究以环斑猛猎蝽和草地贪夜蛾为研究对象,在室内进行了环斑猛猎蝽4龄若虫对草地贪夜蛾1龄、2龄、3龄幼虫的捕食功能实验以及环斑猛猎蝽自身密度对捕食作用的影响。该研究可为在农业生产中合理利用环斑猛猎蝽防控草地贪夜蛾提供理论依据。
1 材料与方法
1.1 供试虫源
在恩施州宣恩县椒园镇(N30.011, E109.443)播种区烟草植株上采集环斑猛猎蝽若虫,在荆州市荆州区(N30.353, E112.190)玉米田中采集草地贪夜蛾幼虫,并在养虫室内进行人工饲养。饲养条件为温度:26℃±1℃,相对湿度:70%±5%,光周期L ∶D=16 ∶8。以人工饲料饲养草地贪夜蛾幼虫(苏湘宁等, 2019),环斑猛猎蝽则以蚜虫、斜纹夜蛾幼虫进行饲养。取发育正常的环斑猛猎蝽4龄若虫、草地贪夜蛾1~3龄幼虫于温度26℃±1℃,相对湿度70%±5%,光周期L ∶D=16 ∶8的人工气候培养箱中进行相关实验(李梦钗等, 2009)。
1.2 环斑猛猎蝽的捕食功能反应
取1头环斑猛猎蝽4龄若虫置于直径90 mm、高15 mm的玻璃培养皿内进行饥饿处理,仅放入经无菌水浸泡的脱脂棉为环斑猛猎蝽提供水分,24 h后放入不同数量的草地贪夜蛾1~3龄幼虫。草地贪夜蛾1龄幼虫的密度为10、20、30、40、50头/皿,草地贪夜蛾2龄幼虫的密度为4、8、12、16、20头/皿,草地贪夜蛾3龄幼虫的密度为3、6、9、12、15头/皿,各处理设置5个重复。取新鲜玉米叶片供草地贪夜蛾幼虫取食,防止草地贪夜蛾幼虫自残,期间观察环斑猛猎蝽对草地贪夜蛾的捕食行为,于24 h后记录环斑猛猎蝽的捕食量。采用Holling II圆盘方程,即Na=aNTr/(1+aThN),拟合环斑猛猎蝽4龄若虫对草地贪夜蛾的捕食功能反应,其中Na为环斑猛猎蝽捕食量,a为瞬时攻击率,N为草地贪夜蛾密度,Tr为试验的总时间即1 d,Th为处理时间,即环斑猛猎蝽捕食1头草地贪夜蛾所需时间(Holling, 1959; 赵英杰等, 2020)。
1.3 搜寻效应
猎物密度是搜寻效应的决定因素,通常情况下,捕食者的搜寻效应与猎物密度呈负相关。利用Holling II型功能反应方程统计相关参数,得到环斑猛猎蝽4龄若虫对草地贪夜蛾幼虫的搜寻效应(S)。其中,S=a/(1+aThN) (丁岩钦, 1994)。
1.4 环斑猛猎蝽自身密度反应
将饥饿处理24 h后的环斑猛猎蝽4龄若虫,密度设置为1、2、3、4、5头/皿。在直径90 mm、高15 mm的玻璃培养皿中放入新鲜玉米叶片,分别接入100头草地贪夜蛾1龄幼虫、50头草地贪夜蛾2龄幼虫、30头草地贪夜蛾3龄幼虫,各处理设置5个重复。24 h后记录各组草地贪夜蛾存活数,评估环斑猛猎蝽的捕食效果。天敌自身密度干扰反应方程为E=QP-m,其中E=Na/NP,E为捕食作用率,Q为搜寻常数,P为环斑猛猎蝽密度,m为干扰常数(Hassell and Varley, 1969)。
1.5 数据分析
利用EXCEL、DPS、GraphPad Prism 8等软件,进行数据统计以及方程拟合,采用Duncan新复极差法进行显著性差异分析。
2 结果与分析
2.1 环斑猛猎蝽4龄若虫对草地贪夜蛾低龄幼虫的捕食行为
环斑猛猎蝽4龄若虫在捕食草地贪夜蛾时,触角和头部会主动朝向猎物,呈试探状,静止片刻后,开始攻击猎物。当猎物个体偏小,如捕食草地贪夜蛾1龄幼虫时,环斑猛猎蝽会用喙刺吸猎物并将其举起,直至将猎物体液吸食殆尽,整个捕食过程较短,约为1~2 min。在捕食草地贪夜蛾2龄幼虫时,环斑猛猎蝽会迅速将口器刺入草地贪夜蛾幼虫头部、胸部或者腹部,使猎物进入麻醉状态,然后将幼虫体液取食殆尽。环斑猛猎蝽在捕食过程中有拖拽行为,整个捕食过程可持续5~15 min。在受到其他环斑猛猎蝽干扰时,环斑猛猎蝽会将猎物拖曳至隐蔽处,独自取食。被捕食的草地贪夜蛾幼虫身体干瘪,体色会迅速呈现黑色。环斑猛猎蝽捕食草地贪夜蛾3龄幼虫的过程与捕食2龄幼虫类似,但环斑猛猎蝽在捕食草地贪夜蛾3龄幼虫时,会多次改变取食位点,整个捕食过程持续25~40 min不等,且在捕食3龄幼虫时,环斑猛猎蝽需要更多的休整时间(图1)。

图1 环斑猛猎蝽4龄若虫对草地贪夜蛾3龄幼虫的捕食Fig.1 Predation of 4th instar nymphs of Sphedanolestes impressicollis against 3rd instar larvae of Spodoptera frugiperda注:A,环斑猛猎蝽4龄若虫开始取食草地贪夜蛾3龄幼虫;B,环斑猛猎蝽4龄若虫完成对草地贪夜蛾3龄幼虫的取食。Note: A, 4th instar nymphs of Sphedanolestes impressicollis began to feed on 3rd instar larvae of Spodoptera frugiperda; B, 4th instar nymphs of Sphedanolestes impressicollis completed feeding on 3rd instar larvae of Spodoptera frugiperda.
2.2 环斑猛猎蝽4龄若虫对草地贪夜蛾低龄幼虫的捕食能力
环斑猛猎蝽4龄若虫对草地贪夜蛾低龄幼虫的捕食作用如图2所示。随着草地贪夜蛾幼虫密度的增加,环斑猛猎蝽的日最大捕食量呈整体上升趋势,但增幅逐渐减缓。在对同一龄期、密度下草地贪夜蛾的捕食功能实验中,环斑猛猎蝽个体间的取食差异较大,且在对3龄草地贪夜蛾的捕食功能实验中表现得更为明显(图2-C)。当草地贪夜蛾1龄幼虫的密度为50头/皿时,环斑猛猎蝽的日平均捕食量达到最大值,为24.2头。当草地贪夜蛾2龄幼虫的密度为16头/皿时,环斑猛猎蝽的日平均捕食量最大,为10.2头。当草地贪夜蛾3龄幼虫的密度为9头/皿时,环斑猛猎蝽的日平均最大捕食量为3.8头。
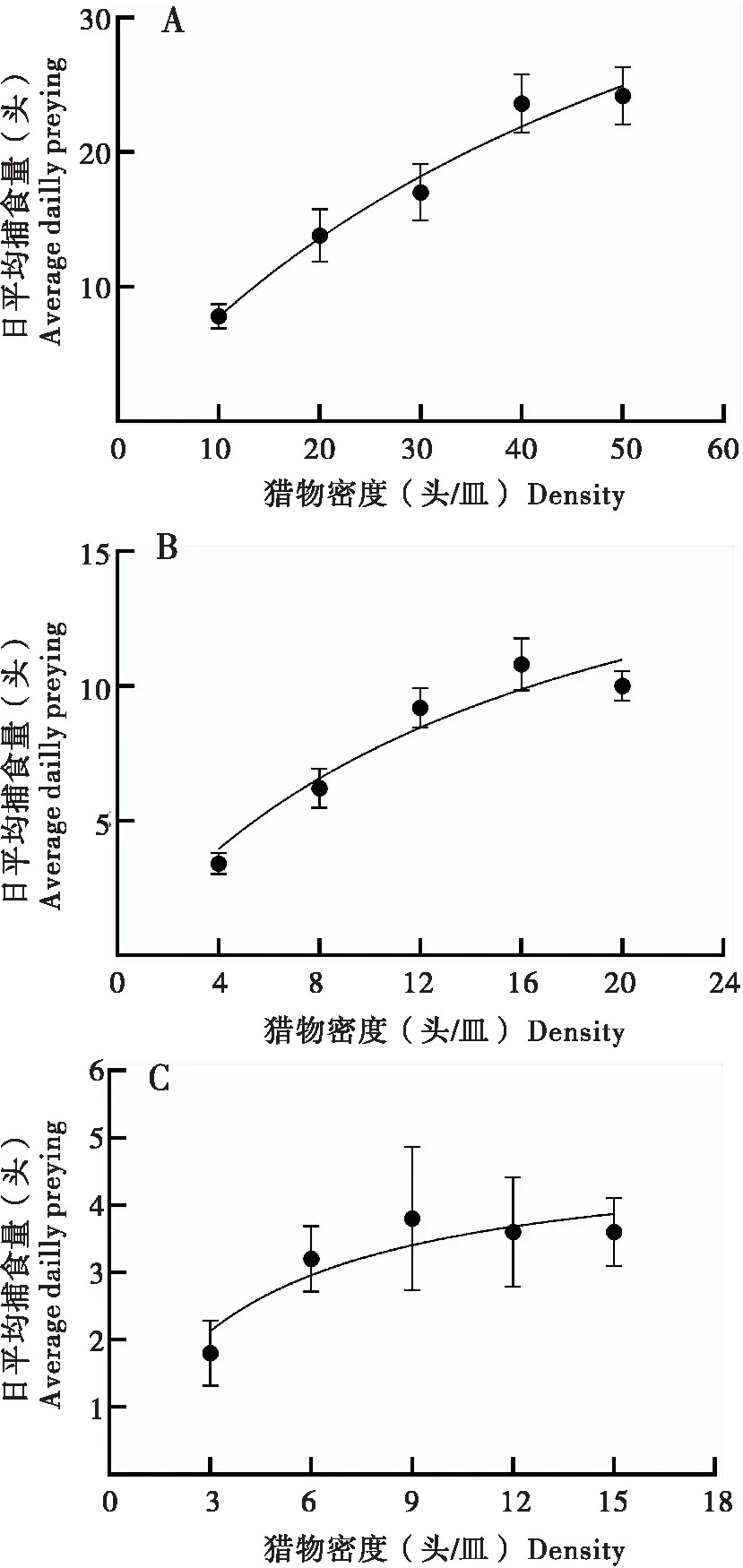
图2 环斑猛猎蝽4龄若虫对草地贪夜蛾低龄幼虫的日平均捕食量Fig.2 Average daily predation of 4th instar nymphs of Sphedanolestes impressicollis against young larvae of Spodoptera frugiperda注:A,1龄幼虫;B,2龄幼虫;C,3龄幼虫。图中数值为平均值±标准误。Note: A, 1st instar larvae; B, 2nd instar larvae; C, 3rd instar larvae. Date of consumption was mean±standard error.
2.3 环斑猛猎蝽4龄若虫对草地贪夜蛾低龄幼虫的捕食功能反应
环斑猛猎蝽4龄若虫对草地贪夜蛾低龄幼虫的捕食功能反应均符合Holling II模型,且各处理功能反应方程的R2均趋近于1,表明环斑猛猎蝽捕食量与草地贪夜蛾幼虫密度显著相关(表1)。环斑猛猎蝽4龄若虫对草地贪夜蛾1龄幼虫的瞬时攻击率、处理时间、日最大捕食量、a/Th值分别为0.900、0.018、55.897、50.313。环斑猛猎蝽4龄若虫对草地贪夜蛾2龄幼虫的瞬时攻击率、处理时间、日最大捕食量、a/Th值分别为1.229、0.050、19.853、24.399。环斑猛猎蝽4龄若虫对草地贪夜蛾3龄幼虫的瞬时攻击率、处理时间、日最大捕食量、a/Th值分别为1.259、0.205、4.871、5.995。结果表明,环斑猛猎蝽4龄若虫对草地贪夜蛾1龄、2龄幼虫的捕食能力显著高于草地贪夜蛾3龄幼虫。

表1 环斑猛猎蝽4龄若虫对草地贪夜蛾幼虫的捕食功能反应Table 1 Functional responses of 4th instar nymphs of Sphedanolestes impressicollis against young larvae of Spodoptera frugiperda
2.4 环斑猛猎蝽4龄若虫对草地贪夜蛾低龄幼虫的搜寻效应
环斑猛猎蝽4龄若虫对草地贪夜蛾1龄、2龄、3龄幼虫的搜寻效应与猎物密度呈负相关(图3)。当草地贪夜蛾1龄幼虫的试验密度为10、20、30、40、50头/皿时,环斑猛猎蝽4龄若虫的搜寻效应分别为0.755、0.681、0.607、0.547、0.499。当草地贪夜蛾2龄幼虫的试验密度为4、8、12、16、20头/皿时,环斑猛猎蝽4龄若虫的搜寻效应分别为0.985、0.822、0.705、0.617、0.549。当草地贪夜蛾3龄幼虫的试验密度为3、6、9、12、15头/皿时,环斑猛猎蝽4龄若虫的搜寻效应分别为0.709、0.494、0.379、0.307、0.258。结果表明,随着单位体积内草地贪夜蛾密度的增大,搜寻效应随之下降。
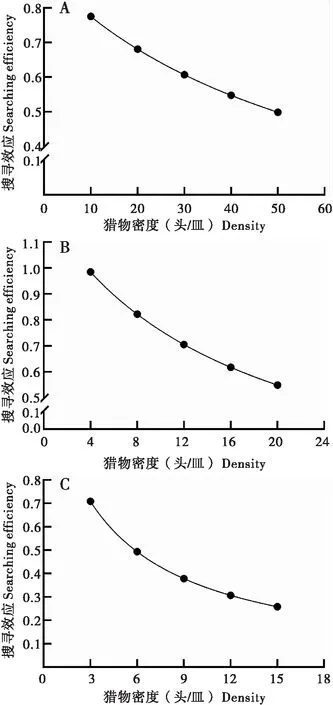
图3 环斑猛猎蝽4龄若虫对草地贪夜蛾低龄幼虫的搜寻效应Fig.3 Searching efficiency of 4th instar nymphs ofSphedanolestes impressicollis against young larvae of Spodoptera frugiperda注:A,1龄幼虫;B,2龄幼虫;C,3龄幼虫。Note: A, 1st instar larvae; B, 2nd instar larvae; C, 3rd instar larvae.
2.5 环斑猛猎蝽4龄若虫自身密度对捕食草地贪夜蛾低龄幼虫的干扰作用
在一定空间且草地贪夜蛾密度固定的情况下,随着环斑猛猎蝽4龄若虫密度的增加,环斑猛猎蝽4龄若虫对草地贪夜蛾1龄、2龄、3龄幼虫的日捕食量逐渐减少,捕食作用率也逐渐降低(表2),表明4龄环斑猛猎蝽个体间存在明显的干扰效应。利用干扰反应方程拟合捕食作用率与天敌密度,结果显示,环斑猛猎蝽4龄若虫对草地贪夜蛾1龄、2龄、3龄幼虫的干扰反应方程分别为E=0.2423P-0.1369(R2=0.966)、E=0.2162P-0.296(R2=0.970)和E=0.129P-0.2041(R2=0.912)。4龄环斑猛猎蝽对草地贪夜蛾低龄幼虫的捕食作用率随天敌密度的增加而降低(图4)。当环斑猛猎蝽4龄若虫在捕食草地贪夜蛾1龄幼虫时,环斑猛猎蝽自身密度对其捕食能力干扰最大,捕食草地贪夜蛾2龄、3龄幼虫时,干扰效应次之。
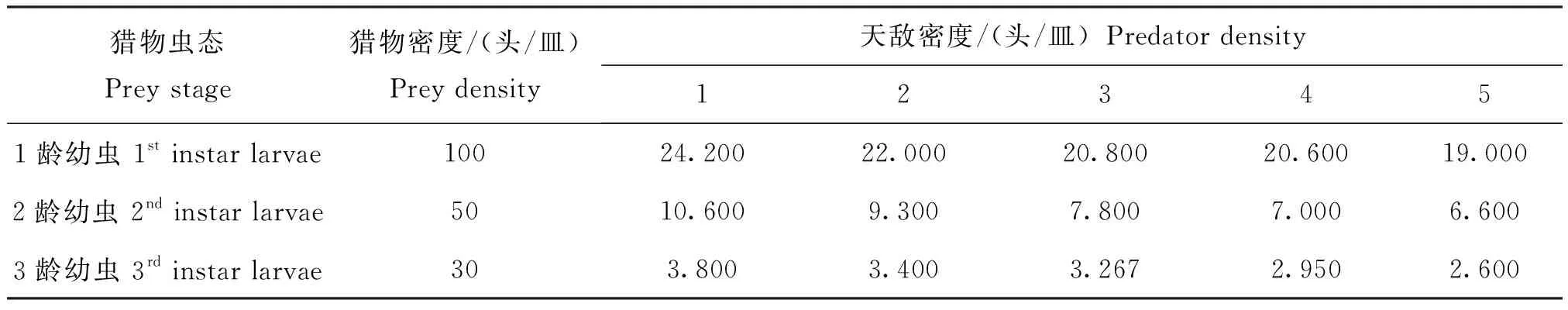
表2 不同密度环斑猛猎蝽4龄若虫对草地贪夜蛾低龄幼虫的日平均捕食量Table 2 Average daily predation of 4th instar nymphs of Sphedanolestes impressicollis under self different density against young larvae of Spodoptera frugiperda
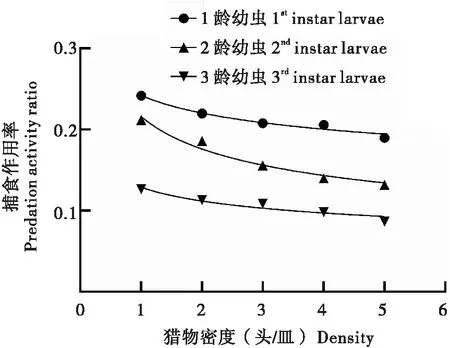
图4 环斑猛猎蝽自身密度对捕食草地贪夜蛾低龄幼虫的干扰作用Fig.4 Interfering effects of 4th instar nymphs of Sphedanolestes impressicollis under self different density against young larvae of Spodoptera frugiperda
3 结论与讨论
在室内条件下,环斑猛猎蝽4龄若虫对草地贪夜蛾1~3龄幼虫表现出良好的搜寻和捕食能力,表明草地贪夜蛾低龄幼虫是环斑猛猎蝽4龄若虫适宜的捕食对象。环斑猛猎蝽4龄若虫对草地贪夜蛾低龄幼虫的捕食行为包括搜寻、捕食、休息等过程,这与黄带犀猎蝽BettamacrostomaDohrn、大红犀猎蝽SycanusfallenStål、蠋蝽ArmachinensisFallou的捕食行为类似(唐艺婷等, 2019b; 侯峥嵘等, 2020; 王亚楠等, 2020)。此外,环斑猛猎蝽喜单独捕食猎物,拒绝共食,当在捕食过程中受到干扰时,环斑猛猎蝽4龄若虫会将草地贪夜蛾低龄幼虫拖拽至隐蔽处,然后再持续吸取猎物体液,猎物最终干瘪死亡。
环斑猛猎蝽4龄若虫对草地贪夜蛾1龄、2龄、3龄幼虫的捕食功能反应均符合Holling II模型,其对草地贪夜蛾1龄幼虫的日最大捕食量为55.897头,而大草蛉ChrysopapallensRambur成虫、东亚小花蝽OriussauteriPoppius雌成虫、大灰食蚜蝇EupeodescorollaeFabricius 2龄幼虫的日最大捕食量分别为13.699头、26.200头、43.480头(徐庆宣等, 2019; Zengetal., 2021; Lietal., 2021)。环斑猛猎蝽4龄若虫对草地贪夜蛾2龄幼虫的日最大捕食量为19.853头,低于龟纹瓢虫PropylaeajaponicaThunberg 4龄幼虫的日最大捕食量(27.6头)(蒋骏等, 2020)。环斑猛猎蝽4龄若虫对3龄草地贪夜蛾的日最大捕食量为4.871头,略高于黄带犀猎蝽4龄若虫的日最大捕食量(3.906头)(王亚楠等, 2020)。从以上结果可知环斑猛猎蝽4龄若虫对草地贪夜蛾低龄幼虫具有一定的捕食能力。在对同一龄期、密度下草地贪夜蛾的捕食功能实验中,不同环斑猛猎蝽个体间的捕食量有一定差异,且在对3龄草地贪夜蛾的捕食功能实验中表现得更为明显,这与环斑猛猎蝽个体间捕食能力的差异密切相关。其次,随着草地贪夜蛾龄期的增加,环斑猛猎蝽的捕食量有逐渐减弱的趋势。出现这一现象的原因在于,相对3龄幼虫,草地贪夜蛾1龄、2龄幼虫防御能力较弱,同时环斑猛猎蝽需要捕食更多的猎物,才能满足自身的生长发育的需求(唐敏, 2019)。环斑猛猎蝽4龄若虫对草地贪夜蛾1龄幼虫表现出最高的捕食量和控害效能,因此在草地贪夜蛾幼虫进入暴食期前,利用4龄环斑猛猎蝽防治草地贪夜蛾1龄幼虫可以压低害虫的种群数量,达到较好的防治效果。此外,本研究发现环斑猛猎蝽5龄若虫及环斑猛猎蝽雌、雄成虫对1~5龄草地贪夜蛾均有较好的捕食能力,相关的捕食功能实验还在进一步研究中。
搜寻效应是捕食者在捕食过程中表现出的一种行为效应。随着猎物密度的增加,捕食者的搜寻效应会降低,捕食者能更快完成对猎物的捕食行为(刘爽等, 2011)。在本研究中,随着草地贪夜蛾1龄、2龄、3龄幼虫密度的增加,环斑猛猎蝽4龄若虫的搜寻效应逐渐降低。同时,搜寻效应曲线逐渐平缓,表明草地贪夜蛾密度对环斑猛猎蝽4龄若虫搜寻效应的影响逐渐减弱。这种现象在天敌与猎物系统中是客观存在的,如大红犀猎蝽3龄若虫和成虫对草地贪夜蛾3龄幼虫的搜寻效应与猎物密度呈负相关(侯峥嵘等, 2020);七星瓢虫CoccinellaseptempunctataLinnaeus成虫对草地贪夜蛾1龄、2龄幼虫的搜寻效应随着猎物密度的增加而降低(孔琳, 2020)。环斑猛猎蝽自身密度对捕食草地贪夜蛾低龄幼虫的干扰试验结果显示环斑猛猎蝽4龄若虫的捕食量及随着猎物密度的增加而降低,其对草地贪夜蛾的搜寻常数分别为0.242、0.216、0.129,干扰系数分别为0.137、0.296、0.204。当环斑猛猎蝽4龄若虫在捕食草地贪夜蛾1龄幼虫时,环斑猛猎蝽4龄若虫自身密度对其捕食能力干扰最大。这与大草岭成虫自身密度对捕食草地贪夜蛾低龄幼虫的结果趋于一致(曹雯星等, 2020)。在一定的生存空间内,随着天敌种群密度的增大,天敌个体间存在剧烈的干扰效应,严重影响其捕食作用率。因此在人工释放环斑猛猎蝽防治草地贪夜蛾幼虫时,应综合考虑环斑猛猎蝽的种群密度对其捕食能力的影响,以达到最佳防控效果。
本试验通过研究环斑猛猎蝽4龄若虫对草地贪夜蛾幼虫的捕食功能反应、搜寻效应,以及4龄环斑猛猎蝽自身密度的干扰作用,明确了环斑猛猎蝽对草地贪夜蛾低龄幼虫的防控能力,为在大田环境中利用环斑猛猎蝽防治草地贪夜蛾提供了理论参考。但是在大田环境下,温度、湿度、作物布局、猎物密度、自身密度等因素均会影响环斑猛猎蝽的捕食能力。因此,需要进一步开展田间条件下环斑猛猎蝽对草地贪夜蛾的捕食功能研究,针对田间草地贪夜蛾的种群结构特点,确定适宜环斑猛猎蝽释放密度和时间,从而做到对草地贪夜蛾的有效防控。
