高原鼠兔干扰对退化草地植物经济类群和土壤理化性质的影响
2022-07-13李积兰卡着才让李希来
李积兰,卡着才让,李希来
(1.青海大学农牧学院,西宁 810016;2.青海省河南蒙古族自治县草原综合专业队,黄南 811599)
生态学中对干扰的定义是指自然或人为因素导致不连续的外部因素或间歇性因素的突然或连续影响[1]。干扰在自然界是普遍存在的现象,植物群落时刻受到干扰的影响[2]。干扰改变了生物生存的环境,导致生物不断适应环境的改变[3],植物群落内物种的地位和作用常随外界干扰而发生变化。青藏高原生态环境脆弱,对生物因子的干扰更加敏感[4,5],草地生态系统的生物干扰,尤其是食草动物的干扰是一个缓慢的过程[6]。草地生态系统中的食草动物除家畜外,小型哺乳动物种群也是其中的重要组分[7],高原鼠兔是青藏高原高寒草甸生态系统中最重要的小型哺乳动物之一[8],高原鼠兔活动干扰植物对环境资源的可利用率[9,10]。
高寒草地是青藏高原自然生态系统的重要组成部分及生态安全的屏障[10],长期以来人畜发展的失衡及高原鼠兔高密度的挖掘破坏,使得植物群落的次生演替速度及规模成为当前人类所面临的重要环境问题[10-12]。高寒草甸长期的过度放牧为草地啮齿动物的活动提供了有利的条件,从而加剧了草地退化的速度[13,14]。高原鼠兔主要通过采食牧草、挖掘洞穴、排泄粪便等活动方式,来改变地上地下的生物量及土壤理化性质[15]。有研究表明高原鼠兔干扰程度越大,其群落结构表现的越单一,土壤养分含量明显下降[16,17]。也有研究认为高原鼠兔活动改变了土壤的结构,加大了土壤的透气性能。近年来,有关高原鼠兔干扰对植物物种多样性的研究较多[18],而本文从高原鼠兔活动干扰程度设置人为控制试验,分析其对退化草地植物经济类群和土壤理化性质的影响,探究在其影响下植物经济类群和土壤理化性质的响应特征,为高寒草甸的合理利用提供基础数据。
1 材料与方法
1.1 研究区概况
研究区位于青海省黄南藏族自治州河南蒙古族自治县克其河滩,样区内过度放牧干扰程度高,植被明显退化,且具有秃斑地。试验地地理位置为东经101°47′、北纬34°44′,海拔约3 700 m。平均气温在-1.3~1.6 ℃,年降水量597.1~615.5 mm。该研究区主要草地类型为高寒草甸,主要植物为矮嵩草(Kobresiahumilis)、黄花棘豆(Oxytropisochrocephala)、针茅(Stipacapillifolia)、冰草(Agropyroncristatum)、火绒草(Leontopodiumleontopodioides)、线叶嵩草(Kobresiacapillifolia)。
1.2 实验设计
实验设计为单因素三水平实验处理,高原鼠兔数量水平分别为无鼠区(定期灭鼠)、中密度区(约75只/hm2)、高密度区(约150只/hm2)[19-21]。每个组合3次重复,共9个实验小区,每实验小区面积25 m×30 m,相邻小区在水平和垂直方向上都相隔5 m。为了防止高原鼠兔和其他食草动物进入样地,用网孔直径2.5 cm×2.5 cm,高50 cm的定制钢丝网围栏将地下部分封闭起来,高50 cm的防鼠铁皮围栏将地上部分封闭起来,钢纱网和镀锌铁皮各留10 cm用铆钉连接。铆钉密度需保证高原鼠兔不能从钢纱网和镀锌铁皮的缝隙中穿过。长1.4 m角铁用于固定围栏,间距2 m一个。为清点现有小区内的高原鼠兔,在每个小区内放置捕鼠器,将捕获的高原鼠兔进行数量和性别调查后按试验设计投放。对小区内的高原鼠兔按试验设计进行数量的补充与剔除。
1.3 样品采集及测定方法
采样时间为8月初,在每个25 m×30 m样地的对角线上,均匀设置3个1×1 m2的样方,在样方内调查植物的多度、盖度、高度及地下生物量,在同一样方内测定土壤紧实度、pH值,并采集0~10 cm、10~20 cm两层土壤,分别装入自封袋中并做好标记带回实验室,一部分土壤样品用于测定含水量,一部分土壤样品自然风干后测定每个样方的植物种类组成、植物高度、盖度、多度等。
盖度测定:目测法测定样方总盖度和各物种分盖度。
高度:样方中每种植物选取10株用钢卷尺测定其自然高度(不足10株的按实际株数测定)。
重要值=(相对盖度+相对高度+相对多度)/3(1-3)
1.4 样品测定
土壤紧实度用土壤紧实度仪(SC-900)测定;土壤含水量用烘干法测定;pH值用土壤原位pH计(IQ150)测定;土壤有机质利用重铬酸钾容量法进行分析;全氮用凯氏定氮法分析;全磷用酸溶-钼锑抗比色法测定;全钾、速效钾用火焰光度计法测定;速效氮用硫酸钠-纳氏试剂比色法测定;速效磷用Olsen法[22]。
1.5 数据整理与统计分析
运用Microsoft Excel 2019进行数据整理及表格制作,用IBM SPSS Statistics 20.0软件进行统计分析,采用单因素方差分析(One-way ANOVA)和Duncan’s法对植物经济类群重要值和土壤指标进行方差分析和多重比较,用Canoco 5.0进行冗余分析,并用向前筛选法和蒙特卡洛置换检验进行环境因子筛选和显著性检验[23]。数据结构采用算数平均值±标准差表示。
2 结果与分析
2.1不同高原鼠兔干扰强度对植物经济类群的影响
同一干扰处理下,4种植物经济类群的重要值为杂类草>禾草类>莎草类>毒草,杂类草与其他植物经济类群差异显著(P<0.05);禁鼠区,禾草类与莎草类、毒草差异显著(P<0.05),莎草类和毒草差异不显著(P>0.05);正常高原鼠兔区和高密度高原鼠兔区,禾草类、莎草类、毒草均差异不显著(P>0.05)。不同干扰处理下,随着高原鼠兔密度的增加,禾草类、莎草类、毒草均为先降低后增加的趋势,而杂类草为先增加后降低的趋势。同一植物类群重要值在不同干扰处理下差异不显著(P>0.05)(表1)。

表1 不同高原鼠兔干扰强度对植物经济类群重要值的影响
2.2不同高原鼠兔干扰强度对土壤理化性质的影响
由表2可知,随着高原鼠兔密度的增加,同一土层土壤全氮、全磷、速效磷、有机质、土壤含水量、土壤紧实度均呈现下降趋势,全钾、碱解氮含量先增加后降低,速效钾、pH值呈现增加的趋势。0~10 cm土层的土壤全钾含量、速效钾含量在高密度鼠兔区与禁鼠区差异显著(P<0.05),土壤有机质含量在禁鼠区与正常高原鼠兔区、高密度高原鼠兔区均差异显著(P<0.05),其他指标无显著性差异(P>0.05)。
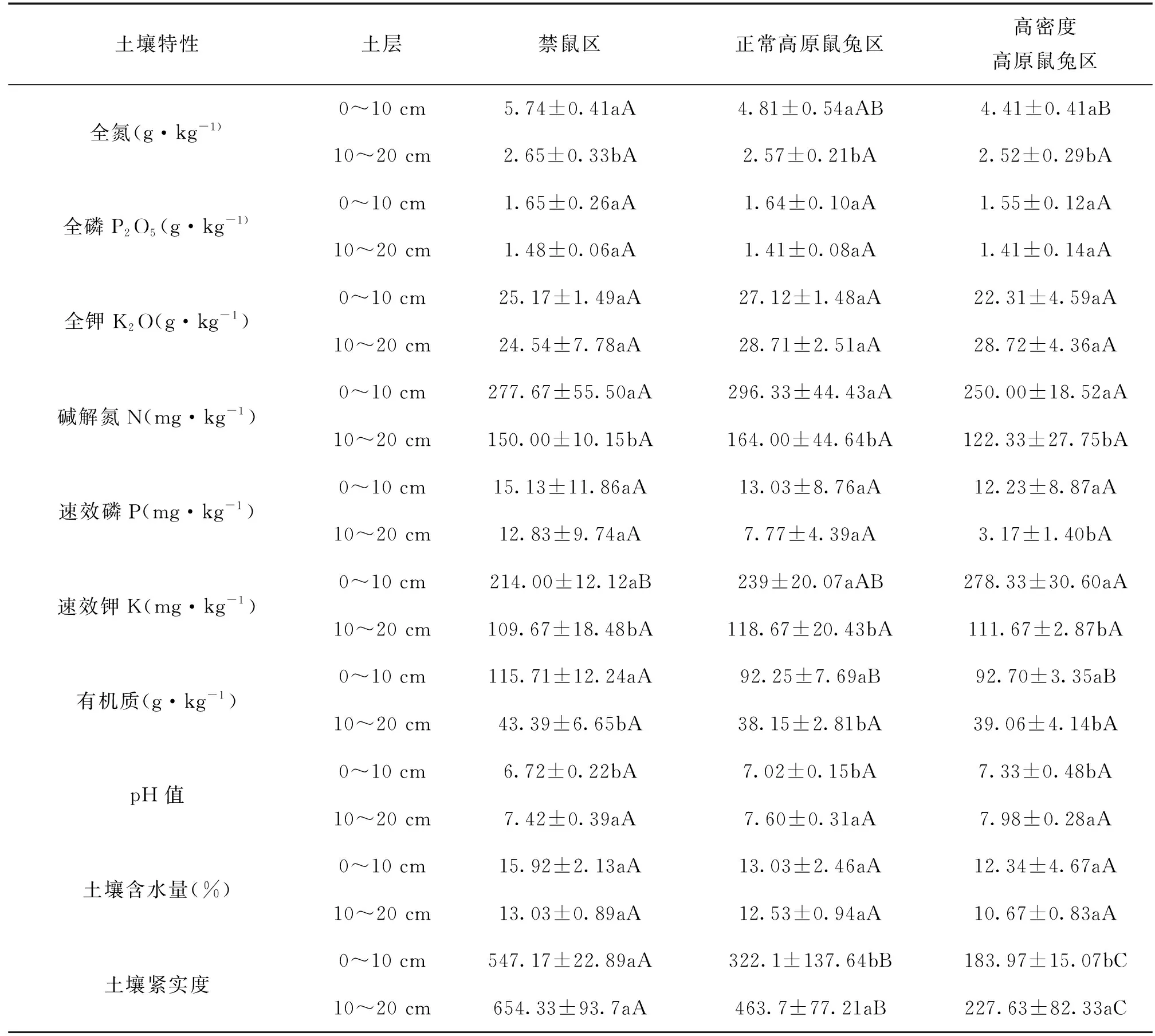
表2 不同高原鼠兔干扰强度下不同土层土壤理化性质
同一处理,随着土层深度的增加,除了土壤pH值增加外,其他指标均降低,且土壤全氮、碱解氮、速效磷(高密度高原鼠兔区)、有效钾、有机质、pH值、土壤紧实度(正常高原鼠兔区、高密度高原鼠兔区)在0~10 cm与10~20 cm土层中均差异显著(P<0.05),其他指标差异不显著(P>0.05)。
2.3 植物经济类群与土壤理化性质的关系
通过植物经济类群重要值与土壤理化性质的RDA冗余分析(图1),轴Ⅰ、轴Ⅱ特征值分别为0.6247、0.2229(图1a 和图1b 特征值一致),在0~10 cm土层(图1a)莎草与土壤碱解氮、pH值呈正相关,与土壤全磷、速效磷、全钾、含水量呈负相关;毒草、禾草与土壤有机质、全氮呈正相关;杂草与土壤全氮、有机质、全磷呈负相关。在10~20 cm土层(图1b),莎草与土壤速效钾、全钾呈正相关,与土壤速效磷呈负相关;毒草、禾草与土壤有机质、全氮、全磷、碱解氮呈正相关,与土壤pH值、全钾、速效磷呈负相关;杂草与土壤速效磷、pH值、全钾呈正相关,与土壤有机质、全氮、全磷、碱解氮呈负相关。RDA排序仅反映土壤环境因子与植物经济类群间存在相关性,难以量化环境因子对其分布的贡献程度。鉴于此,采用向前筛选法和蒙特卡洛检验对8个环境因子进行排序,由表3可知,0~10 cm土层环境因子对植物经济类群的解释力度由大到小依次为:有机质(差异显著,P<0.05)、全氮、pH值、全钾、碱解氮、含水量、速效磷、全磷。10~20 cm土层依次为有机质(差异显著,P<0.05)、全钾、速效钾、全磷、碱解氮、速效磷、pH值、全氮。
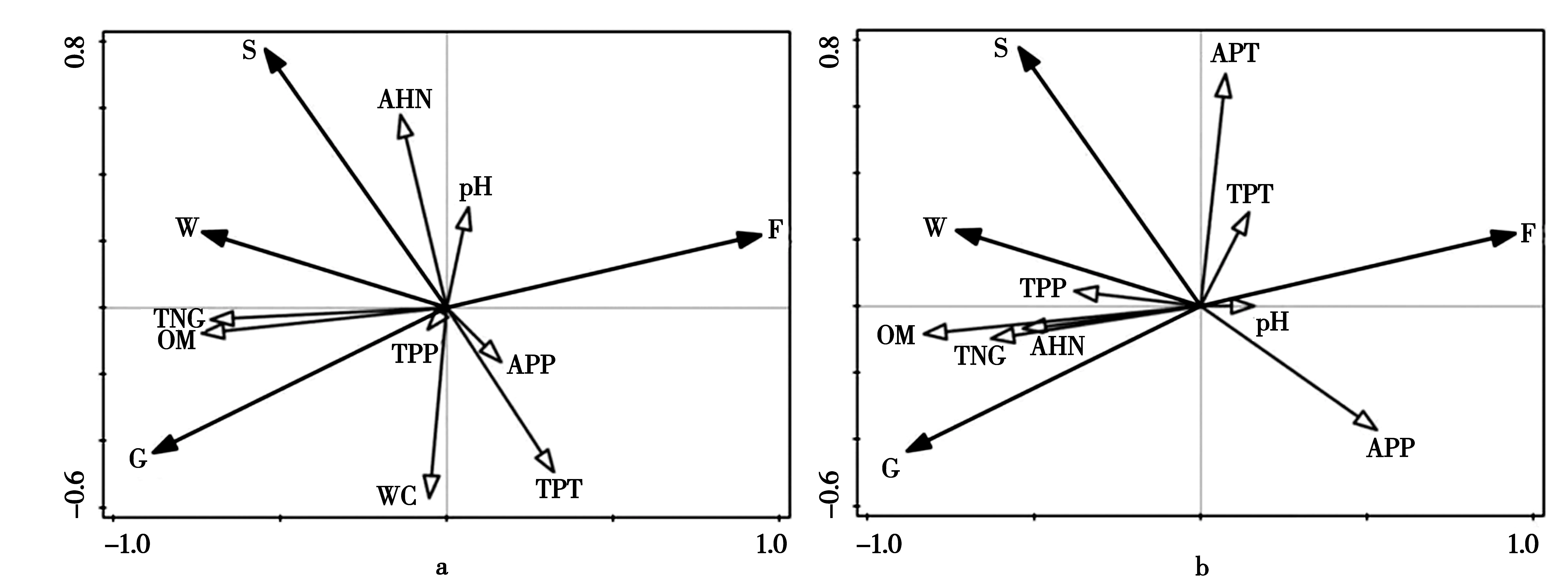
G:禾草;S:莎草;F:杂草;W:毒草;TNG:土壤全氮;TPP:土壤全磷;TPT:土壤全钾;AHN:土壤碱解氮;APP:土壤速效磷;APT:土壤速效钾;OM:土壤有机质;WC:土壤含水量;pH:土壤pH值。
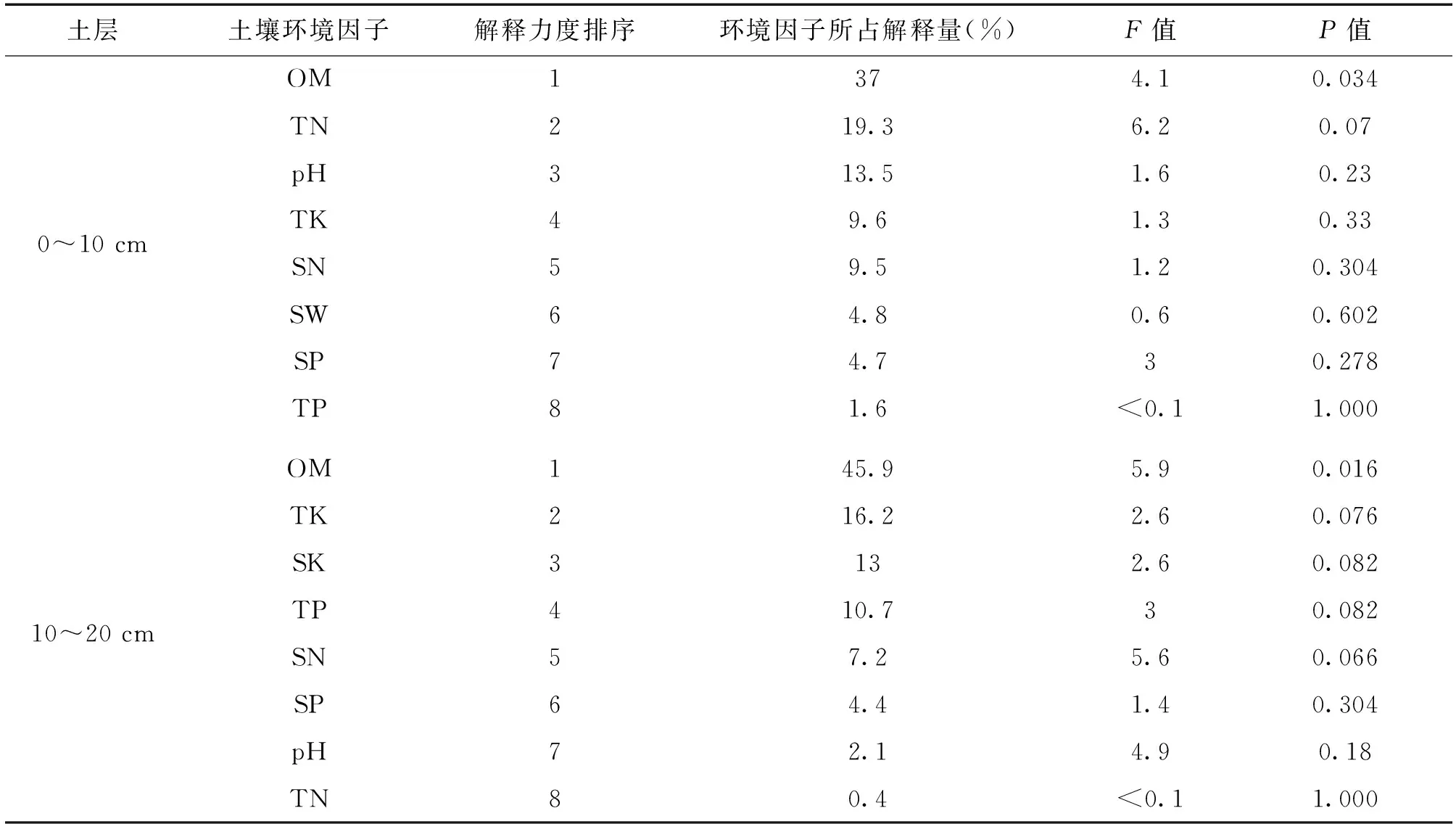
表3 不同土层土壤环境解释力度和显著性检验结果
3 讨论与结论
3.1高原鼠兔干扰对植物经济类群重要值的影响
高原鼠兔作为青藏高原高寒草甸生态系统内群居性的小型哺乳类动物,通过掘洞和采食行为干扰高寒草甸的生境,通过其干扰程度对植物和土壤产生影响[17],本研究发现随着高原鼠兔密度的增加,禾草类、莎草类、毒草的重要值均为先减少后增加的趋势,而杂类草重要值为先增加后减少的趋势。有研究认为,随着高原鼠兔干扰程度的增加莎草类植物的地上生物量会先增加后减少[24],禾草类优势种的重要值会先增加后减少,而毒杂草类会先减少后增加[17],鼠兔干扰过大会减少可食牧草比例[25]。本研究中的禾草、杂类草重要值变化趋势与已有研究结果不一致,原因可能是本实验测定数据为当年试验测定值,受干扰试验时间较短,因此高原鼠兔干扰对植物经济类群指标的影响需要考虑干扰时间。
3.2 高原鼠兔干扰对土壤理化性质的影响
高原鼠兔干扰过大时,会降低土壤养分含量[26],高原鼠兔干扰对土壤全氮、全磷有显著的影响(P<0.05),对速效磷、速效钾无显著影响(P<0.05),而在垂直分布上均有显著的影响[15],也有研究认为随高原鼠兔干扰程度的增加,土壤有机碳含量逐渐降低,全氮含量先升高后降低[8],适宜的高原鼠兔干扰能够增加土壤全磷含量,而过高或过低的高原鼠兔干扰均可降低土壤全磷含量,有机质的含量呈现出先增加后降低的趋势[17],土壤全碳、全氮、全磷和速效磷、速效钾贮量随高原鼠兔干扰强度增加均表现为先增加后降低的过程[27],土壤含水量、紧实度呈下降趋势[28]。本研究发现随着高原鼠兔密度的增加,同一土层土壤全氮、全磷、速效磷、有机质、土壤含水量、土壤紧实度均呈现下降趋势,全钾、碱解氮先增加后降低,速效钾、pH值呈现增加的趋势,这与以往高原鼠兔干扰降低速效钾的研究结论不一致,原因可能是随着高原鼠兔干扰密度的增加,植被草毡表层结构被破坏,从而影响土壤的通气性,加速土壤中速效钾的分解,短时间内使其含量增加。随着土层深度的增加,土壤全氮、碱解氮、速效磷(高密度高原鼠兔区)、有效钾、有机质、pH值、土壤紧实度(正常高原鼠兔区、高密度高原鼠兔区)在0~10 cm与10~20 cm土层中均差异显著(P<0.05),这与以往研究结果一致[15]。
3.3 植物经济类群重要值与土壤理化性质的关系
土壤与植被是一个整体系统,各种因子之间相互联系、共同作用,以往研究主要集中于植物群落多样性与土壤理化性质的相关性研究[17,18,24],而植物经济类群与土壤理化性质的相关性研究较少,本研究发现嵩草与10 cm土层土壤碱解氮、pH值,与20 cm土层土壤速效钾、全钾呈正相关;毒草、禾草与10 cm、20 cm土层土壤有机质、全氮均呈正相关;杂草与10 cm、20 cm土层土壤全氮、有机质、全磷均呈负相关。在不同高原鼠兔密度影响下,土壤理化性质对植物经济类群重要值影响的解释力度最大的为土壤有机质,说明土壤有机质是高原鼠兔干扰下植物经济类群分布的最关键因子。
