圆唇散白蚁工蚁生殖可塑性相关的性腺发育和基因表达
2022-07-13叶晨旭宋转转张文秀吴涛宇邢连喜苏晓红
叶晨旭, 宋转转, 张文秀, 吴涛宇, 刘 鹤, 邢连喜, 苏晓红,*
(1. 西北大学生命科学学院, 西安710069; 2. 陕西省珍稀濒危动物保育重点实验室, 西安710069)
社会性昆虫由于个体之间的分化和相互合作而构成了复杂的品级结构,“分工合作”是社会性昆虫在生态学上高度进化的一个重要因素。在所有的社会性昆虫中,白蚁具有最复杂和显著的品级分化。白蚁在卵期并不包含品级的某种特殊因素,品级分化现象是在卵期以后的发育过程中出现的。品级分化调节因素有3个方面:环境因素、作用于个体之间的信息素和内分泌调节因素(Elliott and Stay, 2008; Korb, 2015)。白蚁通常有兵蚁、工蚁和生殖蚁3个品级,品级的分化最终是由基因表达差异决定的(Yeetal., 2019);不同品级甚至在表皮结构和表皮蛋白相关基因表达都有显著差异,这些差异与品级的功能相适应(Suetal., 2016; Rasheedetal., 2019)。散白蚁属Reticulitermes卵孵化后经过2个龄期幼蚁发育途径向两个方向分化: 一个是若蚁途径(生殖型途径),长翅成虫就是由此途径产生并经过婚飞配对建立新巢成为原始蚁王和蚁后,也可形成翅芽型补充生殖蚁(brachypterous neotenic reproductives)和拟成虫型补充生殖蚁(adultoid neotenic reproductives);另一个是工蚁途径,工蚁发育有3种可能:维持工蚁、经过2次蜕皮成为兵蚁或成为无翅芽型补充生殖蚁(Elliott and Stay, 2008; Korb and Hartfelder, 2008; Korb, 2015; Suetal., 2015)。因此,仅有工蚁的蚁群可以自行产生补充生殖蚁(neotenic reproductives, NRs)和兵蚁发育为成熟蚁群。散白蚁雄性工蚁不需要转化成为形态明显的补充生殖蚁就能直接参与交配,而雌性工蚁经历2次蜕皮才能转化成为雌性补充生殖蚁(Fujita and Watanabe, 2010; Leniaudetal., 2011; Suetal., 2017)。已有的研究证实白蚁巢中的信息素诱导个体内保幼激素(juvenile hormone, JH)合成水平的动态变化,JH通过调控品级相关基因的表达促进工蚁向兵蚁和补充生殖蚁的转化(Korb and Hartfelder, 2008; Oguchietal., 2020)。
雌性工蚁向补充生殖蚁转化最主要的特征是性腺发育的恢复,然而工蚁卵巢的发育一直是不清楚的。已有的报道仅是表明工蚁的性腺发育受抑制,对工蚁生殖发育缺乏深入研究,尤其雌性工蚁必须经过蜕皮转化成补充生殖蚁之后才能启动生殖力,这个过程非常复杂。因此,需要对雌工蚁从低龄发育到老龄,以及老龄工蚁转化成补充生殖蚁这个过程中卵巢和卵母细胞发育过程进行完整研究,只有这样才能阐明工蚁生殖能力的改变是如何发生的。栖北散白Reticulitermessperatus雌性工蚁向补充生殖蚁转化卵巢长度约增加了4倍(Fujita and Watanabe, 2010);对尖唇散白蚁Reticulitermesaculabialis生殖蚁、工蚁和兵蚁卵子发生比较研究表明三者之间卵母细胞发育程度有显著性差异(朱蓉等, 2009; 王云霞等, 2011)。卵母细胞发育无疑是决定生殖力水平变化的关键因素,3龄雌性工蚁发育到6龄(老龄)工蚁,再转化为补充生殖蚁的过程中,卵巢以及生殖细胞发育是如何与其品级的功能相适应的目前还不清楚。
散白蚁属是世界上分布最广的白蚁类群之一,在生态稳定和社会经济中具有重要作用(Jongepieretal., 2018)。我国有白蚁4科44属476种,其中散白蚁属有111种(黄复生等, 2000; 程冬保和杨兆芬, 2014)。散白蚁种群扩张能力强与雌性工蚁向补充生殖蚁转化发育有直接关系。为了揭示雌性工蚁发育和生殖力改变过程中的性腺发育,我们研究了圆唇散白蚁Reticulitermeslabralis雌性工蚁从3龄-4龄-5龄-6龄(老龄)-转化成补充生殖蚁发育过程中的卵巢和卵母细胞动态变化;在圆唇散白蚁若蚁向3类生殖蚁转化的转录组中筛选出与卵母细胞生长期相关的基因,并利用这些基因在工蚁向补充生殖蚁转化发育过程中的表达变化,揭示工蚁向补充生殖蚁转化过程中卵母细胞从滞育到恢复生长发育的起点。
1 材料与方法
1.1 白蚁培养
实验所用的圆唇散白蚁R.labralis采自陕西省西安市,将带有蚁巢的木头带回实验室,放置于塑料培养箱内(80 cm×50 cm×40 cm)室温培养;培养箱中的巢群在实验中作为原巢,从原巢取得的工蚁称为原巢工蚁。6龄及6龄以上龄期的工蚁为老龄工蚁,圆唇散白蚁的老龄工蚁主要为6龄。从原巢中取老龄工蚁和兵蚁组成隔离群,每个隔离群由50头老龄工蚁和2头兵蚁组成,共设置100个隔离群,隔离群中的工蚁称为“隔离工蚁”。每个隔离群都饲养在铺有潮湿松木屑的玻璃培养皿中(直径90 mm)。每天观察和记录隔离工蚁发育情况。在隔离群中,约3周以后出现由雌性工蚁分化的前补充生殖蚁(pre-neotenic reproductives, pre-NRs),4周以后出现雌性无翅型补充生殖蚁。
1.2 工蚁以及工蚁向补充生殖蚁转化过程中的卵巢发育观察
在原巢中采集3-5龄和老龄工蚁;在隔离群中收集培养20 d的工蚁、前补充生殖蚁、转变2 d的补充生殖蚁和膨腹的补充生殖蚁。在PBS溶液中解剖个体取得完整卵巢,利用显微数码照相系统VHX-5000(基恩士公司,日本)拍照并测量每个卵巢的长度和宽度(n=10),卵巢长度包括卵原区、生长区和卵黄区,在卵巢最宽处测量的值计为卵巢宽度; 测量各发育时期卵巢管基部的卵母细胞长径用来表示卵母细胞大小(n=10)。卵巢的长度和宽度以及卵母细胞大小均用平均值±标准差表示。
1.3 工蚁向补充生殖蚁转化过程中的卵母细胞和滤泡细胞发育观察
将原巢雌性工蚁、在隔离群中培养20 d的雌性工蚁、前补充生殖蚁、转变2 d的补充雌性生殖蚁和膨腹的补充生殖蚁个体常规石蜡切片法切片,切片厚度7 μm,苏木精-伊红染色(haematoxylin and eosin staining)。利用数码显微照相系统观察卵母细胞发育和滤泡细胞形态,测量滤泡细胞层厚度(n=10), 滤泡细胞层厚度用平均值±标准差表示。
1.4 基因的筛选
末龄若蚁卵子发生处于卵母细胞生长期,经过一次蜕皮成为翅芽型补充生殖蚁、拟成虫型补充生殖蚁或有翅成虫之后, 卵母细胞开始进入卵黄形成期。雌性工蚁恢复生殖能力必须启动卵母细胞发育至末龄若蚁水平,具有与末龄若蚁相似的卵母细胞形态特征和卵母细胞发育相关基因表达模式。我们已经测定了圆唇散白蚁末龄若蚁、以及由末龄若蚁分化的翅芽型补充生殖蚁、拟成虫型补充生殖蚁和有翅成虫转录组(Suetal., 2016)。利用转录组数据库对若蚁和由末龄若蚁分化的生殖蚁的卵母细胞发育相关基因表达水平进行比较分析,筛选出在末龄若蚁中的表达水平显著高于转化初期生殖蚁的与卵母细胞生长期发育相关的基因,这些基因调控卵母细胞在生长期的发育。
1.5 qRT-PCR测定卵母细胞生长相关基因在工蚁向补充生殖蚁转化中的表达水平
1.5.1RNA提取及反转录:在液氮中分别研磨1.3节样品至粉末状,提取RNA;对RNA进行纯度检测合格后,进行下一步反转录。选取10 μL体系,RNA溶液8 μL,加入PrimeScriptTMRT Master Mix 2 μL,37℃反转录15 min,85℃反转录酶失活反应5 s,获得cDNA文库。
1.5.2引物的筛选:通过BLASTX软件设计所选基因的引物序列(表1),内参基因确定为β-action,配置5 μL梯度PCR实验反应体系: Master Mix 2.5 μL, 上下游引物(10 μmol/L)各0.2 μL, ddH2O 1.9 μL, cDNA 0.2 μL。梯度PCR温度范围为55±5℃,分为12个温度梯度。反应后通过琼脂糖凝胶电泳实验,筛选出可用引物,确定最适退火温度。
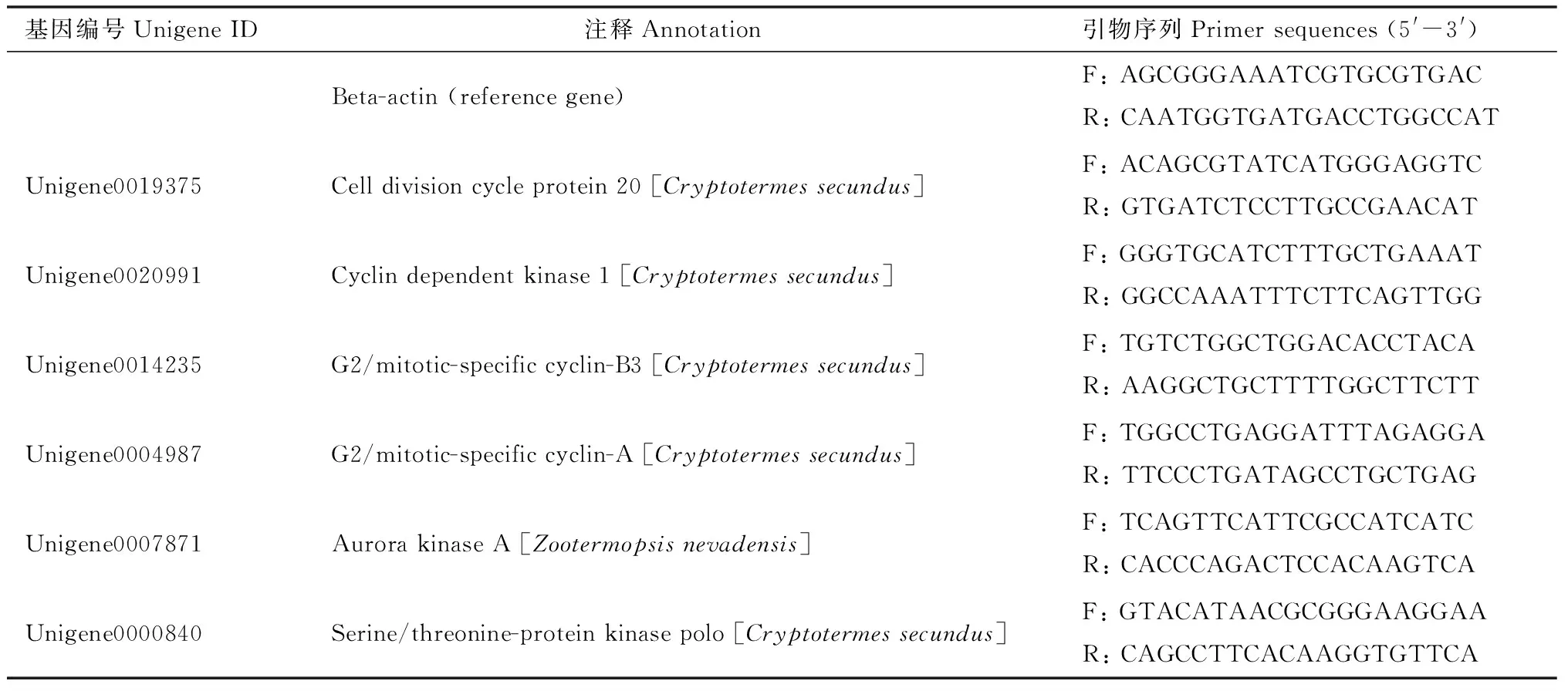
表1 qRT-PCR引物序列Table 1 Primer sequences for qRT-PCR
1.5.3qRT-PCR反应:选用Roche LightCycler®480荧光定量PCR反应仪器, PCR反应体系(20 μL): SYBR Premix Ex TaqTMⅡ 10 μL, 上下游引物(10 μmol/L)各0.8 μL, ROX 0.4 μL, ddH2O 6 μL, cDNA 2 μL。反应程序: 95℃ 30 s; 95℃ 5 s, 60℃ 30 s, 40个循环;融解曲线模式95℃ 5 s, 60℃ 1 min;降温50℃ 30 s。每样品有3个生物重复和3次技术重复。
1.6 数据分析
qRT-PCR反应的基因相对表达水平用 2-ΔΔCt方法计算。用R语言中的aov()函数对数据进行单因素方差分析,通过Tukey HSD()函数进行多重比较得到各样本均值差异的成对检测,P<0.05为显著性差异;最后用multcomp包中的glht()函数对结果进行可视化。
2 结果
2.1 工蚁向补充生殖蚁转化的形态学变化
圆唇散白蚁工蚁身体为浅麦黄色(图1: A);前补充生殖蚁(pre-NRs)身体为乳白色,腹部长度较工蚁的略长(图1: B),并且pre-NRs的第7腹片长度明显变长,覆盖于第8和9腹板上,与末龄若蚁的第7腹片形态非常相似,后缘为圆形,弧度非常明显。NRs身体深麦黄色,头顶上有纵的深棕色色素沉积带;中、后胸背板无翅或翅芽(图1: C)。刚变的NRs,腹部细长,从腹部前端向尾部逐渐变窄,第7腹板也呈明显的圆弧状并覆盖于第8和9腹板上。4周以后,NRs腹部开始明显膨胀,表明卵母细胞内卵黄积累形成(图1: D)。在实验过程中,隔离群内未发现有雄性NRs出现。
2.2 工蚁以及工蚁向补充生殖蚁转化过程中的卵巢发育
圆唇散白蚁工蚁从3龄(W3), 4龄(W4), 5龄(W5)到老龄工蚁(W6)发育过程中,随着龄期增加卵巢长度和宽度逐渐增加,每增加一个龄期,卵巢长度增加约100 μm;工蚁从3龄发育到老龄阶段卵巢长度增加了约1倍。虽然相邻龄期之间卵巢宽度没有显著性差异,卵巢宽度增加不显著,但是老龄工蚁W6卵巢宽度是W3的约1.5倍,从低龄向老龄发育过程中卵巢宽度在逐渐增加。隔离20 d的老龄工蚁与原巢老龄工蚁卵巢长度和宽度没有显著性差异,但是当隔离群中的雌性工蚁经过1次蜕皮转化成pre-NRs之后,卵巢极度变大。pre-NRs卵巢长度和宽度分别是工蚁的约2倍和3倍,卵巢长度增加近1 mm,宽度增加约150 μm。pre-NRs经过1次蜕皮转化成补充生殖蚁,NRs的卵巢与pre-NRs的卵巢相比,长度和宽度都没有显著增加。 NRs膨腹之后,虽然卵巢长度没有显著增加,但是宽度显著增加,膨腹补充生殖蚁(physogastric neotenic reproductives, phy-NRs) 卵巢宽度是刚变NRs的约2倍(图1: E-J; 图2)。
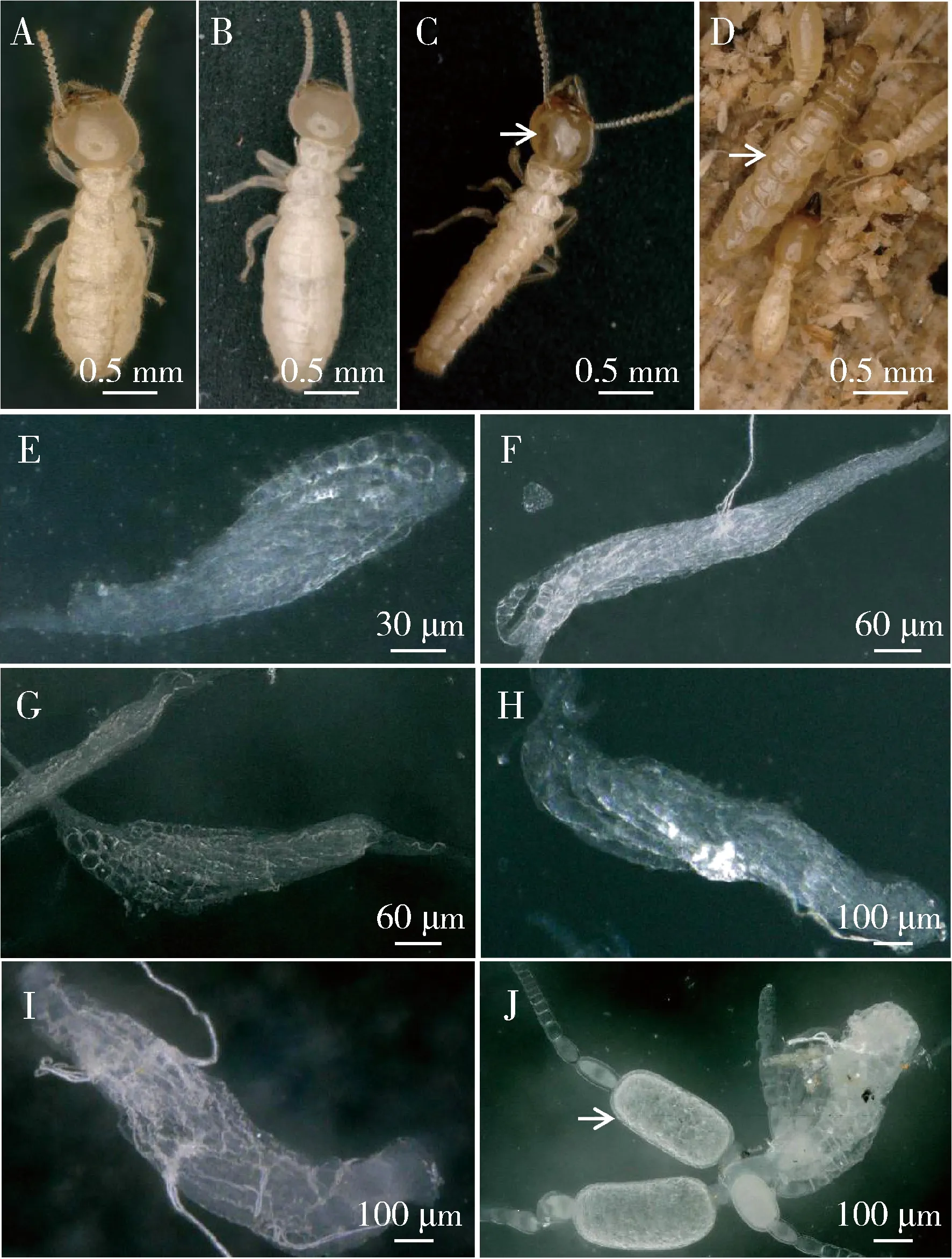
图1 圆唇散白蚁雌性工蚁、前补充生殖蚁和补充生殖蚁的形态Fig. 1 Morphology of the female workers, pre-neotenicreproductives and neotenic reproductivesof Reticulitermes labralisA: 工蚁Workers; B: 前补充生殖蚁Pre-neotenic reproductives; C: 补充生殖蚁身体细长,头部有深色色素条纹(箭头指示) Neotenic reproductives had the elongated abdomens and their heads had dark brown pigmentation stripes (indicated by the arrow); D: 膨腹补充生殖蚁(箭头指示)Physogastric neotenic reproductives (indicated by the arrow); E: 3龄工蚁的卵巢Ovary of the 3rd instar workers; F: 老龄工蚁的卵巢Ovary of the late instar workers; G: 隔离20 d工蚁的卵巢Ovary of isolated workers for 20 d; H: 前补充生殖蚁的卵巢Ovary of pre-neotenic reproductives; I: 补充生殖蚁的卵巢Ovary of neotenic reproductives; J: 膨腹补充生殖蚁卵巢有具卵黄的卵母细胞(箭头指示)Ovary of physogastric neotenic reproductives had vitellogenic oocytes (indicated by the arrow).

图2 圆唇散白蚁工蚁各龄期以及工蚁向补充生殖蚁转化过程中的卵巢发育Fig. 2 Ovarian development of workers at each instar andduring the differentiation of workers into neotenicreproductives of Reticulitermes labralis图数据为平均值±标准差;相同颜色柱子上不同字母表示有显著性差异(P<0.05, Tukey HSD检验)。Data in the figure are represented as mean±SD, and different letters above bars with the same colour indicate significant differences (P<0.05, Tukey HSD test). W3-6: 分别为3-6龄工蚁3rd-6th instar workers, respectively; IW: 隔离工蚁Isolated workers; pre-NR: 前补充生殖蚁Pre-neotenic reproductives; NR: 补充生殖蚁Neotenic reproductives; phy-NR: 膨腹补充生殖蚁Physogastric neotenic reproductives.
2.3 工蚁向补充生殖蚁转化过程中卵母细胞和滤泡细胞的发育
隔离20 d的圆唇散白蚁工蚁与原巢工蚁的卵母细胞和滤泡细胞发育水平没有显著性差异,它们的卵母细胞仍是在生长期,卵母细胞长径没有显著增加(P>0.05),滤泡细胞层比较薄(约1.20 μm)(图3: A, B)。隔离工蚁转化为pre-NRs之后,pre-NRs的卵母细胞和滤泡细胞层的发育水平仍然没有显著改变,卵母细胞停滞在生长期,卵母细胞长径和滤泡细胞层厚度也没有显著增加(P>0.05)(图3: C)。Pre-NRs蜕皮转化成NRs之后,卵母细胞虽然还处于生长期,但是卵母细胞长径和滤泡细胞层厚度显著增加(P<0.05),滤泡细胞为柱状形;NRs卵母细胞长径和滤泡细胞层厚度分别是Pre-NRs的约2倍和3倍(图3: D)。虽然末龄若蚁的卵母细胞也发育至生长期,滤泡细胞层厚度与NRs相比没有显著性差异(P>0.05),但是其卵母细胞长径是NRs的约2倍,刚转化的NRs卵母细胞发育程度低于末龄若蚁(图3: F)。随着NRs卵母细胞进入卵黄期,由于卵黄的积累,卵母细胞迅速增大,膨腹NRs卵母细胞长径是刚变NRs的约5倍(图3: E; 图4)。
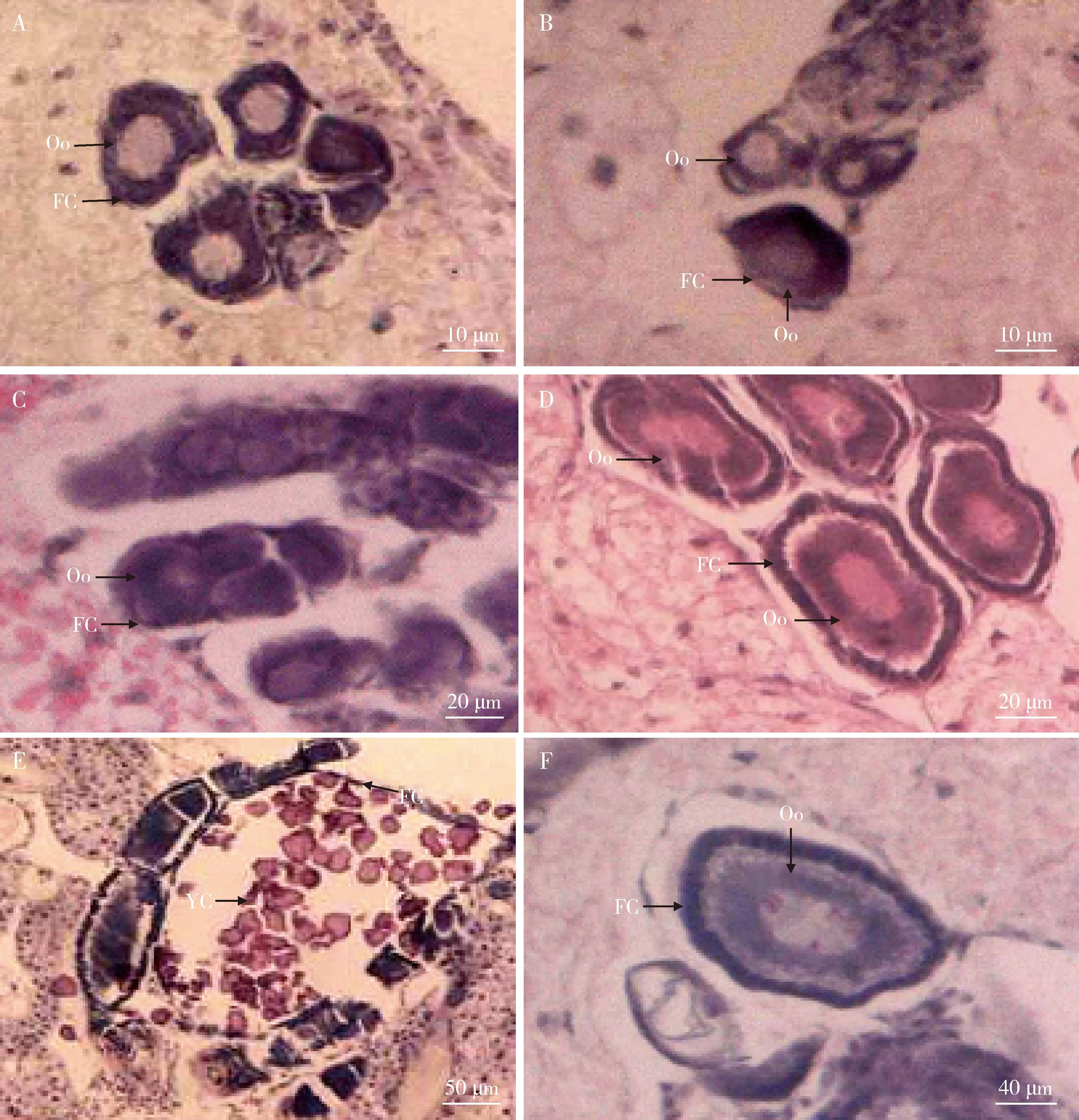
图3 圆唇散白蚁雌性工蚁向补充生殖蚁转化的卵母细胞发育Fig. 3 Oocyte development during the differentiation of female workers into neotenic reproductives of Reticulitermes labralisA: 在原巢工蚁中,卵母细胞被一层薄的滤泡细胞层包围In the workers in the parental nest, the oocytes were surrounded by a thin layer of follicle cells; B: 在隔离 20 d工蚁中,卵母细胞被一层薄的滤泡细胞层包围In the workers isolated for 20 d, the oocytes were surrounded by a thin layer of follicle cells; C: 在前补充生殖蚁中,卵母细胞被一层薄的滤泡细胞层包围In the pre-neotenic reproductives, the oocytes were surrounded by a thin layer of follicle cells; D: 在补充生殖蚁中,卵母细胞被一层厚的滤泡细胞层包围In the neotenic reproductives, the oocytes were surrounded by a thicker layer of follicle cells; E: 在膨腹补充生殖蚁中,卵黄发生末期的卵母细胞被一层薄的滤泡细胞层包围 In the physogastric neotenic reproductives, the oocytes during late vitellogenesis were surrounded by a thin layer of follicle cells; F: 末龄若蚁的卵母细胞被一层厚的滤泡细胞层包围 In the last instar nymphs, each oocyte was surrounded by a thicker layer of follicle cells. FC: 滤泡细胞层 Layer of follicle cells; Oo: 卵母细胞 Oocytes; YC: 卵黄Yolk.
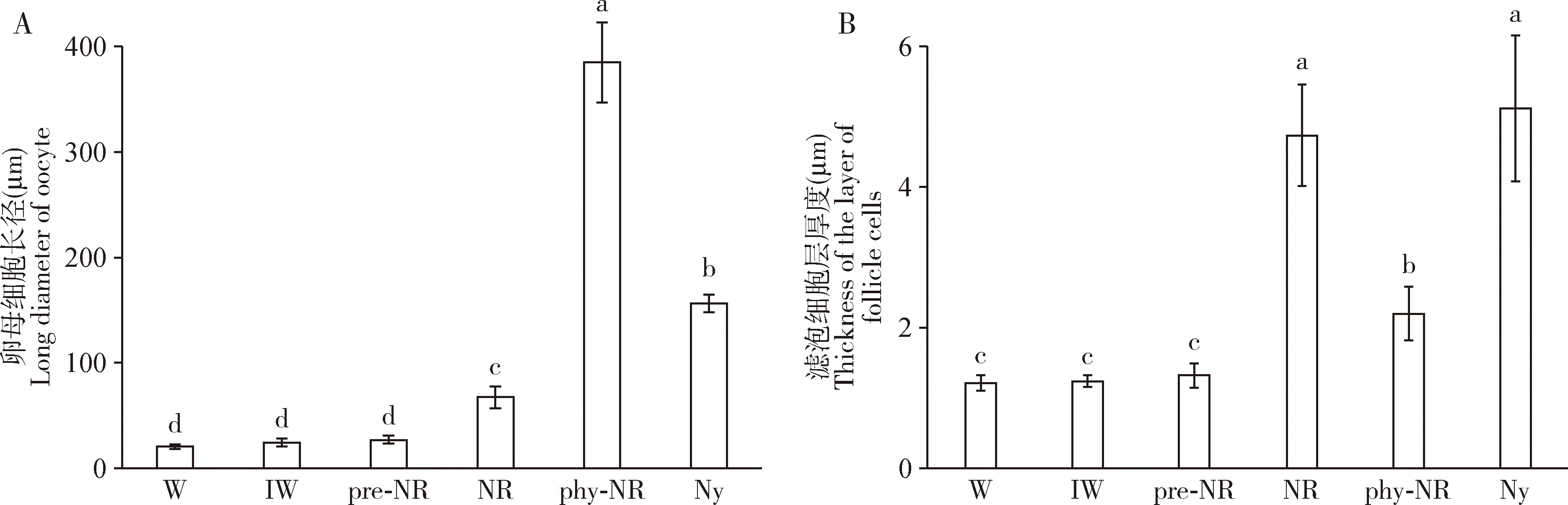
图4 圆唇散白蚁雌性工蚁向补充生殖蚁转化的卵母细胞大小(A)和滤泡细胞层厚度(B)变化Fig. 4 Changes in the size of oocytes (A) and the thickness of the layer of follicle cells (B) during the differentiationof female workers into neotenic reproductives of Reticulitermes labralis图数据为平均值±标准差;柱上不同字母表示有显著性差异(P<0.05, Tukey HSD检验)。Data in the figure are represented as mean±SD, and different letters above bars indicate significant differences (P<0.05, Tukey HSD test). W: 工蚁 Workers; IW: 隔离工蚁Isolated workers; pre-NR: 前补充生殖蚁Pre-neotenic reproductives; NR: 补充生殖蚁Neotenic reproductives; phy-NR: 膨腹补充生殖蚁Physogastric neotenic reproductives; Ny: 末龄若蚁Last instar nymphs.
2.4 调控卵母细胞生长期发育的相关基因
在我们的圆唇散白蚁雌性末龄若蚁向拟成虫型补充生殖蚁、翅芽型补充生殖蚁和有翅成虫转化的转录组数据库中(Suetal., 2016),根据末龄若蚁与这3种类型的生殖蚁的基因差异表达分析和卵母细胞发育调控通路筛选出与卵母细胞生长期发育相关的6个基因表达,它们是celldivisioncycleprotein20(Unigene0019375),cyclin-dependentkinase1(Unigene0020991),G2/mitotic-specificcyclin-B3(Unigene0014235),G2/mitotic-specificcyclin-A(Unigene0004987),aurorakinaseA(Unigene0007871)和serine/threonine-proteinkinasepolo(Unigene0000840)。AurorakinaseA(Unigene0007871)的序列与内华达古白蚁Zootermopsisnevadensis的相近,其余5个基因序列与堆砂白蚁Cryptotermessecundus的相近。这6个基因在雌性末龄若蚁的表达水平显著高于拟成虫型补充生殖蚁、翅芽型补充生殖蚁和有翅成虫(原始生殖蚁)的。末龄若蚁的卵母细胞处于生长期(即卵子发生的第2阶段), 末龄若蚁经过1次蜕皮分化为拟成虫型补充生殖蚁、翅芽型补充生殖蚁和有翅成虫之后,卵母细胞进入卵黄形成期(即卵子发生的第3阶段)。转录组数据显示这6个基因在末龄若蚁中高表达,而在3种类型的生殖蚁中相对表达水平比较低(图5),表明这些基因与卵母细胞生长期的发育有关。

图5 圆唇散白蚁6个与卵母细胞生长期发育相关的基因在末龄若蚁和生殖蚁中的差异表达热图Fig. 5 Heat map of the differential expression of six genesinvolved in oocyte development at the growth stage ofthe last instar nymphs and reproductives ofReticulitermes labralisLN: 末龄若蚁Last instar nymphs; AN: 拟成虫型补充生殖蚁Adultoid neotenic reproductives; BN: 翅芽型补充生殖蚁Brachypterous neotenic reproductives; AR: 有翅生殖蚁Alate reproductives.
2.5 与卵母细胞生长期发育相关基因在工蚁向补充生殖蚁转化过程中的表达
qRT-PCR的测定数据显示圆唇散白蚁工蚁经过pre-NRs转化为NRs之后,调控卵母细胞生长期发育的6个基因在NRs中的表达水平大幅度增加;虽然有3个基因表达水平在pre-NRs中显著高于在工蚁中的,但是增加幅度比较小。工蚁转化为pre-NRs之后,celldivisioncycleprotein20,cyclin-dependentkinase1和serine/threonine-proteinkinasepolo表达水平没有显著性增加;pre-NRs转化为NRs之后,表达水平显著性增加并且增加幅度很大,这3个基因在NRs的表达水平分别是pre-NRs中的约62, 34和106倍。G2/mitotic-specificcyclin-B3,G2/mitotic-specificcyclin-A和aurorakinaseA这3个基因在pre-NRs表达水平显著高于工蚁中的,分别是工蚁的约3, 3和2倍,增加幅度比较小;转化为NRs之后,这3个基因表达水平增加幅度很大,在NRs的表达水平分别是pre-NRs中的91, 36和57倍(图6)。

图6 qRT-PCR分析圆唇散白蚁卵母细胞生长期发育相关基因在工蚁、前补充生殖蚁和补充生殖蚁中的相对表达水平Fig. 6 Relative expression levels of genes involved in oocyte development at the growth stage in workers,pre-neotenic reproductives and neotenic reproductives of Reticulitermes labralis detected by qRT-PCRA: cell division cycle protein 20; B: cyclin-dependent kinase 1; C: G2/mitotic-specific cyclin-B3; D: G2/mitotic-specific cyclin-A; E: aurora kinase A; F: serine/threonine-protein kinase polo. 图中数据为平均值±标准差;柱上不同字母表示有显著性差异(P<0.05, TukeyHSD检验)。Data in the figure are represented as mean±SD, and different letters above bars indicate significant differences (P<0.05, Tukey HSD test). W: 工蚁Workers; pre-NR: 前补充生殖蚁Pre-neotenic reproductives; NR: 补充生殖蚁Neotenic reproductives.
3 讨论
我们研究发现工蚁从低龄到老龄的发育过程中卵巢没有停止生长,但是最终工蚁卵巢发育程度低于末龄若蚁,表明雌性工蚁性腺发育的停滞发生在老龄期。研究认为蚁后和蚁王利用信息素传递它们的生殖地位从而阻止工蚁性腺发育,只有蚁后和蚁王死亡或繁育能力不足的时候,工蚁才会转化成补充生殖蚁开始具备生殖能力(Korb, 2018)。蜜蜂工蜂发育的初期具有正常的卵巢,但在以后的发育中卵巢细胞程序化死亡,导致卵巢管数量从150~200条减少至几条,而且本质上是不育的。蜂后信息素抑制了工蜂卵巢发育,当缺乏蜂后抑制时,工蜂卵巢发育被激活(Capella and Hartfelder, 1998; Van Eeckhoven and Duncan, 2020)。山林原白蚁Hodotermopsissjostedi雄性补充生殖蚁的存在会加速雌性工蚁向补充生殖蚁转化(Oguchietal., 2020)。然而散白蚁的生殖策略比较复杂, 在栖北散白蚁R.speratus野外巢群中雌性补充生殖蚁的数量多于雄性补充生殖蚁,可能与雄性工蚁参与生殖有关(Fujita and Watanabe, 2010)。我们已有的研究也表明虽然圆唇散白蚁R.labralis雄性老龄工蚁精巢的直径显著小于生殖蚁精巢直径, 然而工蚁能够产生正常的精子(Suetal., 2015);雄性工蚁能够与雌性生殖交配并且提供正常的精子,表明雄性工蚁和雌性工蚁在生殖途径和生殖能力方面是不同的(Suetal., 2017)。雌性兵蚁个体终生不能生殖是雌性工蚁个体转化为兵蚁之后失去生殖可塑性的显著表现,经历2次蜕皮转变成兵蚁之后卵巢极度退化,这种退化是不可逆的。本研究发现刚转化的补充生殖蚁卵巢长度是工蚁卵巢长度的近3倍(图2),表明雌性工蚁生殖能力的恢复需要经历复杂的过程和调控。
卵母细胞发育恢复是生殖能力恢复的标志,我们的研究发现只有工蚁转化成补充生殖蚁之后,卵母细胞才能恢复发育,并且卵母细胞发育启动受滤泡细胞支持和调控。散白蚁生殖蚁卵巢管内卵子发生分为3个时期:卵母细胞分化期、卵母细胞生长期和卵母细胞卵黄形成期(朱蓉等, 2009)。虽然圆唇散白蚁工蚁和若蚁一样都发育到了卵母细胞生长期,但是若蚁卵母细胞直径是工蚁的约6倍,工蚁卵母细胞发育明显被抑制(图4: A),这种抑制与包围卵母细胞的滤泡细胞退化有关。当工蚁转化成补充生殖蚁之后,滤泡细胞层厚度会增加,卵母细胞变大。通常昆虫的卵母细胞移入生长区之后大量吸收营养物质,体积迅速增大;果蝇卵母细胞在生长期体积急剧增大是由于从滋养细胞获得了细胞质成分(Kolahietal., 2009)。果蝇卵子发生过程中滤泡细胞的屏障功能可以被重塑,滤泡细胞的形态和生理发生动态变化;在卵黄发生期滤泡细胞旁通透性发生改变,打开滤泡细胞旁通道让卵黄蛋白通过(Duhartetal., 2017; Isasti-Sanchezetal., 2021)。散白蚁卵母细胞被卵泡细胞所包围(无滋养细胞),其发育依赖滤泡细胞支持,显然工蚁卵母细胞发育的停滞和恢复受滤泡细胞调控。我们的研究表明滤泡细胞发育启动是非生殖品级向生殖品级转化生殖力开始恢复的主要标志。
工蚁隔离前和隔离后,卵巢长度和卵母细胞发育水平没有显著性变化,表明隔离工蚁必须经过蜕皮完成品级形态的转化才能恢复生殖能力。白蚁品级的转化是通过环境中的信息素改变个体内激素水平最终诱导基因表达来实现的;保幼激素以及保幼激素通路在低等白蚁品级非遗传多型性分化中起关键调控作用(Korb, 2015; Jongepieretal., 2018);用保幼激素类似物可以抑制雌性工蚁向生殖蚁分化,低水平保幼激素可以促进雌性工蚁向补充生殖蚁分化(Oguchietal., 2020)。近年还发现失去蚁后信息素抑制,工蚁的磷脂酰肌醇信号通路、钙信号通路、Ras信号通路、I3K-Akt 信号通路等信号通路相关基因显著上调,可能参与调控雌性工蚁向补充生殖蚁转化的信号传导(Yeetal., 2019)。我们的研究证实去掉蚁后抑制,工蚁的卵巢和卵母细胞并不能恢复发育,而是启动了向补充生殖蚁转化的调控通路。
我们对圆唇散白蚁末龄若蚁和末龄若蚁转化的3种类型生殖蚁的比较转录组进行了分析,发现了6个与卵母细胞第一次减数分裂前期相关的基因在末龄若蚁中表达水平显著高于刚转化的生殖蚁中的(图5和6),这可能是由于末龄若蚁生长期的卵母细胞正在启动和进行减数分裂的过程,而转化初期的生殖蚁卵母细胞准备进入卵黄积累阶段(卵子发生的最后阶段)。已有的研究表明cell division cycle 20(CDC20) 作为细胞分裂周期蛋白可以激活后期促进复合物/环小体(anaphase-promoting complex/cyclosome, APC/C),触发细胞分裂中期到后期的转变,启动姐妹染色单体分离(Jinetal., 2010; Lara-Gonzalezetal., 2019)。cyclin-dependent kinase 1作为细胞周期蛋白依赖激酶直接参与细胞周期调控,激发细胞周期各期的顺利进行(Shi and Feng, 2021);细胞周期蛋白 Cyclin 是调控真核生物细胞周期有丝分裂及减数分裂过程的一个十分重要的蛋白家族,G2/mitotic-specific cyclin-B3(Cyclin-B3) 是有丝分裂染色体分离启动的一个关键的调控因子(Garridoetal., 2020);G2/mitotic-specific cyclin-A(Cyclin -A)在DNA复制启动中起作用,也是DNA复制过程中核小体组装所必需的;在果蝇卵巢中调控生殖干细胞的发育和分化(Seimetal., 2003; Liuetal., 2017)。aurora kinase A对细胞中心体的成熟、纺锤体组装和染色体凝集过程有调节作用,是有丝分裂中必需的激酶,在动物卵母细胞成熟和激活过程中起着关键性的作用(Doyleetal., 2014; Nguyen and Schindler, 2017);serine/threonine-protein kinase polo (polo kinase)为polo激酶,与减数分裂恢复过程中的协调事件有关,对纺锤体形成有着重要的作用;敲除小鼠polo基因导致减数分裂在同源染色体分离之前停止,导致小鼠不能生育(Little and Jordan, 2020)。我们认为这些与末龄若蚁卵母细胞生长期发育相关的基因表达也可能与工蚁个体卵母细胞发育的抑制和启动有关系。
我们研究发现这6个基因在工蚁和前补充生殖蚁的表达水平较低,而当转化为补充生殖蚁之后全部高表达,表明卵母细胞发育的恢复和减数分裂启动从补充生殖蚁开始;而在末龄若蚁向生殖蚁转化的过程中,卵母细胞减数分裂启动从末龄若蚁开始。当卵母细胞进入生长期时,细胞核正处于第1次减数分裂的前期,卵母细胞合成和积累大量物质并进行DNA复制,这个时期对卵母细胞的成熟、受精和胚胎发育具有非常重要的意义。celldivisioncycle20表达水平低的雌性小鼠没有或很少生育后代,主要是由于卵母细胞减数分裂Ⅰ期染色体分离滞后和染色体错配所致(Jinetal., 2010);celldivisioncycle20在调节果蝇卵母细胞减数分裂中也很重要,突变可导致细胞有丝分裂中期和后期缺陷(Chuetal., 2001)。cyclin-dependentkinase1可以调控减数分裂的恢复和进程(Santamariaetal., 2007; Lietal., 2019);cyclin-B3作为一种有丝分裂周期蛋白在卵母细胞的减数分裂成熟过程中起作用;敲除cyclinB3基因的小鼠不育,用青蛙、斑马鱼和果蝇的cyclin B3可以使cyclin B3缺陷小鼠卵母细胞的减数分裂恢复(Guanetal., 2019; Karasuetal., 2019);G2/mitotic-specificcyclin-A在日本沼虾Macrobrachiumnipponense卵巢不同发育阶段的表达水平与卵巢成熟程度呈正相关,与卵原细胞的增殖和卵母细胞的形成有关(Zhouetal., 2021);aurorakinaseA在有丝分裂由G2进入M期形成纺锤体后表达水平增高达到峰值,在动物卵母细胞成熟和激活过程中起着关键性的作用(Dutertreetal., 2002);serine/threonine-proteinkinasepolo在G2期和M期表达水平较高,表达水平低时会导致卵母细胞减数分裂的阻滞(Little and Jordan, 2020)。工蚁转化的补充生殖蚁与若蚁转化的补充生殖蚁不仅外部形态有显著差异,而且卵母细胞发育水平不同,若蚁转化的补充生殖蚁可以直接进入卵子发生的最后一个阶段即卵黄形成期,而工蚁转化的补充生殖蚁卵子发生将恢复第二阶段的发育即卵母细胞生长期。因此,可以推测若蚁转化的补充生殖蚁比工蚁转化的补充生殖蚁产卵要早。虽然Cyclin-B3,Cyclin-A和aurorakinaseA在前补充生殖蚁的表达水平显著高于工蚁中的,但是表达水平的增高程度与补充生殖蚁中的相比还很低;另外,工蚁蜕皮转化成前补充生殖蚁之后,卵母细胞和滤泡细胞形态结构没有显著改变。因此,这6个基因在工蚁、前补充生殖蚁和补充生殖蚁表达水平比较明确了雌性工蚁向补充生殖蚁转化过程中卵母细胞发育的恢复是从转化为补充生殖蚁开始的。
