采前红蓝光连续光照光强对水培生菜生长、品质及AsA代谢的影响
2022-07-13刘家源张玉彬刘文科
刘家源, 张玉彬, 刘文科
(中国农业科学院农业环境与可持续发展研究所,农业农村部设施农业节能与废弃物处理重点实验室,北京 100081)
抗坏血酸(ascorbic acid,AsA)又称维生素C, 存在于叶绿体、线粒体、液泡、细胞质和细胞壁中,在叶绿体中浓度最高[1]。AsA与其他抗氧化剂共同组成抗氧化系统,保护植物免受有氧代谢、光合作用和多种污染物造成的氧化损害[2]。内源性AsA水平的高低会影响植物开花、衰老相关基因的诱导,与细胞的凋亡、病原体的信号转导和植物体衰老的发展都有联系[3]。维持体内AsA在临界水平之上对人体健康至关重要,缺乏AsA会引起坏血病等疾病。但是人体缺乏催化AsA合成的酶,并且相关基因在进化过程中累积了有害突变[4-6],因此人体自身不能合成AsA,必需通过不断的膳食摄取才能保证人体健康。植物是人类饮食中最主要和最重要的AsA来源,提高植物组织的AsA含量对于维持和促进人类健康具有重要意义。
连续光照通常是指改变植物原有的光周期规律,给植物提供连续24 h的光照[7]。连续光照必需通过人工光源才能实现,因而连续光照是设施园艺中特有的光照模式。连续光照打破了植物原有的生长规律,对于植物而言是一种光胁迫。目前对于连续光照的研究主要是采用人工补光的方式来改变植物原有的明暗交替的光周期规律生长方式,研究其对植物生长发育的影响[8]。周晚来[9]研究发现,在生菜采收前,进行短期连续光照可以显著降低水培生菜(Lactuca sativa L.)的硝酸盐含量,并提高可溶性糖、AsA等营养物质的含量;Ohyama等[10]研究发现,连续光照促使番茄嫁接苗植株鲜重、干重及叶面积等显著提高,且未对番茄叶片造成伤害。事实上,已有大量研究表明,在一定范围内,光强越大,植物体内的 AsA 含量越高[11]。Massot等[12]对番茄植株进行梯度光照强度处理后发现,植株AsA的总量随光照强度的降低而减少,说明这种现象受到了光照强度的调控,对植株中AsA的合成代谢起到一定的调节作用[13]。
生菜是一种广泛食用的全球性叶菜类蔬菜,也是人工光植物工厂广泛种植的代表性蔬菜。高浓度硝态氮的栽培环境是提高水培叶菜产量的有效方法,但在实际生产中,植物工厂水培叶菜高产的同时品质低的问题也十分突出,主要表现为以抗坏血酸为代表的次生代谢物质和其他基础营养物质含量低。截至目前,光照强度对AsA代谢的调控机制仍不清楚。因此,本研究拟在作物生长过程中,通过研究采前LED连续光照的光照强度对生菜AsA等营养物质的含量及AsA相关合成代谢酶活性的影响,探索生菜体内AsA等营养物质的含量及AsA代谢网络对采收前连续光照光强的响应机理。
1 材料与方法
1.1 试验材料
本试验在中国农业科学院农业环境与可持续发展研究所植物工厂进行。试验采取水培方式,种植环境温度为(25±1)℃,相对湿度为65%±5%,CO2浓度为大气CO2浓度。试验用生菜品种为意大利耐抽薹,由本课题组保存。
1.2 试验设计
将生菜种子播于海绵块中育苗,培养15 d后,将长势一致的生菜苗随机移栽于长方形塑料栽培槽(长180 cm×宽60 cm×高6 cm)内,并于次日开始光照试验。试验期间采用营养液水培,营养液采用霍格兰配方。
试验前期正常光照采用LED红蓝光面板灯进行光照处理,红蓝光比例为4∶1,光照强度为150 μmol·m-2·s-1,光周期为16 h/8 h(光期时间段:6:00—22:00),此种光环境设置可以更好地促进生菜产量和品质的提高[14]。植物工厂生菜因品种不同,生长速度也有差异。根据前期试验得到的结果,本试验统一连续培养17 d,生菜达到采收标准后进行72 h的连续光照处理,从定植后第17天的6:00开始连续光照,在第20天6:00光照结束。每个处理种植生菜26株,采用红光波峰为655 nm,蓝光波峰为430 nm的LED红蓝光灯板(49 cm×49 cm)进行光照处理。灯板悬挂于栽培槽上方40 cm处。集中连续光照强度分别为100、150、200、300和500 μmol·m-2·s-1(分别用CL100、CL150、CL200、CL300和CL500表示),以延续此前 150 μmol·m-2·s-1、16 h/8 h 光周期的光照条件为对照(用CK表示)。
1.3 测定项目及方法
在连续光照开始和结束时取样,采用SPAD-502叶绿素含量测定仪(Imaging-PAM,德国Walz)测定叶绿素含量(SPAD)。分别于每个处理中随机选8株生菜作为重复样本,从茎基部切开,其中4株的地上部分将叶片与叶柄分离后,迅速用液氮冷冻,并用高通量组织研磨器在低温下把用液氮冷冻好的植物样品研磨成粉末,放至-80℃冰箱中留样备用。另外4株用电子计数天平称取地上部鲜重和根鲜重,用Li-3100C叶面积仪(美国LI-COR)测量整株生菜叶片的叶面积,然后将生菜100℃下杀青20 min,80℃烘干至恒重,用分析天平分别称取地上部干重和根干重。生理指标和品质指标测定的取样叶位为同叶位。
采用便携式光合仪(LI-6400XT,美国)分别在采前连续光照前后分2次测定生菜不同处理下叶片的净光合速率(net photosynthetic rate,Pn)、蒸腾速率(transpiration rate,Tr)、胞间CO2浓度(intercellular CO2concentration,Ci)和气孔导度(stomatalconductance,Gs)。
硝酸盐含量测定采用硫酸-水杨酸法[15]测定,取新鲜植物样品粉末0.1 g,加入1.5 mL蒸馏水,放入沸水浴中提取。取0.1 mL浸提液至10 mL试管中,加入0.4 mL 5%的水杨酸-浓硫酸溶液,混匀、冷却后后加入9.5 mL 8%的NaOH溶液,冷却后测定并计算硝酸盐的含量。
可溶性糖含量测定采用硫酸-苯酚法,参考李合生[16]的试验方法,取新鲜植物样品粉末0.1 g,加入1.5 mL蒸馏水,放入沸水浴中提取。然后吸取0.5 mL样品液于试管中,加蒸馏水1.5 mL,并以此加入苯酚、浓硫酸溶液、显色,测定并计算可溶性糖的含量。
参照Spinola等[17]的方法,采用 UPLC 测定抗坏血酸(ASA)含量。提取液:1.5% MPA,4% CH3COOH, 0.5mmol·L-1EDTA;缓 冲 液 :200 mmol·L-1Tris;还原液:750 mmol·L-1DTT;反应停止液:0.4 mol·L-1H2SO4。色谱柱为 HSS T3 2.1 mm×50 mm,1.8 μm,流动相为 0.1% 的甲酸溶液(超纯水配制),流速0.25 mL·min-1,进样量2 μL,柱温25℃,样品室温度10℃,检测波长245 nm,运行时间1.5 min。DHA含量、L-半乳糖酸 -1,4-内 酯 脱 氢 酶(L-galactono-1,4-lactone dehydro-genase,GalLDH)酶活性、AsA过氧化物酶(APX)活性、AsA还原酶(DHAR)活性、单脱氢AsA还原酶(MDHAR)活性和谷胱甘肽还原酶(GR)活性的测定参照文献[18]进行。
1.4 数据处理
分别用Microsoft Excel 2010和SPSS 25.0统计分析软件对数据进行分析和差异显著性检验。
2 结果与分析
2.1 采前LED连续光照光强对生菜生长的影响
2.1.1 采前LED连续光照光强对生菜产量的影响 从图1可以看出,采前LED连续光照光强对生菜产量具有显著影响。非连续光照处理72 h(CK)后,生菜植株地上部分鲜重提升了2.65 g;而连续光照处理72 h后,生菜植株地上部分鲜重显著提高,CL100、CL150、CL200、CL300和 CL500处理中,72 h后干重都出现显著增加。非连续光照处理72 h(CK)后,生菜植株根鲜重提升了0.60 g;在连续光照处理72 h后,生菜植株根鲜重显著提高,CL150、CL200、CL300和 CL500处理中,72 h后干重均出现提升。非连续光照处理72 h(CK)后,生菜植株地上部干重增加了0.14 g;而连续光照处理72 h后,高强度光照处理中地上部干重显著提高,CL150、CL200、CL300和CL500处理中,72 h后地上部干重均显著提升。非连续光照处理72 h(CK)后,生菜根干重增加了0.014 g;而连续光照处理72 h后,高强度光照处理中地上部干重明显增加,CL200、CL300和CL500处理中,72 h后根干重分别增加了0.11、0.12和0.24 g。结果表明,随着采收前最后72 h LED红蓝光连续照射的光照强度升高,生菜地上部的干鲜重和根的干鲜重都升高。
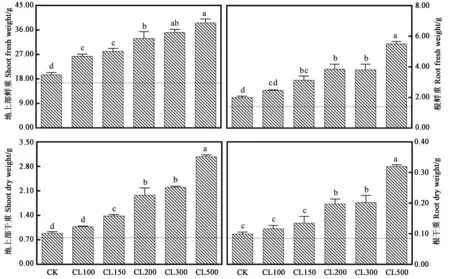
图1 采前LED连续光照光强对生菜产量的影响Fig.1 Effects of LED continuous light intensity on lettuces yield before harvest
2.1.2 采前LED连续光照光强对生菜形态指标的影响 图2显示,非连续光照处理72 h(CK)后,生菜叶面积增加了74 cm2;而连续光照处理72 h后,生菜叶面积显著提高,CL150、CL200、CL300和CL500处理中,72 h后干物质含量均出现增加。非连续光照处理72 h(CK)后,生菜叶片的SPAD略微增加至19.83;而高光强连续光照处理72 h后,生菜叶片SPAD明显增加。非连续光照处理72 h(CK)后,生菜干物质含量无变化;而高光强连续光照处理72 h后,生菜干物质含量明显增加。高光强连续光照处理72 h后,生菜比叶重明显增加。可见,随着采前LED连续光照光强的增加,生菜叶面积、干物质含量和比叶重均显著增加,SPAD在CL200处理时达到最高,之后不再随CL光强的增加而增加。
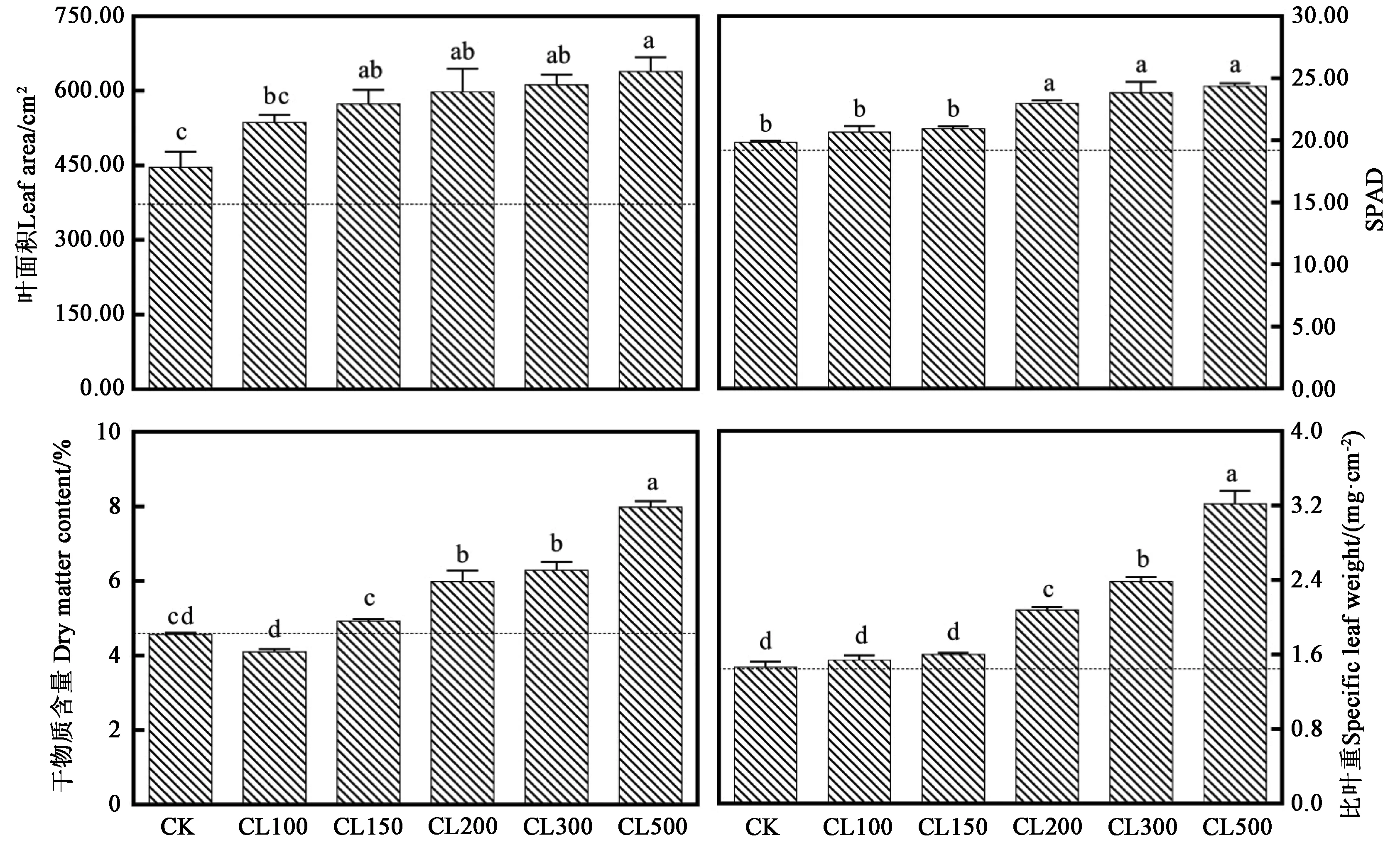
图2 采前LED连续光照光强对生菜形态指标的影响Fig.2 Effects of LED continuous lighting intensity on morphological index of lettuce before harvest
2.2 采前LED连续光照光强对生菜光合特性的影响
图3显示,采前LED连续光照光强对生菜光合特性有一定的影响。非连续光照处理72 h(CK)后,生菜叶片的净光合速率(Pn)增加了1.51 μmol·m-2·s-1;而高光强连续光照处理72 h后,生菜叶片的Pn明显增加。非连续光照处理72 h(CK)后,生菜叶片的气孔导度(Gs)略微降低;而进行连续光照处理72 h后,CL300和CL500处理中生菜叶片的Gs明显增加;CL100和CL150处理中生菜叶片的Gs明显降低,72 h 后 Gs分别降低了 0.102 和 0.108 mol·m-2·s-1。非连续光照处理72 h(CK)后,生菜叶片的胞间CO2浓度(Ci)略微降低;而连续光照处理72 h后,CL300处理中生菜叶片的Ci明显增加;CL150和CL200处理中生菜叶片的Ci均明显降低。非连续光照处理72 h(CK)后,生菜叶片的蒸腾速率(Tr)略微升高;而连续光照处理72 h后,CL300和CL500处理中生菜叶片的Tr明显增加。
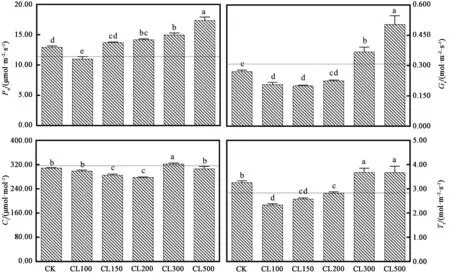
图3 采前LED连续光照光强对生菜光合特性的影响Fig.3 Effects of LED continuous light intensity on photosynthetic characteristics of lettuces
2.3 采前LED连续光照光强对生菜品质的影响
由图4可知,非连续光照处理72 h(CK)后,生菜叶片的可溶性糖含量略微升高;而连续光照处理72 h后,CL150、CL200、CL300和CL500处理中生菜叶片的可溶性糖含量明显增加。非连续光照处理72 h(CK)后,生菜叶柄的可溶性糖含量升高了4.80 mg·g-1;而连续光照处理72 h后,CL200和CL500处理中生菜叶柄的可溶性糖含量明显增加,72 h后分别增加了19.20和33.70 mg·g-1。非连续光照处理72 h(CK)后,生菜叶片的硝酸盐含量略微升高;而连续光照处理72 h后,CL200、CL300和CL500处理中生菜叶片的硝酸盐含量明显降低。非连续光照处理72 h(CK)后,生菜叶柄的硝酸盐含量升高了17.46 mg·kg-1;而连续光照处理72 h后,CL200和CL500处理中生菜叶柄的硝酸盐含量明显增加,72 h后分别增加了116.54和171.54 mg·kg-1。可见,生菜叶片和叶柄的可溶性糖含量随CL光强的升高而升高,生菜叶片和叶柄的硝酸盐含量随CL光强的升高而降低。

图4 采前LED连续光照光强对生菜可溶性糖和硝酸盐含量的影响Fig.4 Effects of LED continuous lighting intensity on soluble sugar and nitrate contents of lettuces before harvest
2.4 采前LED连续光照光强对生菜AsA代谢的影响
为探明LED红蓝光连续光照对水培生菜中AsA含量的调控效果及其相应的代谢机理。本研究以AsA代谢途径(合成、氧化和还原再生)连续光照响应位点与联动机制为突破口,在基因表达和关键酶活性层面研究了红蓝光连续光照对水培生菜AsA代谢关键酶(GalLDH、APX、MDHAR、DHAR和GR)的影响及其与时长、光强和光质的关系,揭示水培生菜AsA代谢对连续光照响应机理。
2.4.1 采前LED连续光照光强对生菜AsA和DHA含量的影响 由图5可知,非连续光照处理72 h(CK)后,生菜叶片的AsA含量略微升高;而连续光照处理72 h后,CL200、CL300和CL500处理中生菜叶片的AsA含量明显增加。非连续光照处理72 h(CK)后,生菜叶柄的AsA含量升高了0.21 mg·g-1;而连续光照处理72 h后,各处理的生菜叶柄中AsA含量均无显著差异。非连续光照处理72 h(CK)后,生菜叶片的DHA含量略微升高;而CL150处理中生菜叶片的DHA含量明显增加,72 h后增加了0.32 mg·g-1。非连续光照处理72 h(CK)后,生菜叶柄的DHA含量略微降低;而连续光照处理72 h后,各处理的生菜叶柄中DHA含量均无显著差异。可见,采前LED连续光照处理后,随着CL光强的升高,生菜叶片AsA含量逐渐升高,生菜叶片DHA含量有先升高后降低的趋势,各处理中生菜叶柄的AsA和DHA含量均无显著差异。从图5中可以明显看出,各处理中,生菜叶片的AsA含量显著高于叶柄的AsA含量。
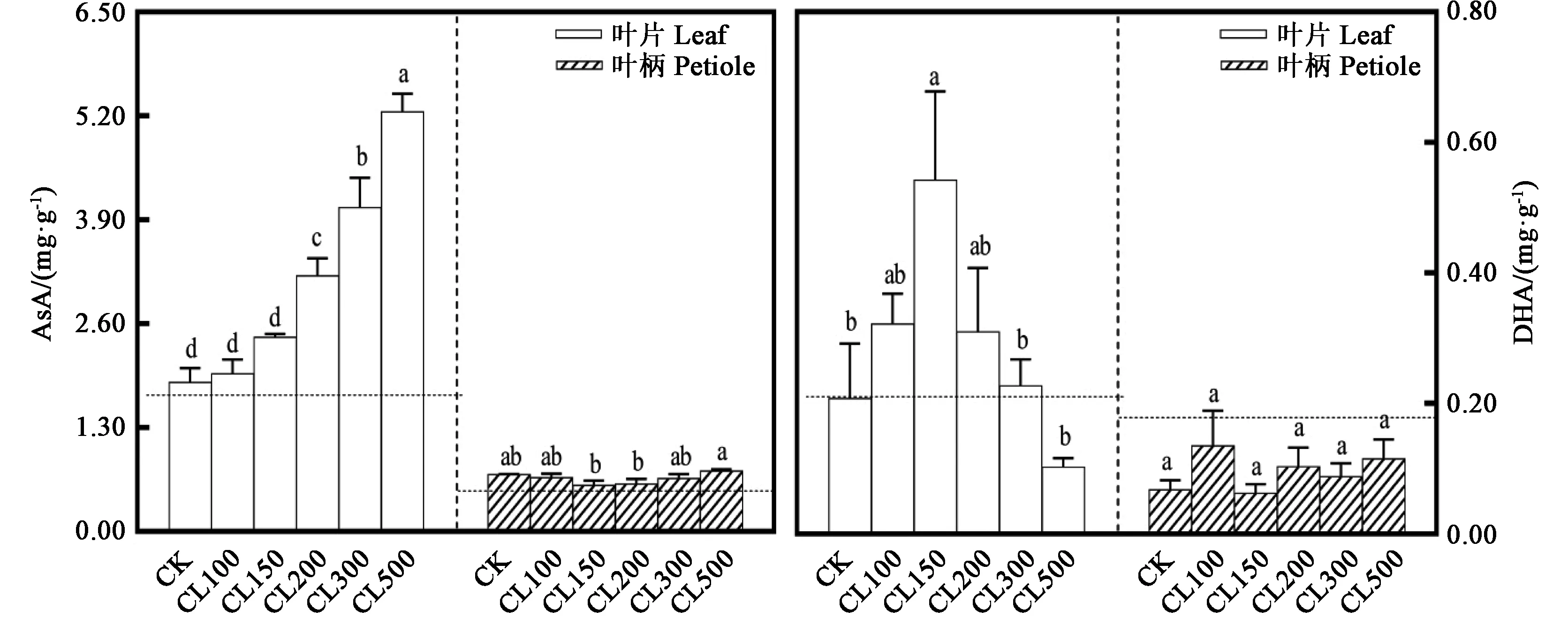
图5 采前LED连续光照光强对生菜AsA和DHA含量的影响Fig.5 Effects of LED continuous light intensity on ASA and DHA contents of lettuces before harvest
2.4.2 采前LED连续光照光强对生菜GalLDH酶活性的影响 由图6可知,非连续光照处理72 h(CK)后,生菜叶片的GalLDH酶活性略微升高;而连续光照处理72 h后,CL100处理中生菜叶片的GalLDH酶活性最高,达到0.338 U·mg-1FW;CL500处理中生菜叶片的GalLDH酶活性明显下降。非连续光照处理72 h(CK)后,生菜叶柄的GalLDH酶活性略微升高;而连续光照处理72 h后,各处理的生菜叶柄中GalLDH酶活性均无显著差异。可以看出,72 h采前LED连续光照后,生菜叶片的GalLDH酶活性随CL光强的升高而逐渐下降,各处理的生菜叶柄中GalLDH酶活性均无显著差异。
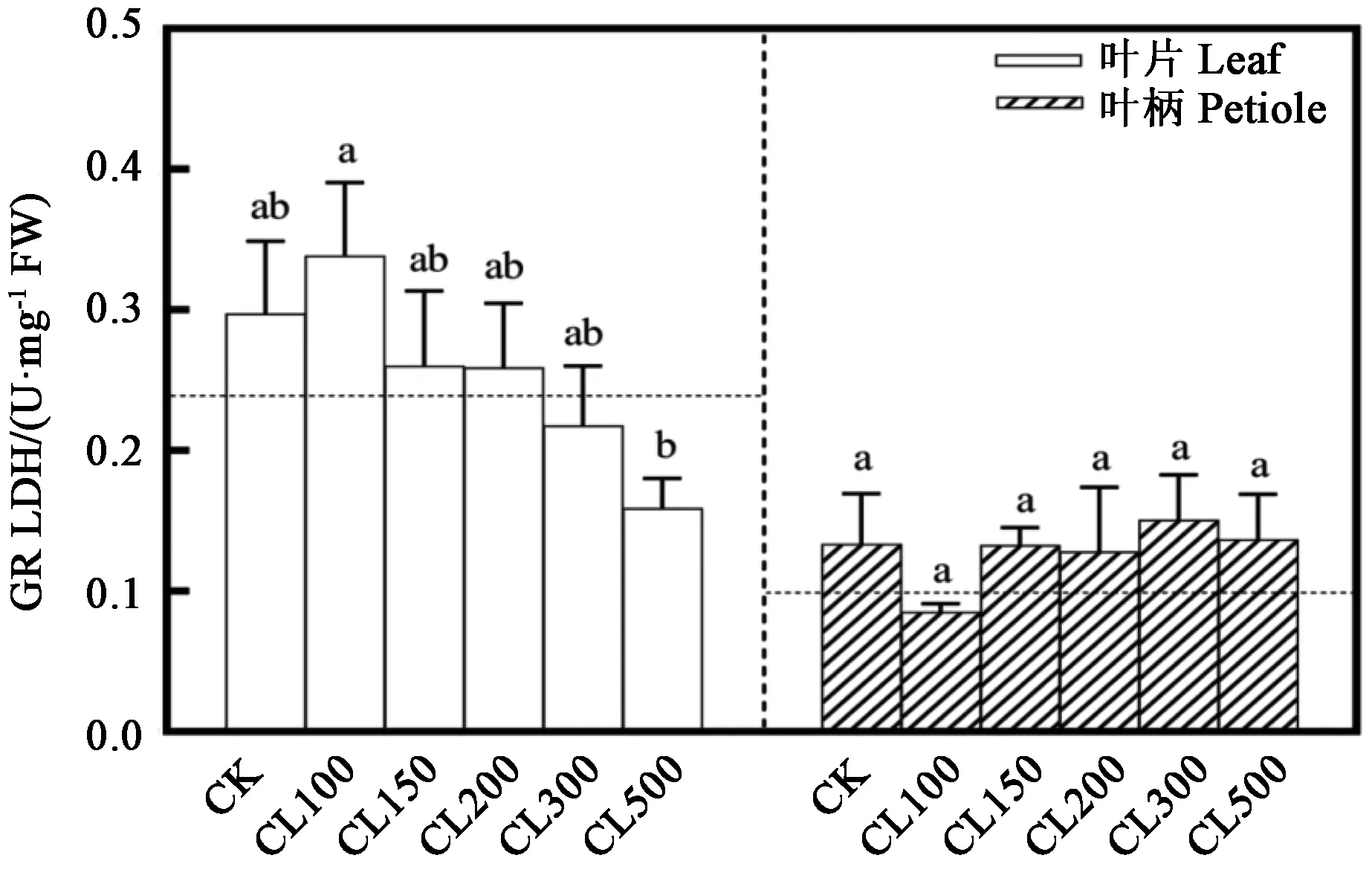
图6 采前LED连续光照光强对生菜GalLDH酶活性的影响Fig.6 Effects of LED continuous light intensity on the GalLDH activities in lettuces before harvest
2.4.3 采前LED连续光照光强对生菜APX、MDHAR、DHAR和GR酶活性的影响 采前LED连续光照光强处理对生菜AsA代谢循环中各种酶活性具有不同的影响。图7显示,非连续光照处理72 h(CK)后,生菜叶片的APX酶活性略微升高;而高光强连续光照处理72 h后,生菜叶片的APX酶活性明显增加。非连续光照处理72 h(CK)后,生菜叶柄的APX酶活性略微升高;而高光强连续光照处理72 h后,生菜叶柄的APX酶活性明显增加。非连续光照处理72 h(CK)后,生菜叶片的MDHAR酶活性略微降低至2.12 U·mg-1FW;而高光强连续光照处理72 h后,生菜叶片的MDHAR酶活性明显增加。非连续光照处理72 h(CK)后,生菜叶柄的MDHAR酶活性略微升高;而连续光照处理72 h后,生菜叶柄的MDHAR酶活性略微增加,但无显著差异。非连续光照处理72 h(CK)后,生菜叶片的DHAR酶活性略微降低至1.52 U·mg-1FW;而高光强连续光照处理72 h后,生菜叶片的DHAR酶活性明显增加。非连续光照处理72 h(CK)后,生菜叶柄的DHAR酶活性略微升高;连续光照处理72 h后,各处理中生菜叶柄的DHAR酶活性无显著差异。非连续光照处理72 h(CK)后,生菜叶片的GR酶活性升高了0.013 U·mg-1FW;而连续光照处理72 h后,CL200、CL300和CL500处理中生菜叶片的GR酶活性明显增加。非连续光照处理72 h(CK)和连续光照处理72 h后,各处理的生菜叶柄的GR酶活性均无显著差异。
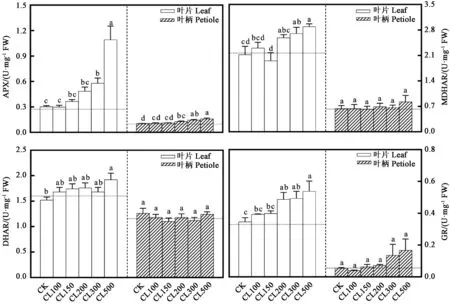
图7 采前LED连续光照光强对生菜APX、MDHAR、DHAR和GR酶活性的影响Fig.7 Effects of LED continuous light intensity on APX,MDHAR,DHAR and GR activities of lettuces before harvest
3 讨论
本研究结果显示,采前LED连续光照后生菜地上部鲜重和干重均显著增加,且随着连续光照的光照强度的提高,生菜叶片的净光合效率增大,生菜地上部的干物质含量增大,这与Sysoeva[19]等的研究结果一致。通过延长光合作用的时间而提高生菜叶片的净光合效率,促进光合产物的累积,从而使得生菜产量得到显著提高。因此,通过采收前集中连续LED红蓝光照射是一个增加产量的有效途径。
周晚来[9]通过对生菜进行50~200 μmol·m-2·s-1光强的采前连续光照发现,水培生菜的硝酸盐含量随连续光照光强的升高逐渐降低,可溶性糖及抗坏血酸含量随连续光照光强的升高而逐渐升高。本研究进一步将采前LED连续光照的最高光强处理提高到500 μmol·m-2·s-1,发现随着连续光照光强的升高,可溶性糖含量逐渐升高,硝酸盐含量逐渐降低,但在光强达到150 μmol·m-2·s-1之后继续增加,硝酸盐含量虽进一步降低,但已无显著差异。这可能是由于植物体内所含物质的含量都有个浓度范围,在150 μmol·m-2·s-1的72 h 连续光照后,生菜体内的硝酸盐含量已经达到下限,再通过升高连续光照的光强来进一步降低硝酸盐含量会逐渐变的困难。
本研究发现,随采前连续光照光强的升高,叶片AsA含量逐渐增加,叶片DHA含量先升高后降低。随采前连续光照光强的升高,叶柄的AsA含量略有增加,DHA含量略有降低,与非连续光照处理(CK)对比,各处理间叶柄的AsA和DHA含量均无显著差异。植物体内的抗坏血酸含量取决于其合成和分解的速率及合成和分解的动态平衡状态[20]。本研究结果表明,随着连续光照光强的提高,叶片中GalLDH活性逐渐降低。因此,采前LED连续光照光强对生菜叶片GalLDH活性的影响与抗坏血酸含量变化相反。研究表明,AsA含量的高低主要与APX、MDHAR、DHAR和GR等酶活性的高低有关[21-22]。本研究结果表明,叶片APX酶活性随连续光照光强的增大而增大,与AsA含量的变化趋势一致,说明连续光照光强越高,APX酶活性越高,催化分解AsA的量越多。叶片MDHAR酶活性随连续光照光强的增大而增大,与AsA含量的变化趋势一致,说明连续光照光强越高,MDHAR酶活性越高,催化分解为DHA那部分的MDHA转化合成AsA的量越多。叶片DHAR酶活性随连续光照光强的增大而增大,与AsA含量的变化趋势一致,说明连续光照光强越高,DHAR酶活性越高。叶片GR酶活性随连续光照光强的增大而增大,也说明连续光照光强的升高促进了DHA还原为AsA的效率。可见,连续光照光强对AsA含量的影响主要是与参与AsA再生循环系统的MDHAR、DHAR和GR酶活性相关。
