植物覆盖对蕉园土壤微生物群落功能多样性的影响
2022-07-10魏加弟王永芬杨佩文俞艳春郑泗军徐胜涛
魏加弟,王永芬,何 翔,杨佩文,俞艳春,郑泗军,4,徐胜涛
(1云南省农业科学院农业环境资源研究所,昆明650205;2云南农业大学动物科学技术学院,昆明 650201;3云南省农业科学院热带亚热带经济作物研究所,云南 保山 678000;4国际生物多样性中心,昆明 650205)
0 引言
中国的香蕉主产区主要分布在广东、海南、福建、云南、广西、台湾六省[1],是当地农民增收、农业增效的重要产业[2]。近年来,由于地表裸露造成蕉园土壤退化问题阻碍了中国香蕉产业发展,如土壤板结、养分流失和微生物群落结构被破坏等。通过采用保护性耕作措施,可有效改善蕉园土壤肥力,提高蕉园土壤生产力,促进香蕉产业健康可持续发展。土壤是农业赖以发展的基础性资源,土壤肥力直接影响着作物产量和品质的形成。目前,蕉园管理过程中蕉农为了避免其他植物与香蕉的水肥竞争,往往清除地面全部杂草,导致蕉园土壤大范围的呈裸露状态[3],破坏了土壤结构,造成了土壤板结、土壤保水保肥性能降低及土壤透水透气性减弱等土壤退化问题[4]。另外,蕉园肥料施用不合理,导致土壤酸化及土壤中有害元素富集[5]。因此,为了改善蕉园土壤肥力状况,提高蕉园土壤生产力,促进蕉园土壤可持续利用,需转变目前的蕉园管理模式,采用用地养地为一体的管理措施。植物覆盖措施是一种科学的土壤管理制度,中国于20世纪90年代引入植物覆盖技[6],植物覆盖措施在不影响作物生长的前提下改善土壤理化性质,这种模式较单一种植作物而言不仅增加了土壤中的生物量,而且提高了土壤的生产力,在空间分布及时间利用过程中形成了互利共生的关系,能够增大土壤储水量、改善土壤结构、提高土壤肥力、调节田间小气候等功能[7],对提高土壤生产力,促进中国农业健康可持续发展有重要意义。土壤微生物是土壤的重要组成部分,推动整个土壤生态系统中的能量流动和物质循环,在促进植物吸收养分及调节土壤肥力与健康等方面有着不可替代的作用[8]。土壤微生物群落结构不仅受土壤温度、水分、通气性及土壤肥力等因素的影响,也受耕作措施的影响[9],体现了土壤中各种生物化学过程的强度及方向,是土壤生态系统恢复的敏感性指标[10]。植物覆盖措施通过覆盖作物根部分泌的各种微生物,直接有效的增加蕉园土壤微生物群落结构多样性;也可以通过改善田间小气候间接提高蕉园土壤微生物多样性。随着分子生物学技术的不断发展和完善,土壤微生物多样性研究将在土壤生态系统功能恢复及重建方面提供更多的理论依据[11]。植物覆盖措施已广泛应用于提升果园的土壤肥力,但是由于与现阶段蕉农的种植理念相悖,有关于植物覆盖对蕉园土壤微生物多样性的研究较少,植物覆盖能有效的将蕉园的农业生产与土壤保持相结合,是用地养地为一体的保护性田间管理模式。本研究利用微生物Biolog鉴定技术探究植物覆盖对蕉园土壤微生物多样性的影响,以期为改善蕉园土壤质量、促进香蕉产业可持续发展提供理论依据。
1 材料与方法
1.1 试验地概况
试验区位于云南省保山市潞江镇(98°53′14″E,24°57′58″N),海拔约700 m,年平均气温21.3℃,全年基本无霜,绝对最高气温40.4℃,绝对最低气温0.2℃,≥10℃活动积温7800℃,年日照时数2333.7 h,年降水量750 mm,年蒸发量2100 mm,相对湿度70%,属于亚热带干热河谷气候类型。土壤属砂壤土,试验地土壤、气候条件和土壤类型均能满足香蕉正常生长。香蕉品种选择该试验区主栽品种‘云蕉1号’,在该地区生长周期约1年。
1.2 试验方法
试验以常规种植行间地表裸露为对照(CK),分别以行间种植自然生杂草覆盖(Natural weed,NW)和人工种植豆科植物覆盖(Cover plant,CP)为处理,研究香蕉种植区(Plant Zone,PZ)与覆盖区(Cover Zone,CZ)土壤微生物功能多样性差异。其中,自然生杂草以牛筋草[Eleusine indica(L.)Gaertn.]和马唐[Digitaria sanguinalis(L.)Scop.]为主,豆科覆盖植物品种为大翼豆[Macroptilium atropurpureum(DC.)Urb]。大翼豆种植时间为2018年4月,种植方式为条播。覆盖区可以通过刈割和补播控制其高度及覆盖度,无其它管理措施。在香蕉生长前期,杂草及大翼豆高度应低于30 cm避免影响香蕉生长,需保证60%以上的覆盖度才能达到覆盖效果。试验设4次重复,采用随机区组排列。小区面积为100 m2,每小区定植香蕉苗40株。香蕉苗为宽窄行种植,宽行行距3.5 m,窄行行距1.5 m,香蕉苗在窄行上采取“之”字形种植,株距2.0 m,2018年7月定植香蕉苗,香蕉栽培和水肥管理参照当地香蕉种植模式。
1.3 测定指标及方法
土壤样品用“S”形沿小区对角线采集,土壤样品分为种植区和覆盖区,种植区土壤为香蕉假茎周围30 cm内的土壤,覆盖区土壤为宽行种植覆盖植物的土壤。采样时间为2019年7月和2020年7月,取样深度为0~30 cm。
土壤微生物功能多样性测定:采用Biolog系统鉴定法,用电子分析天平称取10 g新鲜土壤加入到盛有90 mL灭菌生理盐水(85%NaCl)的锥形瓶中,置于250 r/min的摇床上震荡30 min,待土样充分摇匀后取出静置10 min,用移液枪吸取5 mL的上清液加入盛有45 mL灭菌生理盐水的锥形瓶中,重复稀释步骤得到10-3菌悬液,将此菌悬液接种到Biolog生态板中,每个微孔接种150 uL,接种好的板置于25℃黑暗条件下培养,每隔24 h在微生物自动鉴定仪上测定590 nm处的吸光度值,共测定216 h。
平均颜色变化率(AWCD):是反应土壤微生物利用单一碳源的能力,是微生物代谢活性的一个重要标志,AWCD值越大,说明微生物活性越高。计算公式如式(1)所示。

式中Ci为第i个非对照孔在590 nm下的吸光度;R为对照孔吸光度;n为碳源种类数(n=31);Ci-R≤0的孔在计算中记为0;每间隔24 h测定AWCD值,绘制AWCD与培养时间的动态变化图。
采用接近渐近线的168 h处的吸光值进行标准化处理,计算表示微生物碳源代谢功能多样性特征指数,包括:Shannon-Wiener多样性指数(H):是评估微生物物种的丰富度与分布均匀程度的综合指标。Simpson优势度指数(D):是对微生物多样性方面的集中性的度量,反映土壤群落中最常见物种。McIntosh均匀度指数(U):反映群落物种均匀度。McIntosh Evenness指数(E)为McIntosh均匀度指数。Richness Index丰富度指数(R)指被利用的碳源总数目,为每孔中(Ci-R)的值大于0.20的个数。计算公式如式(2)~(6)所示。

式中Pi为第i个非对照孔吸光度与所有非对照孔吸光度总和的比值;Pi=(Ci-R)/∑(Ci-R);ni是第i孔的相对吸光度值;S为被利用的碳源数目。
1.4 数据处理
采用Excel 2010软件对试验数据进行数据整理,使用SPSS 16.0统计软件进行单因素方差分析和主成分分析。
2 结果与分析
2.1 土壤微生物平均颜色变化率(AWCD)分析
AWCD值可以衡量土壤微生物对碳源的利用能力,是微生物代谢活性的重要标志,绘制AWCD随微生物培养时间变化的动态曲线,可以表示不同培养时间土壤微生物代谢强度的动态变化。如图1所示,AWCD曲线变化特征为:24 h后AWCD开始迅速升高,48 h后进入指数期,192 h后进入稳定期。图1中2019年种植区(PZ),NW、CP、CK处理下土壤微生物AWCD曲线的增长幅度在0~168 h无明显差异,但是在第192 h后NW和CP处理的土壤微生物AWCD曲线高于CK。图1中2019年覆盖区、2020年种植区及2020年覆盖区,土壤微生物代谢强度变化特征为:CP处理的AWCD曲线增长幅度显著高于NW和CK,说明大翼豆覆盖下蕉园土壤微生物代谢活性较高,对于覆盖区土壤微生物而言,其碳源代谢强度大小为:CP>NW>CK。

图1 植物覆盖下蕉园不同类型土壤微生物AWCD值变化
2.2 土壤微生物对不同碳源的利用
Biolog-ECO微平板的31种碳源可归纳为6类,分别为糖类、氨基酸类、羧酸类、聚合物类、胺类和酚酸类。用培养了168 h的碳源代谢强度值分别计算6类碳源的利用率(如图2),发现不同年份种植区和覆盖区土壤微生物对6种碳源的利用情况大致相同。2019年PZ区,CP和NW处理下土壤微生物对糖类、氨基酸类利用较好,对羧酸类、聚合物类利用较差。随着覆盖年限增加,2020年CP和NW处理促进了种植区土壤微生物对羧酸类及酚酸类利用。2019年CZ区,NW处理下土壤微生物对糖类碳源利用较好,CP处理下对胺类及酚酸类碳源利用较好,随着覆盖年限增加,CP和NW处理下土壤微生物对糖类和氨基酸类碳源的利用显著高于对照区。

图2 蕉园植物覆盖对不同类型土壤微生物的碳源利用影响
2.3 土壤微生物群落结构多样性分析
如表1所示,随着覆盖处理的年限增加,土壤微生物各个指数都升高。对种植区(PZ)土壤微生物而言,2019年NW和CP处理下土壤微生物Shannon-Wiener、McIntos指数和Richness Index指数与CK处理差异显著(P<0.05),到2020年,土壤微生物Shannon-Wiener指数和Simpson指数在各处理间差异显著,结果表现为:CP>NW>CK。对于覆盖区(CZ)土壤微生物而言,2019年NW和CP处理下土壤微生物McIntosh、McIntosh Evenness指数和Richness Index指数与CK处理差异显著(P<0.05),2020年CP处理下土壤微生物Shannon-Wiener指数和Simpson指数与CK处理差异显著,CP处理土壤微生物Shannon-Wiener指数和Simpson指数较CK区分别高出2.11%、1.04%。
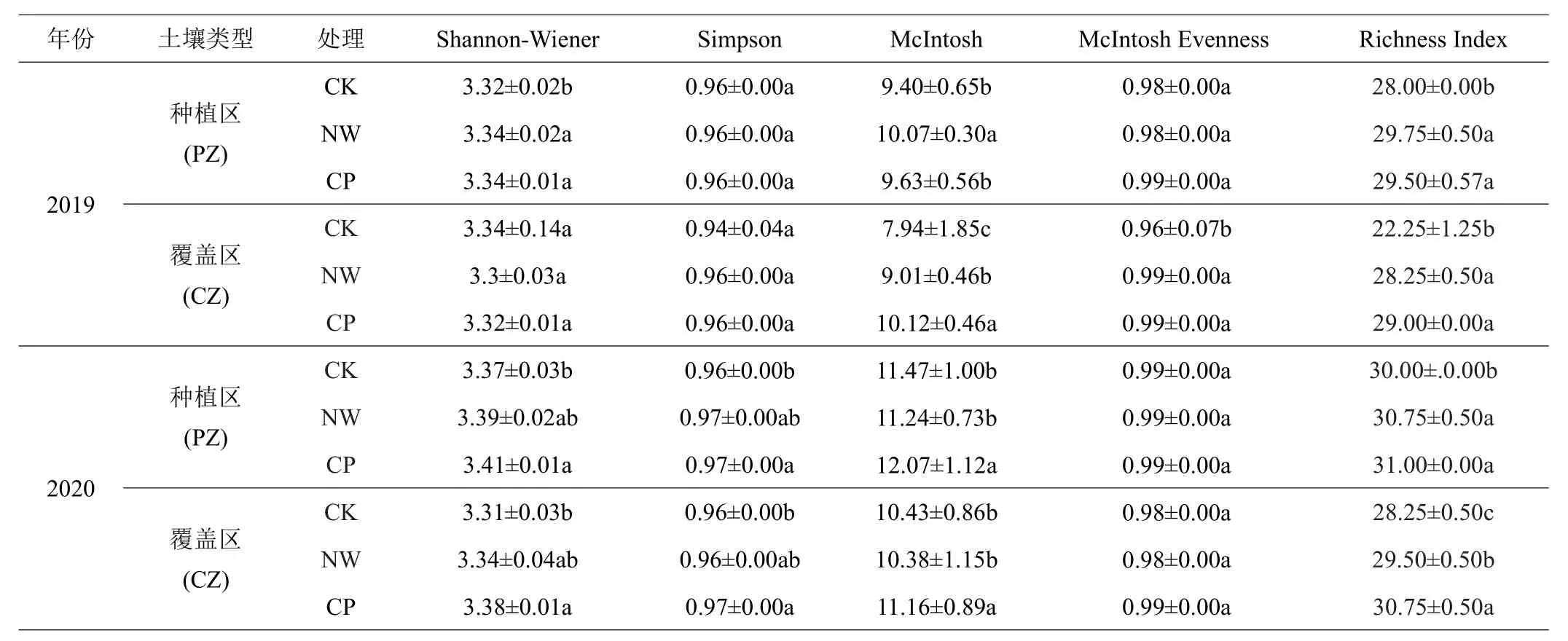
表1 植物覆盖对蕉园土壤微生物群落结构多样性影响
2.4 土壤微生物群落主成分分析
根据培养168 h的土壤微生物碳源代谢强度值,对种植区和覆盖区各处理土壤微生物群落的碳源利用特征进行主成分分析。由图3可知,对种植区(PZ)土壤微生物而言,PC1的贡献率为27.6%,PC2的贡献率为22.0%,其中CP分布在第一象限,NW分布在第二象限。对覆盖区(CZ)土壤微生物而言,PC1的贡献率为48.8%,PC2的贡献率为19.3%,因此各处理间的差异主要体现在PC1轴,图中CP主要分布在PC1的正半轴,NW和CK主要分布在PC1的负半轴。

图3 蕉园植物覆盖下不同类型土壤微生物群落功能的主成分分析
载荷图反映了主成分中包含的因子种类,包含某一类的碳源因子数量越多,则证明该碳源对主成分的影响也越大。从表2可以看出,对种植区土壤微生物而言,对PC1贡献较大的碳源有15中,其中羧酸类6种、氨基酸类4种、糖类3种、聚合物类2种,可见对PC1起主要作用的碳源为糖类、氨基酸类和羧酸类;在PC2中,贡献较大的有3类碳源,其中糖类6种、胺类2种和酚酸类1种,对PC2起主要作用的碳源为糖类和胺类。

表2 蕉园植物覆盖下种植区土壤微生物主成分载荷矩阵
从表3可以看出,对覆盖区土壤微生物而言,对PC1贡献较大的碳源有15中,其中糖类8种、氨基酸类4种、羧酸类5种、聚合物类2种,可见对PC1起主要作用的碳源为糖类、氨基酸类和羧酸类;在PC2中,贡献较大的有3类碳源,其中糖类1种、胺类1种和酚酸类2种,对PC2起主要作用的碳源为酚酸类。

表3 蕉园植物覆盖下覆盖区土壤微生物主成分载荷矩阵
3 结论
植物覆盖可以改善蕉园土壤微生物功能多样性,且随着覆盖年限的增加效果越显著,能一定程度解决蕉园土壤退化问题。不同覆盖植物对土壤微生物功能多样性影响差异显著,大翼豆覆盖较自然生杂草覆盖更能改善土壤微生物群落多样性,在蕉园裸露行间人工种植大翼豆的方法可有效减轻土壤退化程度,以植物多样性促进土壤微生物多样性的提高,提高香蕉园土壤健康可持续利用。
4 讨论
土壤微生物在调节土壤养分循环和能量流动方面发挥着重要作用,并且对土壤管理措施非常敏感[12],通过对蕉园土壤微生物群落结构的研究可以在一定程度上表征蕉园土壤生态系统稳定性及土壤养分变化。土壤微生物群落对碳源的整体利用情况表征了土壤微生物活性状况,通常对碳源利用的效果越强说明其微生物活性越高。本研究中Biolog-ECO微平板培养微生物的时间为216 h,0~24 h为滞后期,AWCD曲线增长缓慢,土壤微生物正在适应微平板的环境;24~168 h为指数期,AWCD曲线指数型增长,土壤微生物代谢强度最旺盛;168 h以后为平缓期,AWCD曲线与横坐标轴趋向平行,此时由于微平板中的碳源底物量无法维持微生物代谢,所以其代谢活性降低,该曲线符合微生物在Biolog-ECO板中的生长规律[13-15]。本研究认为连续两年在蕉园裸露行间覆盖植物能显著提升土壤微生物碳源利用多样性(P<0.05),并且大翼豆覆盖(CP)>自然生杂草覆盖(NW)>无覆盖(CK),相同条件下由于大翼豆也可以作为绿肥,提高土壤含氮量,增加土壤有机质含量,所以大翼豆覆盖比自然杂草覆盖在提高土壤微生物活性方面更有效,因此在蕉园种植大翼豆更有利于蕉园土壤环境的改善[16]。该结果与王锐等[17]、魏树伟等[18]的研究结果一致,认为植物覆盖可以提高土壤微生物活性因为覆盖作物不仅保持土壤水分,为微生物提供良好的生存环境,同时,覆盖作物本身还有大量有机质和N、P、K等元素,还田以后为土壤微生物提供了丰富的营养,促进土壤微生物的繁殖。
地上植物不同,土壤微生物数量及碳源利用类型也不同,土壤微生物对碳源利用规律直接反映了其代谢偏好[19]。Biolog-ECO生态板中,至少有9中碳源是根系分泌物的组分,本研究结果表明覆盖区土壤微生物在NW与CP处理之间的碳源利用差异较大[20]。研究发现覆盖区土壤微生物对糖类及氨基酸类碳源的利用较CK区高,而氨基酸类碳源大部分来自植物根系分泌物[21],糖类极易被微生物同化,可以形成碳源比较丰富的环境[22],说明植物覆盖可以通过增加根系分泌物或促进土壤中的糖类形成,为微生物生长繁殖提供物质保障,从而提高土壤微生物活性及多样性。酚酸类化合物大部分是三羧酸循环的中间体,对土壤微生物的活性影响很大,土壤中的酚酸类物质通过改变土壤养分、pH及化感作用等显著影响土壤微生物的多样性及群落结构[23],本研究中随着植物覆盖年份的增加土壤微生物对酚酸类碳源的利用能力增强,说明植物覆盖可以通过提高土壤中酚酸类碳源促进土壤微生物多样性。
目前土壤微生物Simpson(D)指数、McIntosh(U)和Shannon-Wiener(H)指数被广泛应用于衡量土壤微生物多样性[24]。本研究发现植物覆盖措施可以显著提高香蕉种植区土壤微生物群落结构的多样性、优势度及微生物分布的均匀度(P<0.05),该结果与钟哲科等[25]的研究结果一致,其原理在于覆盖作物有利于土壤增温保湿[26]、促进土壤的物质循环,从而提高香蕉种植区土壤微生物群落结构均匀度。本研究中随着覆盖年限的增加种植区NW、CP间土壤微生物Shannon-Wiener指数和Simpson指数差异显著,且CP>NW,说明植被类型是影响种植区土壤微生物群落结构多样性的重要因素之一,在环境条件一致的情况下,不同植物对土壤微生物群落结构影响不同[27]。对于覆盖区土壤微生物而言,NW和CP处理下土壤微生物McIntosh(U)、McIntosh Evenness(E)、Richness Index(R)指数较 CK显著增强,这是因为覆盖作物不仅可以通过其根系活动直接改善土壤的团粒结构,为微生物生长繁殖提供良好的环境,而且还为土壤中的微生物提供了丰富的碳源、氮源和其他所需养分,有利于微生物的生长繁殖,从而提高微生物活性[28-30]。大翼豆最突出的特点是耐高温干旱,并且具有抑制草害、保持水土、土改土壤肥力等特点,被广泛应用于改善干热河谷地区土壤生态条件[31]。本研究结果表明,CP土壤微生物群落结构的McIntosh均匀度指数、Richness Index丰富度指数及McIntosh Evenness指数较NW上升效果更显著,表明豆科牧草的固氮作用使土壤中的C/N下降,加快了土壤中养分的分解,提高了土壤微生物活性,大翼豆覆盖对改善干热河谷地区土壤肥力具有重要意义。
主成分分析结果表明,不同处理土壤微生物群落的代谢多样性有明显差异。覆盖区土壤微生物碳源利用特征是:CP能较好的利用糖类、氨基酸类及羧酸类碳源,NW能较好的利用胺类和酚酸类碳源,CK对各类碳源利用较弱,因此地上植物群落结构差异是导致土壤微生物功能多样性表现出空间异质性的主要原因,这与井赵斌等[32]关于植物覆盖对猕猴桃园土壤微生物群落结构的影响的研究结果一致。种植区CK、CP和NW都能很好的利用氨基酸类和羧酸类碳源,而CP和NW也能较好地利用糖类和胺类碳源,这一结果可能与在蕉园管理过程中化肥和农药降解到地表有关,土壤微生物对农药及化肥反应敏感,从而影响其碳源利用特征。糖类、氨基酸类和羧酸类碳源是优势碳源也是敏感碳源[33],本研究中,糖类、氨基酸类和羧酸类碳源是土壤微生物的主要碳源类型,这3类碳源中微生物对糖类的利用率最高,这与王楠楠等[34]的研究结果一致。
