齐口裂腹鱼上溯过程中“冲刺-滑行”行为对水动力的响应
2022-07-08李沁芝邓月曦唐锡良严忠銮王小明卢晶莹安瑞冬
李沁芝 邓月曦 唐锡良 严忠銮 王小明 卢晶莹 李 嘉 安瑞冬
(1.四川大学水力学与山区河流开发保护国家重点实验室,成都 610065;2.中国三峡建工(集团)有限公司,成都 610041;3.中国长江三峡集团有限公司,武汉 430010)
大坝等拦河建筑物在带来巨大经济效益的同时,也破坏了河流的连通性,阻断了鱼类的洄游通道,导致鱼类资源大幅度减少。修建鱼道是缓解水利工程开发不利影响和水资源保护的有效措施,有效的过鱼设施设计需要结合鱼类行为学。鱼类的运动行为具有多样性,包括游泳和滑行等[1]。其中鱼的游泳是一种复杂的、综合的运动,具有对各种环境变化做出可塑性响应的能力[2]。随着机电一体化技术、计算机技术、流体力学和仿生学等相关学科的发展,研究人员研制出了多种仿生机器鱼,其游动机理将是未来仿生机器鱼研究的关键[3]。不同的鱼表现出来的游泳行为也不尽相同,但除了在鱼道进出口等高流速区采用冲刺游泳行为外,一般情况下都采用“冲刺-滑行”游泳行为[4]。“冲刺-滑行”游泳行为反映了鱼类在极短时间内逃避敌害和越过水流障碍的能力,是鱼类生活史中重要的游泳行为之一[5]。量化鱼类在复杂流场中的游泳行为显得尤为重要[6]。冲刺阶段鱼通过摆尾来获取前进的动力,滑行阶段鱼的身体接近直线状态[7]。表征该行为的指标很多,比如游泳动力学、游泳能力和游泳模式[8]。通过分析鱼类游泳动力学指标,例如摆尾角度与摆尾频率等重要的基础参数,有助于理解目标鱼类的游泳姿态转换过程及运动的能量消耗[9]。Tudorache等[10]在较长的水槽中测定鲤(Cyprinus carpio)的临界游泳能力,发现其可以通过“冲刺-滑行”游泳获得更高的临界游泳速度。Li等[11]发现鱼通过调节内在循环以维持所需的速度,通过改变冲刺与滑行的时间比,保持“冲刺-滑行”周期的稳定。Zhao等[12]提出了一种基于鱼的关键特征点识别的摆尾频率计算模型,用于实时检测鱼的摆尾频率。Goettel等[13]通过在一个宽浅的水槽中增减壁面处障碍物的厚度,来改变水槽中的水流流态,将不同的流场与视频录像中的鱼类轨迹叠加,讨论了鱼类对不同流场的响应。
齐口裂腹鱼(Schizothorax prenanti)隶属鲤科(Cyprinidae)裂腹鱼亚科(Schizoth Pracinae)裂腹鱼属(Schizothorax),主要分布于我国长江上游的金沙江、岷江、大渡河、青衣江及乌江下游等水域,是我国西南山区河流中较具代表性的优势鱼种,是我国特有的重要经济鱼类[14]。水流速度障碍在自然界中广泛存在,它能够刺激鱼类感觉器官产生相应行为反应,对其摄食、代谢及生长繁殖造成影响[15]。鱼类“冲刺-滑行”游泳行为主要在鱼类捕食、逃逸敌害、受到刺激后的应激和越过水流障碍过程中应用[16]。针对摆尾频率和摆尾幅度的计算目前主要采用的方法包括视频软件分析,跟踪定位及人工计数等方法[17]。目前的研究对游泳动力学指标中摆尾角度的自动识别与计算方法较少,且研究侧重于鱼尾鳍缺失对鱼游泳能力的影响,如Cai等[18]通过测量了3组鱼(完整尾鳍、部分尾鳍和无尾鳍)的游泳速度和摆尾频率,发现摆尾频率降低会明显影响其游泳能力。通过解译其游泳行为模式,对模拟洄游性鱼类通过自然水流障碍或水工建筑物提供了理论支撑[19]。因此本研究运用具有流速梯度的水槽创造非均匀流场条件,得到齐口裂腹鱼在室内试验水槽内进行上溯的视频图像。对其游泳姿态进行图像识别,计算其上溯全过程的游泳动力学指标摆尾角度与摆尾频率。构建了新的指标体系,引入滑行流速系数、摆尾模式等。在此基础上做了新的量化分析,实现生态学与水动力学的耦合研究,进而得到其上溯过程中“冲刺-滑行”行为对水动力学条件的响应规律,对鱼类行为学研究中齐口裂腹鱼的行为学研究进行补充。
1 材料与方法
1.1 生态行为学试验设计
试验装置本研究采用的试验装置为宽4.0 m×深1.0 m×长20.5 m的无坡度的钢化玻璃-钢铁框架结构的大型明渠水槽(图 1)。试验装置顶部放置4个摄像头(WIM SkyStar,30 fps),共同记录试验鱼的游泳行为,为后续运用行为学软件logger pro得到完整上溯过程的运动轨迹坐标创造条件。为了减小流动摩擦以及降低边界层效应,水槽两侧和底板接近光滑。通过固定电机频率使水流稳定。进口处格栅密度最高的流态I的流速略高于封闭游泳室中试验鱼种齐口裂腹鱼的感应游泳能力(0.01—0.13 m/s)。格栅较少的流态Ⅱ的流速接近齐口裂腹鱼临界游泳速度(0.65—1.09 m/s),对于流态Ⅲ的无格栅通道,流速设计在暴发游动能力范围(0.85—1.53 m/s)内。
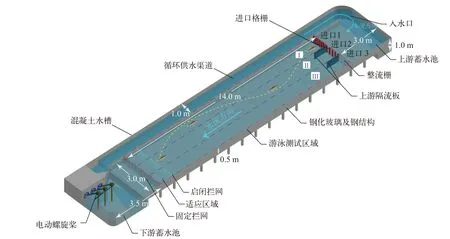
图1 试验装置图Fig.1 Testing device
试验对象试验齐口裂腹鱼体长(26.91±2.35) cm,叉长(29.57±2.49) cm,总长度(32.32±2.73) cm,体重(304.21±77.15) g。试验前将所有试验鱼在室内暂养7d,暂养期间24h持续充氧,每2天更换暂养水的10%,每天投食饵料并吸粪便和残渣1次。试验条件:水温(15.83±0.24)℃,pH (7.6±0.4),溶解氧饱和度保持在95%左右,自然光照。
试验过程在试验装置适应区域放入一尾健康的试验鱼,经过0.5h的适应,试验鱼已基本适应当前温度和流速条件。移除活性屏障,让试验鱼在试验装置的游泳测试区自由游泳。同时利用安装在水面上方4 m的4台摄像机连续记录试验鱼的游动行为。当试验鱼经过整个游泳测试区域,最终进入上游3个入口之一时,认为该试验鱼成功完成上溯,结束试验。当试验鱼无上升动机且在试验装置适应区域停留超过1h时,同样结束试验。采用高分辨率声学多普勒测速仪,对游泳测试区域(图 1中的矩形)沿纵向、横向和垂直方向测量其瞬时流场分量,频率为50 Hz,持续60s。纵向和横向测量距离间隔分别为0.25和0.1 m,共记录了1421个测量位置。
1.2 游动姿态的图像识别与鱼体几何模型构建
本研究选取了上溯时间较短,视频亮度较高,游动状态较为清晰的10尾鱼(图 2)。为保证鱼体摆尾状态的连续性,视频帧率选择为30fps。在鱼类快速启动游泳的过程中,鱼体首次肌肉收缩后的形态特征不尽相同,有些鱼类呈“C”型(早期也被称为“L”型),而有的鱼类形态呈“S”型[20]。鱼的体轴主要包括主轴、矢轴与横轴,主轴亦称中心轴,是由头部到尾部纵贯体中央的一个轴,矢轴是从鱼体最高处作一垂线,横轴横贯鱼体中心[21]。通过分析大量试验图片,认为齐口裂腹鱼在冲刺阶段鱼体大都呈“C”型。保留鱼体基本特征,构建了鱼体俯视的二维几何模型(图 3),其中各特征点含义如表 1所示。直线AE为鱼体主轴所在位置。本研究定义了一个铰点E,认为鱼体在摆尾时身体可分为刚体段与柔体段,铰点所在位置即为两部分的分界点。θ即为鱼体摆尾角度。由某一试验鱼的图像识别结果所构建的鱼体俯视二维几何模型(图 4)。
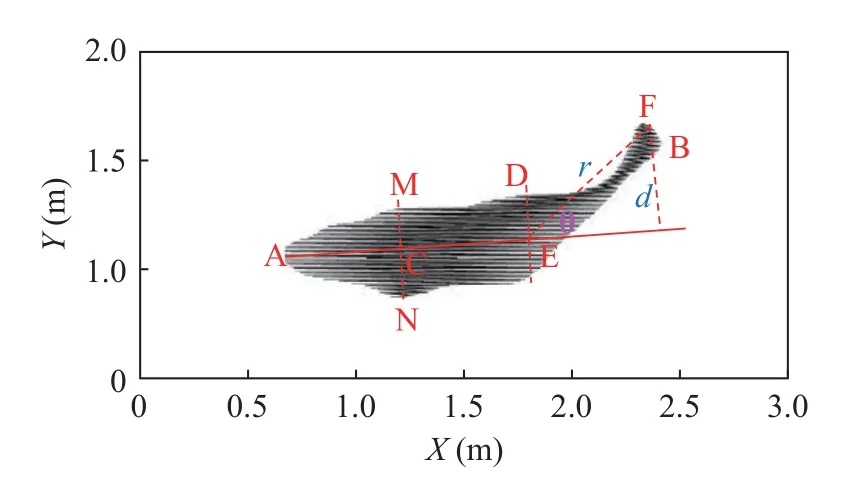
图4 图像识别与模型构建Fig.4 Image recognition and model building

表1 鱼体俯视简化二维几何模型中各特征点含义Tab.1 The meaning of each feature point in the simplified 2D geometric model of the fish body in top view

图2 上溯过程视频分帧解析图Fig.2 Upstream process video frame splitting analysis
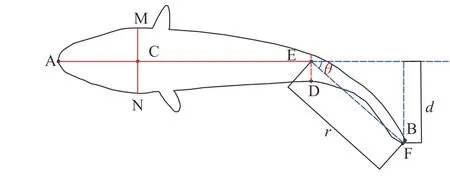
图3 鱼体俯视简化二维几何模型Fig.3 Simplified 2D geometric model of the fish body in top view
1.3 游动行为的描述
摆尾角度计算在对组成鱼体轮廓的点的直角坐标进行提取的基础上,运用式(1)逐个计算鱼体主轴两侧轮廓线上,相邻两点连线的斜率k,结果存在1、0和−1三种值。进一步根据该斜率计算结果的变化情况,可以分别提取鱼体主轴两侧的第一处斜率突变点M(XM,YM)、N(XN,YN),及鱼体摆尾时凹侧的第2处斜率突变点D(XD,YD)。运用式(2)得到线段MN的中点C坐标(XC,YC)。根据模型中各线段的几何位置关系,运用式(3)计算摆尾角度。式(3)中的两个参数均为距离值,其中d值为F点到直线AE的距离,r值为F点到E点的距离,关键在于铰点E坐标的计算。先假设出铰点E坐标,结合鱼体俯视二维几何模型中的点线关系,联立求解二元一次方程组,进而求得关键点E的坐标。
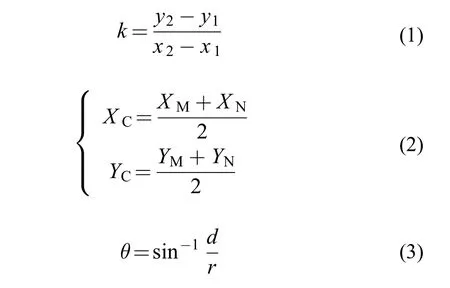
摆尾频率计算摆尾频率指的是1s内鱼摆尾的次数,从鱼尾摆到最高点开始到下一次摆到最高点计为1次。通过对10尾鱼上溯视频进行逐帧分析,计算得到10尾鱼上溯过程中的摆尾频率。
2 结果
2.1 冲刺摆尾的水力因子偏好
摆尾位置冲刺阶段鱼体姿态的改变往往体现在较大幅度的摆尾。绘制10尾鱼摆尾位置热点图(图 5)。结合图 6的流场图分析得到,摆尾行为多发生在不同流速区域交界处,即存在明显的流速梯度处,如y=1 m与y=2 m附近区域。这说明齐口裂腹鱼在上溯过程中往往会选择在具有流速梯度的区域进行冲刺,从而适应非均匀流场。
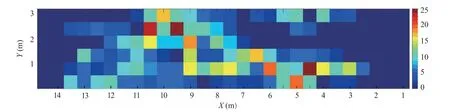
图5 摆尾位置热点图Fig.5 Hot spot diagram of tail-beat position
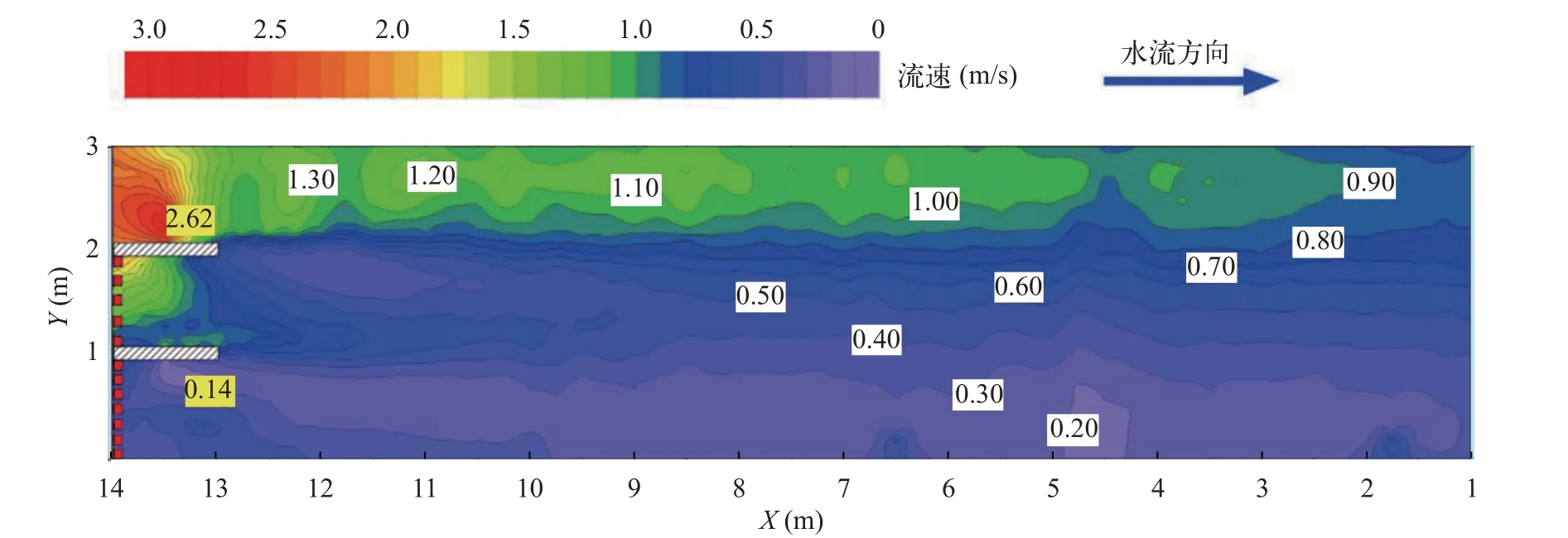
图6 流场插值分布图Fig.6 Flow field interpolation distribution
偏好流速范围“冲刺-滑行”游泳由一个或多个连续的冲刺游泳和紧随的滑行游泳组成,冲刺游泳阶段包括从稳态迅速增加到高游速的快速启动阶段和尾鳍等幅摆动维持高游速的匀速阶段,滑行游泳阶段包括从高游速逐渐降到相对水流静止的减速阶段和停止游泳被水流往后冲的阶段[15]。分别绘制10尾鱼摆尾角度与摆尾频率分布图(图 7和图 8),统计得到其冲刺阶段喜好摆尾角度为25°—35°,喜好摆尾频率为2.5—3.5次/s。进一步提取其做出喜好游泳动力学指标范围内的摆尾行为,所对应的摆尾位置的水流速度。叠加绘制“所有摆尾”情形与“喜好摆尾”情形的频率分布图,后者频率高于前者频率片段所对应的流速,为本试验条件下齐口裂腹鱼的偏好流速(图 9)。在本试验条件下,其冲刺阶段偏好流速为0.20—0.40 m/s。
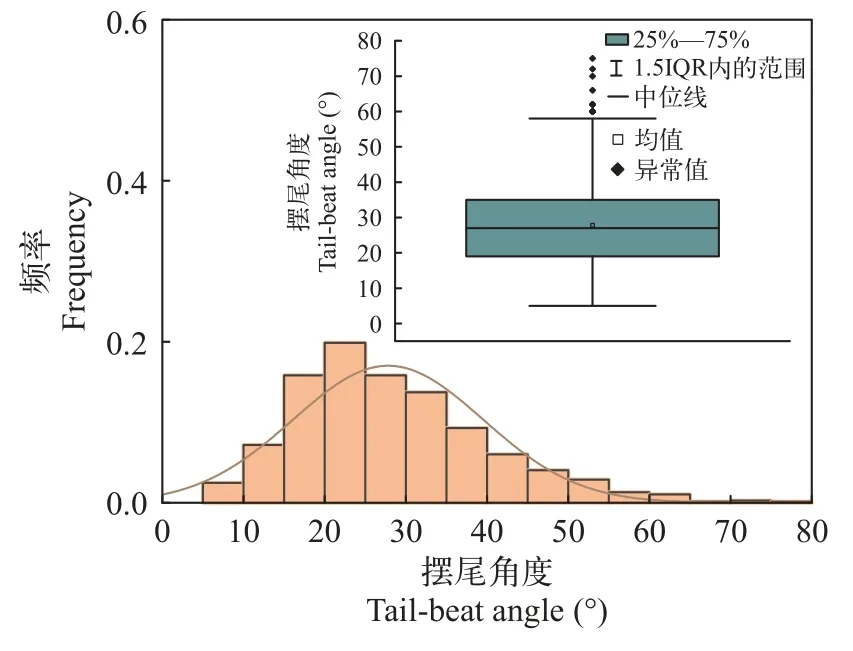
图7 摆尾角度分布图Fig.7 Tail-beat angle distribution and box line diagram
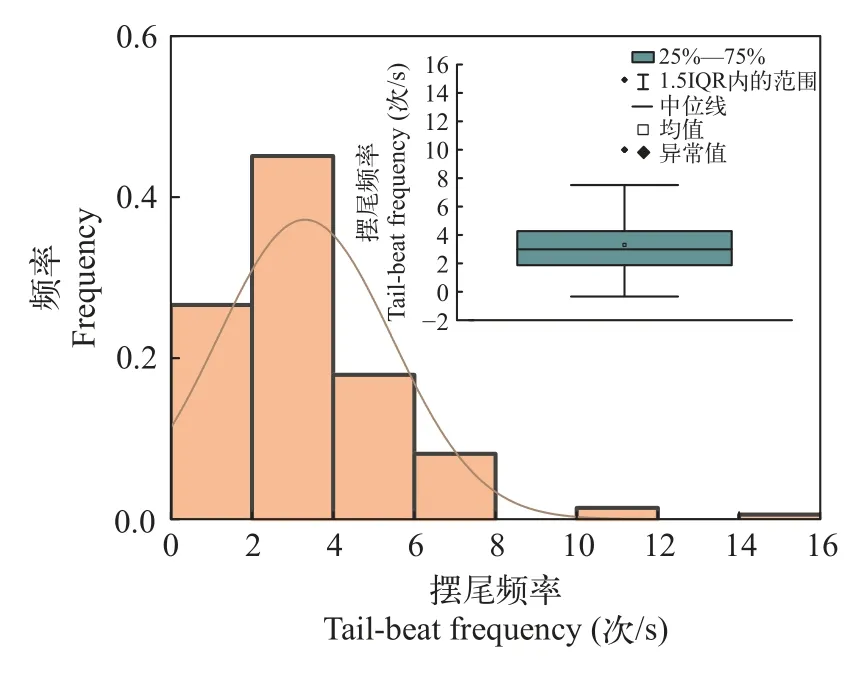
图8 摆尾频率分布图Fig.8 Tail-beat frequency distribution and box line diagram
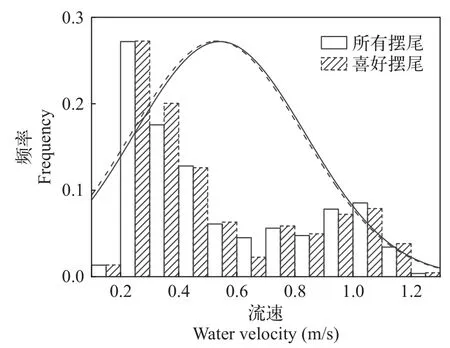
图9 摆尾位置流速分布图Fig.9 Water velocity distribution of tail-beat position
2.2 摆尾行为与流场耦合的动态规律
摆尾角度变化过程为进一步探索齐口裂腹鱼上溯过程中冲刺阶段摆尾角度的动态变化规律,分别计算10尾鱼上溯过程中摆尾角度波峰值的均值、波谷值的均值。提取摆尾角度波峰值、波谷值出现处的水流速度,并计算其均值。最终得到齐口裂腹鱼在上溯过程中,摆尾角度波峰值平均值与波谷值平均值随着流场改变的变化规律趋势图(图 10)。随着流速的增大,摆尾角度的波峰值与波谷值都呈现递减趋势,即摆尾角度数值与流速大小的耦合关系呈现负相关,但摆尾角度数值随流场变化的范围有限。在本试验条件下,波峰值最小值趋近于40°,波谷值最小值趋近于8°。
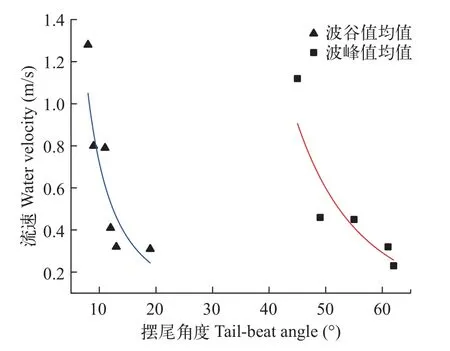
图10 摆尾角度随流场变化趋势图Fig.10 Trend of tail-beat angle with water flow field
摆尾模式为进一步分析摆尾行为与流速变化是否具有同步性,对上溯过程中摆尾角度与流速的强弱变化规律进行统计,其中数值增大视为强度增强,赋值为“1”;数值减小视为强度减弱,赋值为“0”。进一步统计得到摆尾角度与流速强弱变化同步率(表 2),数据显示,10尾鱼中有7尾鱼的同步率未超过50%,由此可见齐口裂腹鱼上溯过程中摆尾行为与流速变化的同步性较弱,即齐口裂腹鱼偏好在流速由大变小的区域摆尾冲刺加速。
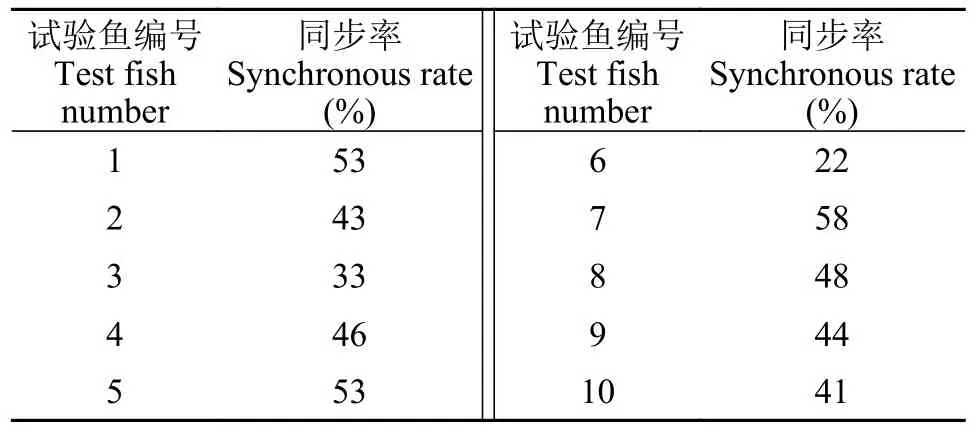
表2 摆尾角度与流速增减同步率Tab.2 Synchronization rate of tail-beat angle and water flow rate increase/decrease
采用排列组合的方式,将连续无间隔的3次强弱变化视为一组,各组之间无重复,统计得到10尾鱼中出现概率最高的摆尾模式(表 3)。综合分析表中数据,齐口裂腹鱼在上溯过程中喜好摆尾模式为“010”,即摆尾角度强度变化“弱强弱”的摆尾模式。
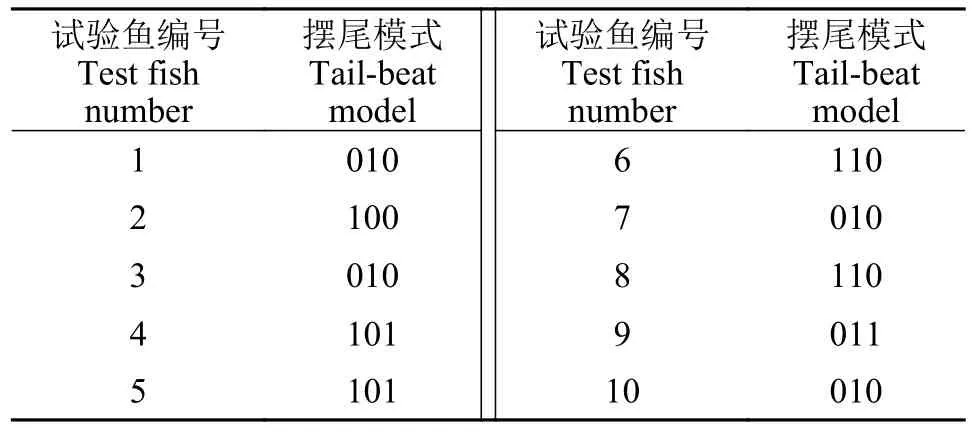
表3 十尾鱼最显著摆尾模式统计Tab.3 Statistics of the most significant tail-beat patterns of 10 fish
2.3 滑行距离与滑行方向
滑行流速系数为了进一步分析滑行阶段摆尾角度(θ)、滑行距离(L)和流速(v)三者之间的耦合规律,首先引入摆尾推进效率(η),它表示单位摆尾角度内鱼体前进的绝对距离,计算公式如式(4)所示。为进一步加入流速这个水动力学参数,最终引入滑行流速系数(α),它表示单位流速下摆尾推进效率的标准化值,计算公式如式(5)所示。汇总统计10尾鱼的滑行流速系数(α),结果表明(图11),当其数值为1.0—3.0时具有一定的代表性,从而定量得到了滑行阶段摆尾角度(θ)、滑行距离(L)和流速(v)三者之间的耦合关系。
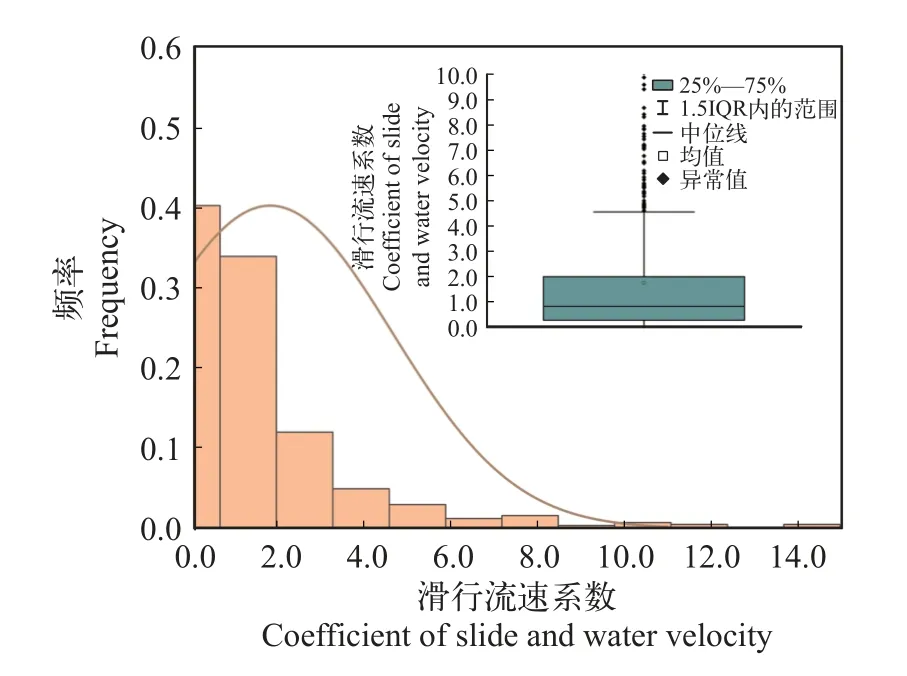
图11 滑行流速系数分布图Fig.11 Slide velocity coefficient distribution and box line diagram
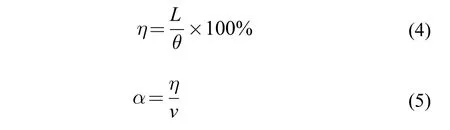
滑行方向与水流负方向夹角的偏好范围滑行阶段,鱼体保持近似直线的状态进行减速运动,直到再次冲刺摆尾加速。通过计算其滑行过程中滑行的绝对距离以及在流速负方向上的位移,可以计算“冲刺-滑行”行为行进的路程对水流负方向位移的贡献率,进而得到其每一次滑行的方向与水流负方向的夹角(表 4)。绘制10尾鱼滑行方向与水流负方向夹角的概率分布图(图 12),得到齐口裂腹鱼上溯过程中滑行方向与水流负方向夹角的偏好为40°—60°。
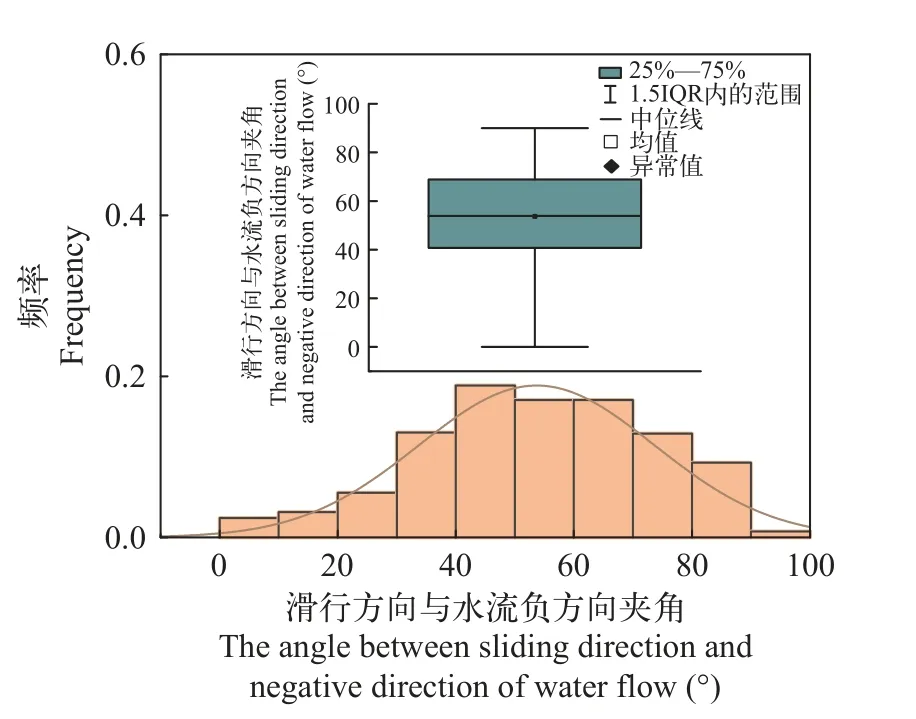
图12 滑行方向与水流负方向夹角分布图Fig.12 The angle between sliding direction and water flow negative direction distribution and box line diagram
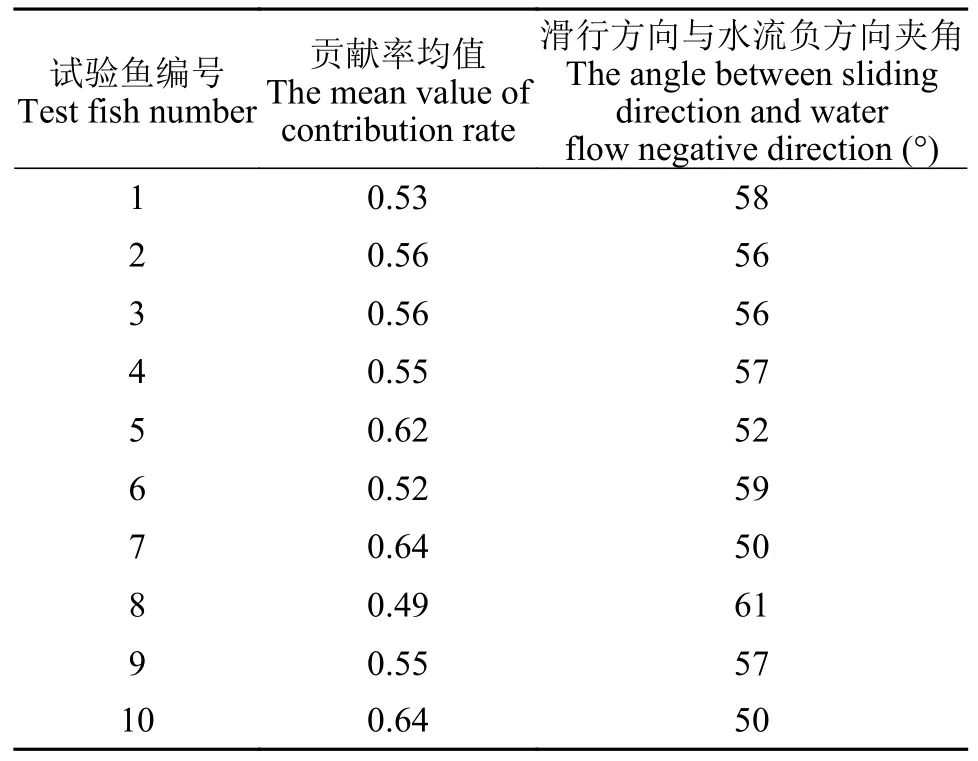
表4 贡献率均值及其对应滑行方向与水流负方向夹角Tab.4 The mean value of contribution rate and its corresponding angle between the glide direction and the negative direction of water flow
3 讨论
大坝已成为洄游鱼类游动的障碍,研究洄游鱼类的上溯行为有助于鱼道的设计[22]。鱼类能否通过鱼道是由游泳耐力、行为和动机综合决定的[23]。太平洋七鳃鳗(Lampetra japonicum),已被证明其耐力是其能否成功上溯的一个重要影响因素[24]。本研究对齐口裂腹鱼在上溯过程中的“冲刺-滑行”行为对水动力学条件的响应进行分析,为鱼道的设计提供支持。太平洋七鳃鳗已被证明上溯时优先选择低速通道[25];鳟通过涵洞时,低流速区的设计成为其能否成功通过涵洞的一个关键因素[26],本研究中成功上溯的试验工况里,较少出现选择在高流速区域进行上溯的情况,而多偏向于对于中低速流场的选择。因此鱼道设计时要充分考虑低流速区的塑造。在自然界中,大多数鱼类是通过尾鳍摆动对周围流体产生推力,从而获得流体的反作用力前进。鱼通过持续的摆尾,来保持身体的平衡,也能通过摆尾产生向前进的推力,提供动力来源。“冲刺-滑行”游泳需要通过大的摆幅,才能获得更多的能量,产生大的推力,使鱼能顺利地向前游[27]。研究其冲刺阶段的摆尾行为,对研究其上溯过程中的耗能情况具有重要意义。本研究表明,齐口裂腹鱼上溯过程中摆尾位置多为具有明显流速梯度的地方,这应该与其需要通过摆尾来获得更大的动力来越过流速障碍有关。
夏丹等[28]提出随着鱼摆尾频率和摆尾角度的增大,其游动速度、功率和效率均有所提高,但速度功率的比值有所降低。选择较小的摆尾频率和摆尾角度有利于减小功率,提高速度功率比,游动同样的距离,能量消耗也会明显降低[28]。该结论从一定程度上解释了本研究得到的随着流速增大,齐口裂腹鱼选择减小摆尾角度这一结果。Thandiackal等[29]提出鱼体总机械能的10%—20%是由水体对鱼体做的负功,且主要集中在鱼体前、中部区域,而不是尾部。由此推测水体对鱼体尾部做功应多为具有促进作用的正功。本研究得到齐口裂腹鱼在前进过程中偏好摆尾角度“弱强弱”的变化模式,可能是在水体做正功为主,负功为辅的情况下,通过尽可能地减小摆尾角度来更好地运用与节省能量。
建立鱼类游泳运动学参数之间的关系有助于建立鱼类生物学仿真模型,为传统过鱼设施设计提供依据[30]。本研究引入的滑行流速系数,可类比于经验系数。若已知流速,则可以反推出对应的摆尾推进效率的范围。若进一步得知摆尾角度,则可以推得该次摆尾后滑动距离的范围。在此基础上,可结合本研究中得出的另一个结果,即齐口裂腹鱼上溯过程中滑行方向与水流负方向夹角的偏好范围综合分析,即在已知摆尾角度和流场条件的情况下,可以为齐口裂腹鱼上溯轨迹的预测提供一定的研究思路与理论依据。
4 结论
本研究运用具有流速梯度的水槽创造非均匀流场条件,运用图像识别技术,量化计算其上溯全过程的游泳动力学指标。构建了新的指标体系,引入滑行流速系数、摆尾模式等。在此基础上做了新的量化分析,实现生态学与水动力学的耦合,进而得到“冲刺-滑行”行为对水动力学条件的响应规律。本研究表明,齐口裂腹鱼上溯过程中喜好在具有流速梯度处通过改变摆尾角度、摆尾频率等来适应非均匀流场。在本试验条件下,其喜好摆尾角度为25°—35°,喜好摆尾频率为2.5—3.5次/s,偏好流速为0.20—0.40 m/s。随着水流速度的增大,摆尾角度呈现逐渐减小的趋势,且齐口裂腹鱼偏好在流速由大变小的区域,进行摆尾冲刺加速,更趋向于摆尾角度变化为“弱强弱”的摆尾模式。滑行阶段引入滑行流速系数,量化表示摆尾角度、滑行距离和流速三者间的耦合关系,通过计算滑行距离对水流负方向上位移的贡献率,得到滑行方向与水流负方向夹角。研究表明,滑行流速系数的数值为1.0—3.0时具有代表性,齐口裂腹鱼对滑行方向与水流负方向夹角的偏好为40°—60°。本文利用多指标量化评价的方法,以复杂流场为背景条件,进一步满足过鱼设施建设需求。
在今后的研究中,在控制鱼体本身的特性、环境因子和能量代谢等因素基本一致的情况下,可进一步细化分析其上溯过程。运用滑行流速系数的概念,结合滑行方向与水流负方向夹角偏好等理论依据,建立鱼类上溯轨迹预测模型,并利用试验数据进行验证。本研究仅选择了上溯过程中的游泳动力学指标作为切入点,之后的研究还可以结合游动摆尾时速度和游泳加速度等,对其上溯过程的游泳策略进行更加详细的补充。
