花生紫色酸性磷酸酶基因家族的鉴定及表达分析
2022-07-08杜普旋刘浩胡冬秀陈小平洪彦彬李友国
杜普旋,刘浩,胡冬秀,陈小平,洪彦彬*,李友国
(1.华中农业大学,农业微生物学国家重点实验室,湖北 武汉,430070;2.广东省农业科学院作物研究所,国家油料作物改良中心南方分中心,广东省农作物遗传改良重点实验室,广东 广州,510640)
磷是植物生长发育过程中不可缺少的大量元素,它不仅在糖代谢、蛋白质代谢和脂肪代谢等生理代谢过程中起着重要作用,而且对提高植物抗逆性及增加作物产量有着重大影响。然而在大多数土壤中无机磷的含量远低于植物生长所需,大多数磷以有机形式或难溶性的磷酸钙、磷酸铁和磷酸铝的形式存在,难以直接被植物利用[1]。为了能在低有效磷环境中生存,植物产生了各种各样的适应性反应以提高磷利用效率,如改变根系结构、释放有机酸、诱导和分泌酸性磷酸酶等[2]。紫色酸性磷酸酶(Purple acid phosphatase,PAP)是金属磷酸酶超家族(Metallo-phosphatase superfamily)的成员,在植物组织中含量丰富,大多数PAP 是非特异性的,能水解一系列的磷酸酯,包括ATP、PEP、糖磷酸酯、单核苷酸和无机焦磷酸酯等[3]。PAPs含有一个双核金属离子活性中心,在哺乳动物中多为Fe(Ⅲ)-Fe(Ⅱ)中心,在植物中则为Fe(Ⅲ)-Zn(Ⅱ)或Fe(Ⅲ)-Mn(Ⅱ)中心,其特征性的粉红色或紫色源自双核中心内酪氨酸残基和三价铁离子之间的电荷转移跃迁[4]。植物中的PAP 可分为两类,大分子量PAP 和小分子量PAP。系统发育分析表明大分子量PAP与真菌和分枝杆菌中的PAP 亲缘关系较近,而小分子量PAP 与哺乳动物和细菌中的PAP 亲缘关系较近。大分子量PAP 常以二聚体形式存在,其单体分子量约为55 kDa,通常含有N 端和C 端两个结构域,而小分子量PAP 通常是分子量在35 kDa 左右的单体蛋白质,只有一个C 端结构域[5]。N 端结构域由两个β-折叠构成,每个β-折叠由三条反向平行的β-链组成。C 端结构域由两个β-α-β-α-β 结构组成的混合β-折叠结构构成,并且含有五个高度保守基序(DXG/GDXXY/GNH(D/E)/VXXH/GHXH),粗体为结合金属离子的七个保守氨基酸残基[6]。目前,PAP已经在多种植物中被鉴定和分离,拟南芥(Arɑbidopsis thɑliɑnɑ)、水稻(Oryzɑ sɑtivɑ)、大豆(Glycine mɑx)中分别含有29个、26个、35个PAP基因。
酸性磷酸酶(APase)的诱导和分泌被认为是在低无机磷条件下促进植物生长的重要策略,PAP 是一类重要的植物APase,可分泌到根际,利用有机磷促进植物生长发育[7]。大豆GmPAP14在有机磷(植酸)处理后强烈诱导表达,体外酶活性测定表明Gm-PAP14具有较高的植酸酶活性。此外,GmPAP14过表达增加了分泌型酸性磷酸酶和植酸酶活性,提高了植物对外源植酸的利用率,进而使植株磷含量上升,地上部分重量增加[8]。拟南芥的衰老叶片中AtPAP26的转录本和免疫反应性多肽显著积累,酸性磷酸酶活性增加了8 倍,而AtPAP26的T-DNA 插入突变体的衰老叶片中酸性磷酸酶活性显著降低,黑暗处理诱导条件下叶片衰老时间延迟了3 天,揭示了AtPAP26可以通过提高磷从衰老叶片到发育组织和种子的再利用来提高作物磷利用效率[9]。除了参与磷素的利用外,PAP 还参与其他的生理生化反应。GmPAP3的表达受盐胁迫和氧胁迫诱导却不受低磷胁迫诱导,可以通过减少活性氧的积累来应对非生物胁迫[10]。当进行NaCl、PEG 处理时,GmPAP3在烟草细胞中异位表达的结果与抗坏血酸的保护作用相似,都使死亡细胞百分比和活性氧积累减少;经NaCl、PEG、百草枯处理后拟南芥GmPAP3转基因植株的根伸长率显著高于野生型,脂质过氧化作用降低[11]。AtPAP2是第一个被证明调节碳代谢的紫色酸性磷酸酶,通过其C-末端疏水基序(TMD/CT)靶向叶绿体和线粒体,AtPAP2过表达拟南芥植株抽薹提前,种子产量增加,地上部的糖类(蔗糖和己糖)与三羧酸循环代谢产物含量升高[12]。Nt-PAP12过表达的烟草细胞中糖苷酶活性降低,纤维素和胼胝质合成酶的活性增强,进而促进了细胞壁的合成[13]。AtPAP23的启动子定位发现其在花中转录水平较高,而在根、茎、叶、花萼、角果、种子中未检测到表达,表明AtPAP23可能参与调节花的生长发育[14]。总而言之,现有的证据表明PAP 是多功能蛋白,无论在正常还是逆境条件下,它们都可能在植物的生长发育中发挥重要作用。
花生(Arɑchis hypogɑeɑL.)是重要的油料经济作物,主要种植于亚洲、非洲和美洲,种植面积超过2.5×106hm2,全球总产量超过4200 万吨,因此与大豆、棉籽、油菜籽和向日葵并列世界五大油料作物[15,16]。栽培种花生为异源四倍体(AABB 2n=4×=40),A 亚基因组来源于二倍体野生种花生A. durɑnensis,B 亚基因组来源于二倍体野生种花生A.ipɑensis[17]。花生的基础生物学研究相较于模式植物来说进展比较缓慢,但栽培种花生的全基因组测序为功能基因组学和花生品种改良奠定了基础[18]。土壤有效磷缺乏已经成为制约花生生长发育的主要因素,选育磷高效花生品种具有重要意义。鉴于植物PAP 在磷素利用中的潜在作用已被广泛研究,而目前对花生中PAP 的研究尚未见报道,因此本研究通过生物信息学手段从花生基因组数据库中鉴定AhPAPs基因家族成员,对候选基因的系统进化关系、基因结构、蛋白质保守结构域与理化性质以及组织特异性表达模式进行分析,为揭示AhPAP基因家族的功能提供理论基础。
1 材料与方法
1.1 AhPAP基因家族成员的鉴定
利用花生基因组数据库(https://www. peanutbase. org/)中的Gene & Gene Family 功能,以Purple acid phosphatase 为关键词检索AhPAP基因家族成员;然后从拟南芥数据库(https://www. arabidopsis.org/index.jsp)下载AtPAP基因家族28个成员的氨基酸序列,以28 个AtPAP氨基酸序列为模板在花生基因组数据库中进行Blastp 同源比对,将比对结果与通过关键词检索获得的AhPAP基因进行合并整理,并通过NCBI 在线网站CDD(https://www. ncbi. nlm.nih.gov/Structure/cdd/cdd.shtml)对蛋白保守结构域进行分析,剔除不含有金属磷酸酶结构域(MPP_PAP)的蛋白序列,筛选出花生基因组中的AhPAP候选基因序列。从花生基因组数据库中获取候选Ah-PAP基因的染色体位置信息,采用MG2C(http://mg2c. iask. in)在线网站绘制AhPAP基因的染色体定位图。
1.2 AhPAP基因家族系统发育关系及基因结构分析
使用MEGA-X 软件对拟南芥、水稻、苜蓿、花生PAP 的氨基酸序列进行比对,并采用Neighbor-Joining 方法对其构建系统进化树,各参数设定为:Possion model、Gamma Distributed、Pairwise deletion、Bootstrap Replications 设为2000。在花生基因组数据库中下载候选AhPAP基因家族成员的基因组DNA序列和mRNA序列,将mRNA序列提交到NCBI的ORFfinder 在线网站(https://www. ncbi. nlm. nih.gov/orffinder/)获得其CDS 序列,根据候选AhPAP基因的基因组DNA 序列和CDS 序列,利用在线网站GSDS 2.0(http://gsds. cbi. pku. edu. cn)对其基因结构进行分析。利用在线网站ExPasy(https://www.expasy.org/)预测AhPAP基因家族蛋白的分子量和等电点等理化性质。
1.3 AhPAP基因家族的表达特征分析
从花生基因组数据库中下载AhPAP基因家族成员在主茎叶、根、根瘤、花、雌蕊、雄蕊、营养茎尖、生殖芽尖、果针、荚果、种子等22个组织转录组数据的FPKM 值,利用TBtools 软件中的heatmap 功能对AhPAP基因在不同组织中的表达量进行聚类、绘制表达热图。以航花2 号花生为材料,分别取成熟期的根、根瘤、茎、叶、花、种子、果壳、外露果针等部位100 mg,利用RNAprep Pure 多糖多酚植物总RNA 提取 试 剂 盒(TIANGEN)提 取 总RNA。取1 µg 总RNA,使用FSK-101 反转录试剂盒(TOYOBO)进行cDNA 的合成,反应体系为20 µL。采用ABI StepOne Plus 系统与SYBR Premix ExTaq(TaKaRa)试剂进行荧光定量PCR 反应,每个样品设置3 个生物学重复。以根中的表达量为对照,利用2-ΔΔCT法计算相对表达量,内参基因为Ah18S[19]。
2 结果与分析
2.1 AhPAP基因家族成员的鉴定
通过关键词检索和同源比对从花生基因组数据库中获得了59个AhPAP基因,利用NCBI在线分析工具CDD分析其蛋白保守结构域,剔除20个不含有金属磷酸酶结构域的蛋白序列,最终鉴定得到了39个AhPAP基因家族候选基因。采用MG2C 在线网站对39个AhPAP基因进行染色体定位(图1),结果显示:大多数AhPAP基因对称分布于两个亚基因组内,13号染色体上含有6个AhPAP基因,3号染色体上含有5个AhPAP基因,1号和11号染色体上各含有4个Ah-PAP基因,6号、8号、16号、18号染色体上各含有3个AhPAP基因,2号、4号、7号、10号、12号、14号、17号、20号染色体上每条只含有1个AhPAP基因,而5号、9号、15号、19号染色体上不含有AhPAP基因。
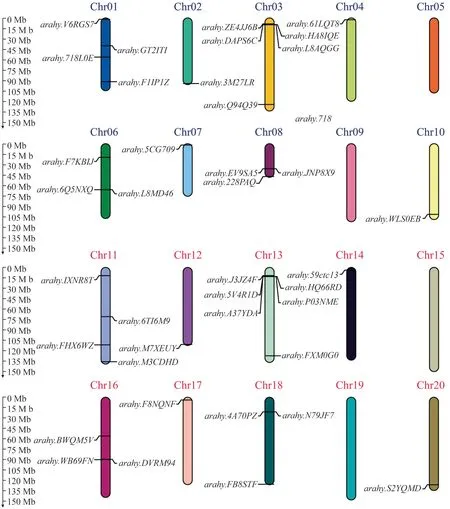
图1 AhPAP基因家族成员的染色体定位Fig.1 Chromosome location of AhPAP gene family
2.2 AhPAP基因家族的系统发育关系分析
为了分析AhPAP基因家族的进化关系,将其与拟南芥、水稻、苜蓿PAP 共74 个蛋白序列(花生39个、拟南芥28个、水稻5个、苜蓿2个)构建系统发育树(图2),结果表明它们可分为四组(GroupⅠ、Ⅱ、Ⅲ和Ⅳ),Group Ⅳ又可分为两个亚组(Group Ⅳa 和Ⅳb)。GroupⅠ包含9 个序列,GroupⅡ包含15 个序列,GroupⅢ和Ⅳ都包含25 个序列。GroupⅠ中含有8 个AtPAP基因家族成员,而AhPAP基因家族成员只有一个,而且它们之间的系统进化距离较远,ɑrɑhy. GT2ITI单独形成一个分支,推测该组成员在相应物种中的进化程度比较高。
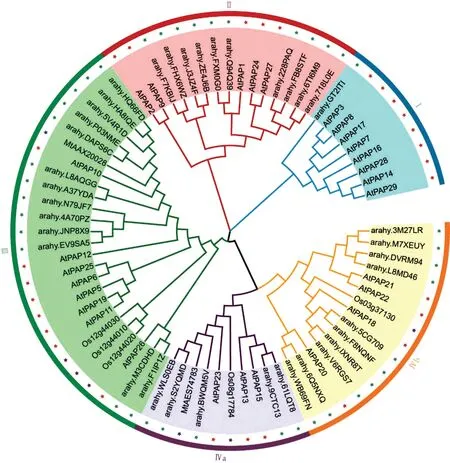
图2 PAPs基因家族系统进化分析Fig.2 Phylogenetic analysis of PAPs gene family
2.3 AhPAP基因家族保守基序及理化性质分析
使用MEGA-X 软件将39 个AhPAP基因家族成员的氨基酸序列比对,然后利用Highlight conversed sites 功能统计这些序列中5 个保守基序(DXG/GDXXY/GNH(D/E)/VXXH/GHXH)的存在状态,连同分子量和等电点等理化性质列于表1。从表中可以看出AhPAP基因家族的氨基酸长度为205~905,分子量为21.92~102.09 kDa。等电点为4.45~8.64,ɑrɑhy. DVRM94、ɑrɑhy. EV9SA5、ɑrɑhy. M3CDHD、ɑrɑhy.N79JF7的等电点大于7,为碱性蛋白质;其余蛋白的等电点均小于7,为酸性蛋白质。39 个AhPAP基因家族成员中有29 个具有上述5 个高度保 守 基 序,ɑrɑhy. 3M27LR、ɑrɑhy. F1IP1Z、ɑrɑhy.M7XEUY具有4 个保守基序,ɑrɑhy. 61LQT8、ɑrɑhy.BWQM5V、ɑrɑhy.IXNR8T、ɑrɑhy.N79JF7具有3 个保守基序,ɑrɑhy. F7KBIJ具有2 个保守基序,ɑrɑhy.FHX6WZ、ɑrɑhy.GT2ITI具有1个保守基序。

表1 AhPAP基因家族保守基序及理化性质Tabel 1 Conserved motifs and physicochemical properties of AhPAPs gene family
2.4 AhPAP基因家族的基因结构分析
使用TBtools 软件将AhPAP进化树与基因结构分析结果一起进行可视化作图(图3)。结果显示AhPAP基因的外显子数量存在较大差异,39 个基因分别含有1~20 个外显子,其中含有8 个外显子的基因数目最多。ɑrɑhy.FXM0G0含有20个外显子,ɑrɑhy.DAPS6C含有8 个外显子,而ɑrɑhy.5CG709只含有1 个外显子。此外大部分基因可聚类成两两一组,同一组内两个AhPAP基因具有相似的结构,如ɑrɑhy. 228PAQ与ɑrɑhy. FB8STF都含有12 个外显子、ɑrɑhy. S2YQMD与ɑrɑhy. WLS0EB含有9 个外显子、ɑrɑhy. A37YDA与ɑrɑhy. L8AQGG含有8 个外显子等。ɑrɑhy. F7KBIJ只存在5’非编码区,ɑrɑhy.ZE4J6B只存在3’非编码区,而ɑrɑhy.GT2ITI基因两端都不具有非编码区,其余基因均含有5’非编码区和3’非编码区。

图3 AhPAP基因家族的基因结构分析Fig.3 Gene structure anlysis of AhPAP gene family
2.5 AhPAP基因家族的表达特征分析
为了初步了解AhPAP基因家族的功能,从Peanutbase 中下载鉴定出的39个AhPAP基因在22个组织中的表达量数据,使用TBtools 软件绘制表达热图(图4),图中灰色表示未检测到基因的表达。结果显示,这些基因呈现出不同的表达特征。ɑrɑhy.A37YDA、ɑrɑhy.L8AQGG在雌蕊中表达量较高,在其他组织中表达量低或未检测到表达。ɑrɑhy.P03NME、ɑrɑhy. DAPS6C在根瘤中有较高表达量,在其他组织中表达量较低。ɑrɑhy.6Q5NXQ、ɑrɑhy.WB69FN、ɑrɑhy. DVRM94、ɑrɑhy. J3JZ4F、ɑrɑhy.ZE4J6B、ɑrɑhy. F7KBIJ、ɑrɑhy. L8MD46等在花中高度表达,其中ɑrɑhy. 6Q5NXQ、ɑrɑhy. WB69FN、ɑrɑhy.DVRM94、ɑrɑhy.J3JZ4F在雌蕊中表达量也比较高 。ɑrɑhy. 3M27LR、ɑrɑhy. M7XEUY、ɑrɑhy.JNP8X9、ɑrɑhy.N79JF7等在果壳、荚果中表达量显著高于其他组织。ɑrɑhy. 6TI6M9、ɑrɑhy. 718L0E在营养茎尖、生殖芽尖、根、根瘤、荚果中表达量较高,在根瘤中的表达量最高。ɑrɑhy. 4A70PZ、ɑrɑhy.EV9SA5在根、种子中表达量高于其他组织。
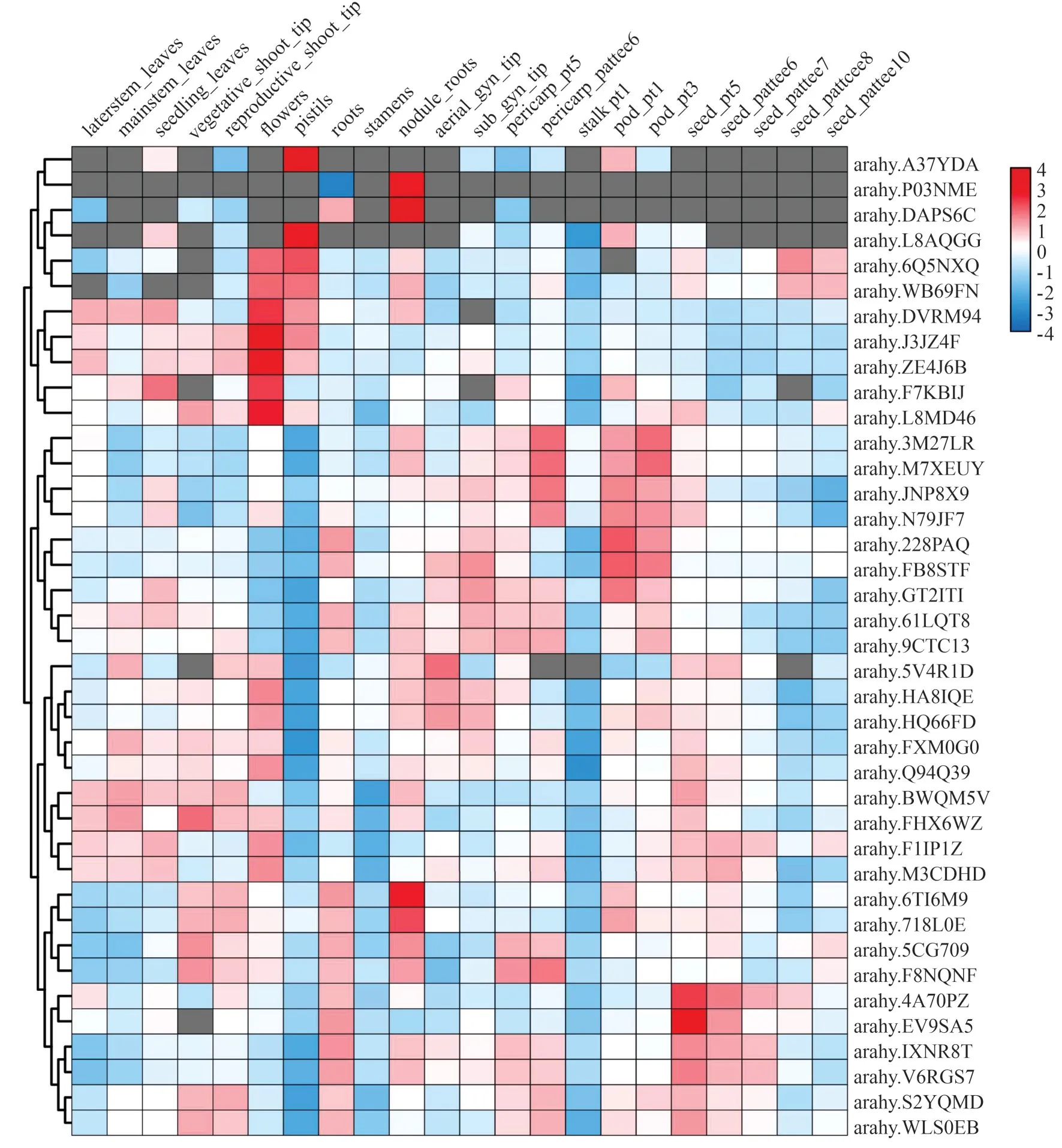
图4 AhPAP基因家族的表达模式Fig.4 Expression pattern of AhPAP gene family
2.6 AhPAP基因表达量验证
为了验证转录组数据的可靠性,对花生不同组织(根、根瘤、茎、叶、花、种子、果壳、外露果针等)中ɑrɑhy.718L0E、ɑrɑhy.DVRM94、ɑrɑhy.HQ66FD、ɑrɑhy. V6RGS7四个AhPAPs基因家族成员进行qRTPCR分析,利用TBtools软件的Super Heatmap Browser 功能对每个基因的相对表达量绘制热图(图5),所用引物列于表2。结果表明,ɑrɑhy. 718L0E、ɑrɑhy. DVRM94、ɑrɑhy. HQ66FD、ɑrɑhy. V6RGS7四个基因分别在根瘤、花、外露果针、种子中的相对表达量最高,与Peanutbase中的RNA-seq数据基本一致。

表2 qRT-PCR引物Tabel 2 Primers used for qRT-PCR
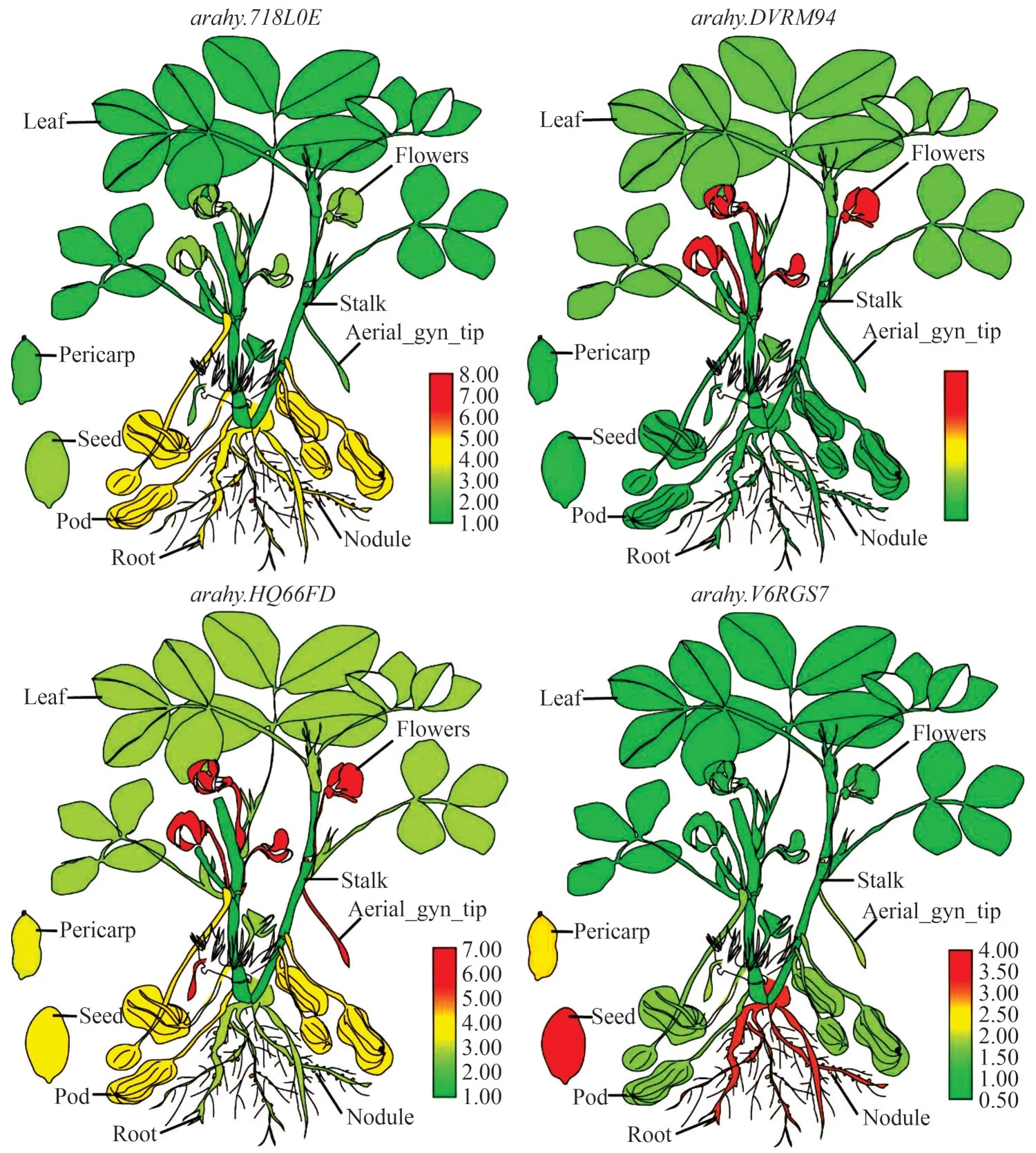
图5 AhPAPs的组织表达量分析Fig.5 Analysis of tissue expression of AhPAPs
3 讨论
当今世界上几乎所有的粮食生产都依赖于磷矿作为单一的磷肥来源,20 世纪中期加速使用开采的磷矿,加上使用哈伯-博施法(Haber-Bosch process)合成氮肥,极大地提高了全球作物产量。但是,磷素是通过农业从矿山向海洋单向流动的,我们正在迅速消耗一种关键但有限的资源,同时造成了世界河流和海洋的污染[20]。面对世界人口的增加、全球磷矿储量的减少、豆科植物对磷的需求相对较高等现状,提高豆科植物对磷的利用效率是一个全球性的挑战[21]。PAP 参与植物的多种生理功能,尤其是对磷缺乏的适应。在低磷环境下植物PAP 的转录丰度和蛋白质水平显著增加,进而将细胞内的磷活化再利用并且分解土壤环境中的有机磷供植物吸收利用[22]。随着基因组数据库的不断丰富,PAP基因已在多种植物中被分离鉴定,并证实有些PAP基因还参与碳代谢、调节花的生长发育和细胞壁合成等过程[23]。因此,AhPAP基因家族成员的鉴定和分离有助于揭示花生低磷适应机制,为选育磷高效花生品种提供候选基因,对提高花生的磷素利用效率具有重要意义。
本研究从花生全基因组中鉴定出39 个AhPAP候选基因,其中29 个基因含有完整的5 个保守基序和7 个保守氨基酸残基,其余10 个基因虽然缺失了部分保守基序,但由于其C 端含有金属磷酸酶结构域,因此仍把它们归为AhPAP基因家族成员。对AhPAP基因家族成员理化性质分析发现,其中33 个成员的分子量大于35 kDa,因此花生中的PAP 大多数为大分子量PAP。而且有35 个成员的等电点在4.45~6.98 之间,这与PAP 在酸性条件下能够催化磷酸单酯或酸酐水解的特性相符。以花生、拟南芥、水稻、苜蓿共74 个PAP 蛋白构建的系统发育树可分为四组(GroupⅠ、Ⅱ、Ⅲ和Ⅳ),每组中都含有来自不同物种的PAP,并没有因为物种差异而各自聚为一类,说明不同物种间的PAP 可能具有相似功能。AtPAP15具有植酸酶活性,通过调节叶片抗坏血酸的合成来改变植物对盐和氧化胁迫的抗性[24],ɑrɑhy. 61LQT8、ɑrɑhy. 9CTC13与AtPAP15的系统进化距离最近,推测它们可能具有相似的抗逆功能。AhPAP基因家族中含有17 对旁系同源基因,除ɑrɑhy. JNP8X9、ɑrɑhy. EV9SA5为同一基因的两个拷贝之外,其余的16对基因多数为对称分布于不同亚基因 组 内 的 同 一 基 因。 而ɑrɑhy. GT2ITI、ɑrɑhy. 5V4R1D、ɑrɑhy. N79JF7、ɑrɑhy. 4A70PZ、ɑrɑhy.BWQM5V仅有一个基因存在于亚基因组内,在各自的分组内单独形成一个分支,可能在进化中发生了正向选择。基于转录组数据与qRT-PCR 对AhPAP基因家族的表达特征进行了分析,结果显示一些基因的表达具有组织特异性。ɑrɑhy.6Q5NXQ、ɑrɑhy.WB69FN、ɑrɑhy. DVRM94、ɑrɑhy. J3JZ4F在花和雌蕊中的表达量显著高于其他组织,可能具有调节花的 生 长 发 育 的 功 能。ɑrɑhy. P03NME、ɑrɑhy.DAPS6C、ɑrɑhy. 6TI6M9、ɑrɑhy. 718L0E在根瘤中表达量相对较高,推测它们参与根瘤中的磷代谢与共生固氮过程。AhPAP基因家族的具体功能还需要大量实验验证。
目前,PAP 的功能已经在拟南芥、水稻、大豆等植物中得到了广泛探究,而花生作为我国重要的油料经济作物,却鲜有关于PAP 的报道。本研究在花生全基因组水平上对AhPAP基因家族进行了鉴定,并对其染色体定位、系统进化关系、保守结构域、基因结构、蛋白理化性质、组织表达模式等进行了生物信息学分析,以期为深入研究AhPAP基因家族在花生中的功能及调控机制奠定基础。
