水杨酸对低温胁迫下菜豆种子萌发及抗寒性的影响
2022-07-04孙梦笛冯国军刘大军闫志山杨晓旭
孙梦笛,刘 畅,冯国军,刘大军,闫志山,杨晓旭
(黑龙江大学 现代农业与生态环境学院,哈尔滨 150080)
0 引 言
菜豆(PhaseolusvulgarisL.)也称芸豆,豆科,菜豆属[1],是短日照植物,喜温,不耐冻。如今菜豆在我国大面积地种植,菜豆的品种资源非常丰富,属于我国主要的蔬菜种类之一,也是重要的食用豆类之一。 菜豆种子萌发开始温度为8 ℃左右,适宜生长温度为20~25 ℃,种子萌发适应的地温为10~12 ℃[2]。
植物在生长过程中,经常会受到低温的影响,降低植株的出苗率,导致其产量迅速下降。种子的萌发是其生长过程中的重要阶段,萌发过程所发生的生化反应都需要适宜的温度,低温会导致种子的萌发受到抑制,贮藏物质水解迅速,发芽率降低,过氧化物酶的功能减退,破坏了活性氧原有的平衡,导致细胞代谢紊乱。因此,研究在低温胁迫下对菜豆种子的影响,对菜豆的生产具有重要的意义[3]。
水杨酸作为一种小分子酚类物质,也是植物内源激素,在植物逆境条件下具有较强的生理作用。对于低温的逆境条件,水杨酸能够进一步缓解低温胁迫对植物生长造成的负面影响。近年来,关于水杨酸在低温胁迫下的研究表明:经外源水杨酸处理的植物抗寒性有所提高,如茄子[4],玉米[5],棉花[6]等,但是水杨酸在低温胁迫条件下对菜豆种子萌发的影响少有报道。本文以低温敏感的“热那亚”为试验材料,研究在低温胁迫条件下,施加外源物质水杨酸对菜豆种子萌发以及生理生化特征的影响,为菜豆低温胁迫的有效防御提供参考。
1 材料与方法
1.1 试验材料
本研究选择低温敏感型的“热那亚”菜豆为试验材料,由黑龙江大学呼兰校区园艺学团队提供。
1.2 试验方法
1.2.1 种子萌发试验
本研究选取低温敏感型的“热那亚”品种。选择大小一致、成熟饱满的种子,用10%的次氯酸钠消毒10 min,再用蒸馏水冲洗3~5次,吸干水后置于带有滤纸的培养皿中,每皿50粒种子。在对菜豆种子进行低温胁迫实验时,发现低温处理12 h种子的发芽率较高,因此设置3个处理组,每组进行3次重复试验。①CK:25 ℃+30 mL蒸馏水,置于25 ℃培养箱培养;②LT:4 ℃+30 mL蒸馏水,置于4 ℃培养箱中培养12 h进行低温胁迫处理,再置于25 ℃培养箱中继续培养;③LT+SA:4 ℃+30 mL,2.5 mg·L-1SA,置于4 ℃培养箱中培养12 h进行低温胁迫处理,再置于25 ℃培养箱中继续培养。
以胚根伸出种皮2 mm作为发芽标志,计算发芽率、发芽势、发芽指数,第4天测量胚根胚轴长度。发芽势=(发芽初期规定日期内发芽数/供试种子数)×100%,发芽率=(发芽末期规定日期内发芽数/供试种子数)×100%,发芽指数=不同时间的发芽数/对应的发芽天数。
1.2.2 生理指标的测定
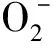
1.3 生理指标测定方法

1.4 数据处理
试验数据均采用DPS软件进行差异显著性分析,试验图表均采用Excel软件绘制。
2 结果分析
2.1 外源水杨酸对低温胁迫下菜豆种子萌发的影响
外源水杨酸对低温胁迫下菜豆种子萌发的影响见表1。由表1可见,与CK相比,在低温胁迫下,菜豆种子的发芽率,发芽势,发芽指数分别降低了53.43%,58.74%,61.19%;在低温下施加水杨酸,与LT相比分别提高了94.13%,113.58%,117.16%;与CK相比,LT+SA处理分别降低了9.59%,11.89%,15.70%。由此表明:在低温胁迫下,菜豆种子的萌发受到抑制,但施加外源SA能够有效缓解低温对菜豆种子萌发的抑制,菜豆种子发芽率,发芽势,发芽指数显著提高,但未恢复到常温组原有的发芽水平。

表1 外源水杨酸对低温胁迫下菜豆种子萌发的影响Table 1 Effects of exogenous salicylic acid on Phaseolus vulgaris seeds under low temperature stress
2.2 外源水杨酸对低温胁迫下菜豆种子胚轴胚根的影响
外源水杨酸对低温胁迫下菜豆种子胚轴胚根的影响见表2。由表2可见,CK和LT处理的胚轴长差异显著,但胚根长差异不显著,在低温条件下与CK相比,胚轴,胚根长分别降低了36.82%,17.68%。因此,低温胁迫抑制了菜豆种子的生长。LT+SA组的胚轴和胚根长与LT相比分别提高了72.93%,28.83%,表明施加外源SA促进菜豆种子胚的伸长。

表2 外源水杨酸对低温胁迫下菜豆种子胚轴胚根的影响Table 2 Effects of exogenous salicylic acid on Phaseolus vulgaris seeds under low temperature stress
Note: The averages and standard errors are from 3 independent experiments. Different lowercase letters in each column indicate significant differences at the 0.05 level.
2.3 外源水杨酸对低温胁迫下菜豆种子抗氧化酶活性的影响
低温胁迫下SA对菜豆种子抗氧化酶活性的影响见图1~图3。由图1可见,LT组处理的POD活性明显低于CK组,经SA处理的POD活性提高,并且随着处理时间的变化逐渐升高,与LT比,POD活性在12、24、36、48 h分别提高101.33%,70.82%,17.28%,20.65%,且差异显著。

图1 低温胁迫下SA对菜豆种子过氧化物酶活性的影响Fig.1 Effects of SA on POD activity of Phaseolus vulgaris seeds under low temperature stress
由图2可见,在4个不同的时间段中,低温条件CAT活性受到抑制,施加外源SA使CAT活性分别上升了69.84%,33.33%,27.91%,37.78%。并且施加外源SA的CAT活性高于常温对照组,在36 h和48 h时差异显著。

图2 低温胁迫下SA对菜豆种子过氧化氢酶活性的影响Fig.2 Effects of SA on CAT activity of Phaseolus vulgaris seeds under low temperature stress
由图3可见,在低温处理下SOD活性降低,与CK相比在36 h显著降低;与LT相比,SA处理下的SOD活性略微提高,均高于对照组CK且在12、24、48 h差异显著。

图3 低温胁迫下SA对菜豆种子超氧化物歧化酶活性的影响Fig.3 Effects of SA on SOD activity of Phaseolus vulgaris seeds under low temperature stress
由此说明施加水杨酸可以通过提高抗氧化酶的活性来实现保护生物膜的目的,进一步减少了低温对膜系统的伤害,提高菜豆种子的抗寒能力。
2.4 外源水杨酸对低温胁迫下菜豆种子MDA含量和产生速率的影响
MDA是判断植物膜结构被损害程度的标准之一。低温胁迫下SA对菜豆种子丙二醛含量的影响见图4。由图4可见,低温条件明显提高了菜豆种子的MDA含量,随着处理时间的延长,MDA含量逐渐升高,与CK对比,分别提高了44.14%,4.64%,19.44%,35.89%,且差异显著。施加外源水杨酸减少了MDA在低温下的积累,与LT对照比在12、24、36、48 h分别降低了13.02%,1.38%,7.89%,9.80%;且在12 h和48 h时差异显著,在24 h和36 h时差异不显著。

图4 低温胁迫下SA对菜豆种子丙二醛含量的影响Fig.4 Effects of SA on MDA content of Phaseolus vulgaris seeds under low temperature stress

图5 低温胁迫下SA对菜豆种子产生速率的影响Fig.5 Effects of SA on production rate of Phaseolus vulgaris seeds under low temperature stress
因此,低温下外源水杨酸可以降低菜豆在冷胁迫时自由基和活性氧的含量,避免积累过多的MDA,缓解了低温对膜系统的伤害程度。
2.5 外源水杨酸对低温胁迫下菜豆种子脯氨酸和可溶性蛋白含量的影响
低温胁迫下SA对菜豆种子脯氨酸和可溶性蛋白质含量的影响见图6,图7。由图6,图7可见,在4 ℃低温对照组,脯氨酸和可溶性蛋白的含量随着处理时间的延长,均呈现先升高后下降的趋势,在36 h达到最大值,与CK比分别提高了5.77%,6.46%。在低温下加入外源SA,与LT处理比增加了脯氨酸和可溶性蛋白的积累,其中,脯氨酸在不同的时间段分别增加了11.85%,6.64%,4.77%,6.24%,可溶性蛋白分别增加13.82%,9.85%,2.41%,13.84%。

图6 低温胁迫下SA对菜豆种子脯氨酸含量的影响Fig.6 Effects of SA on Proline content of Phaseolus vulgaris seeds under low temperature stress

图7 低温胁迫下SA对菜豆种子可溶性蛋白含量的影响Fig.7 Effects of SA on Soluble protein content of Phaseolus vulgaris seeds under low temperature stress
由此表明:菜豆种子在低温冷害下,施加水杨酸促进脯氨酸和可溶性蛋白含量的积累,进而有效地保护种子免受低温的伤害。
3 讨 论
由已报道的实验研究表明水杨酸常作为一种信号调节因子,不仅参与植物的各种胁迫反应,而且还会促进植物的生长,改善植物的生理生化特征[11]。通常水杨酸通过充当抗氧化剂来清除植物体内的活性氧,并且可以激活植物的抗氧化系统来直接抵御逆境给植物造成的氧化破坏影响[12]。在低温条件下,植物出现不同的胁迫反应,损害生物膜的结构,影响膜的透性,植物生理生化也发生不同程度的变化。本实验低温环境明显抑制了菜豆种子的萌发,施加外源水杨酸能够抵御低温冷害对菜豆种子造成的伤害,增强菜豆种子自身的抗寒特性。这与Diao Q N等[13]试验结果是一致的。
低温条件下,会使细胞膜受到伤害,发生膜脂过氧化反应最终产生丙二醛。丙二醛作为判断植物受到逆境危害程度的重要指标之一,与种子的活力密切相关,一般情况下,MDA的含量高,表明:种子受到一定程度的伤害导致种子发芽能力降低。在本实验中,低温逆境抑制种子的萌发,种子发芽率,发芽势和发芽指数有所下降,而MDA的含量有所上升,这与王宏燕等[14]研究情况相同。通过外源水杨酸的处理,能够抑制膜脂过氧化现象的产生,降低了MDA的含量,以此来减少低温逆境对菜豆种子带来的伤害,保持细胞膜的稳定性,提高菜豆种子在萌发阶段的抗寒能力。
脯氨酸作为一种渗透调节物质,在诸多胁迫环境中均有表现,同样在低温的条件下也积累游离的脯氨酸。在植物体内,脯氨酸能够加强蛋白质的水合作用,稳定细胞的结构,促进可溶性蛋白的含量的积累,保护细胞不受逆境的伤害。可溶性蛋白也是衡量植物抗寒性强弱的一种指标,其含量的提高表明植物对低温胁迫的适应性强,赵萌[15]认为可溶性蛋白的含量与植物抗寒的能力呈正相关。研究中,低温下的菜豆种子积累少量的脯氨酸和可溶性蛋白,但SA的添加提高了菜豆种子中脯氨酸和可溶性蛋白含量,进一步提高了菜豆种子在萌发过程中的抗寒性,该结果在玉米种子[16],扁稽豆种子[17]上早有报道。
活性氧在低温逆境的诱导下产生速率加快,导致种子的萌发受到抑制,为了防止活性氧带来的伤害,种子内形成保护酶系统来抵抗植物面临的逆境条件,保护酶系统主要包含POD,SOD,CAT 3种酶,本实验与常温对照比在低温下的保护酶的活性均降低,施加SA后可以提高菜豆种子内各个部位清除活性氧的能力,导致菜豆种子的POD和CAT活性显著提高,而SOD有略微的提高。
4 结 论
在低温逆境下,菜豆种子的发芽率、发芽势、发芽指数分别降低了53.43%,58.74%,61.19%;通过外加水杨酸分别提高了94.13%,113.58%,117.16%。与LT对照比,外源SA的施加减少了膜的损害,使菜豆种子MDA含量在12、24、36、48 h分别降低了13.02%,1.38%,7.89%,9.80%。脯氨酸在不同的时间段分别增加了11.85%,6.64%,4.77%,6.24%,可溶性蛋白分别增加了13.82%,9.85%,2.41%,13.84%。抗氧化酶作为ROS的清除系统之一,外源SA的处理增强了抗氧化酶对细胞膜的保护,提高抗氧化酶的活性,缓解低温逆境对菜豆种子的伤害。由此表明:低温胁迫打破了菜豆种子内部原有的动态平衡,导致氧代谢失调,积累大量的活性氧及MDA,降低抗氧化酶的活性,使细胞膜的透性发生改变,影响菜豆种子的正常萌发。而水杨酸作为植物生长过程中的调节物质,能够进一步改善菜豆种子在低温胁迫下的萌发情况,提高菜豆种子的抗氧化酶活性,同时减少了MDA和活性氧的积累,保护细胞膜的稳定性,维持菜豆种子内原有的平衡状态,能够有效抵御低温胁迫对菜豆种子带来的伤害,使菜豆种子在萌发过程中对低温环境产生一定的适应能力,提高其抗寒性。
