lnvestigating the specificity of endothelin-traps as a potential therapeutic tool for endothelin-1 related disorders
2022-06-21ArjunJainKristofBozovicarVidhiMehrotraTomazBratkovicMartinJohnsonIraJha
lNTRODUCTlON
Endothelin-1 (ET-1) is a vasoactive peptide synthesized and secreted by a diverse range of cells, and thus implicated in signaling events in a wide variety of target tissues[1]. ET-1 plays a key role in physiological functions. However, supraphysiological levels of ET-1 induce pathology and are implicated in a host of different diseases, including cardiovascular disease[2-4], neurodegenerative disorders[5-8], chronic kidney disease, different cancers, such as prostate cancer[1,9,10], pregnancy disorders like preeclampsia[7,11], as well as diabetes[8,12,13]. Given that a key feature of these diseases is elevated ET-1 Levels, one proposed strategy of therapeutic intervention is to target the increased levels of ET-1. To this end, we have created ET-traps, molecular constructs that bind and sequester increased levels of endogenous ET-1.
Diabetes is a serious metabolic complication that affects about more than 7% of the world population[14]. An increase in different extracellular matrix (ECM) proteins has been found to be a key pathological factor of diabetes[15,16]. The study by Jain
[12] found an increase in collagen 4α1 and fibronectin both at the mRNA and protein levels. This increase was found in heart and kidney tissues and was found to be ET-1 dependent[12]. In addition, the increase in ECM proteins due to high glucose levels was found to be mediated
ET-1[17]. ET-1 Levels are in fact increased in patients with diabetes compared with control subjects[18,19]. Accordingly, our previous
work confirmed the ET-traps to have an efficacious effect on cells treated with a pathological dose of ET-1, as well as those treated with pathologically high glucose (25 mmol/L)[12]. We also established the proof-of-concept (PoC) for ET-traps as a therapeutic in the diabetes disease space at the
level. The use of ET-traps gave a significant reduction in different markers of diabetes disease pathology, which suggested the ET-traps could be considered a therapeutic for diabetes with a novel mechanism of action. Importantly, the ETtraps were found to be non-toxic at the proposed therapeutic concentration both
and
[12,13].
ET-1
ET-1 exerts its effects by binding to the endothelin A and B receptors, two highly homologous cellsurface proteins that belong to the G-protein-coupled receptor superfamily[20]. The two receptors share about 60% similarity at the level of primary structure[1],
both receptors exhibit a high polypeptide sequence identity with each other. Nevertheless, the two receptors show a clear distinction in ligand binding selectivity based on their ligand-binding domains.
As we walked home, pushing the bike as we made our way along the sidewalk, she asked me about a conversation she d overheard7 me having with my mother the night before.
Orry
[21] constructed a model of interaction of the ET-1 peptide with the endothelin A receptor, where ET-1 makes contacts with both the N-terminal receptor domain and two different extracellular loops (ECL).
Further, amino acids of the C-terminal and residues in the third intracellular loop are important for ET-1 binding[22]. In this study, we performed binding affinity experiments to ascertain that our selected ET-traps bind specifically just to ET-1.
Therefore, both the ET-1 paralogs (ET-2 and ET-3) are essential for different physiological processes and so it is important that any ET-1 sequestering agent selectively targets ET-1 and hence the problems associated with increased expression of ET-1 to avoid disrupting the remaining processes of the ET system.
Homologs of ET-1
The ETs are a family of potent vasoactive peptides. ET-1 has two paralogs in the ET family; ET-2 and ET-3[23]. ET-1 was identified by Yanagisawa
[24] in 1988. A year later, 2 homologs of ET-1 were discovered; ET-2 and ET-3[25].
ET-2 is a peptide encoded by the
gene located on chromosome 1 in humans[26]. ET-2 has a key role in ovarian physiology[25]. Previous research findings have also revealed that ET-2 is critical for the growth and survival of postnatal mice and plays important roles in energy homeostasis, thermoregulation, and the maintenance of lung function[26].
ET-3 is a peptide that in humans is encoded by the
gene[27]. The active peptide is a ligand for ET receptor type B (EDNRB). The interaction of this ET with EDNRB is essential for development of neural crest-derived cell lineages, such as melanocytes and enteric neurons[28].
My pretty girlfriend had a failed marriage in the past. After divorce, her parents shed the deepest protection and care to her by helping5 her attend to the child and offering financial aids. Her parents’love made her pull herself together and forgot the man who had hurt her before.
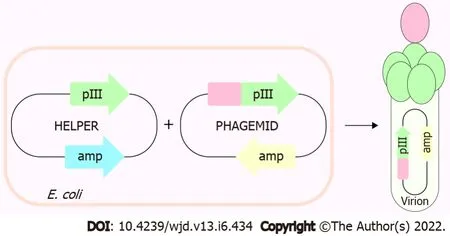
Phage display is a powerful technique commonly used today to identify different protein-protein interactions. We displayed individual ET-traps in a monovalent setting (
3 + 3 display type[32]) to prevent avidity effects on binding to ET-1. The cMyc-tag peptide present in the linker region that tethers ET-traps to the anchoring phage coat protein p3 allows for assessment of constructs’ display levels by phage ELISA against anti-cMyc antibody. These were, in turn, used to normalize signals from phage ELISA where binding of ET-traps to biotinylated ET-1 was analyzed (Figure 2).
And as she drank thus, and leaned right over the water, the rag containing the three drops of blood fell from her bosom11 and floated down the stream, and she in her anxiety never even noticed her loss
MATERlALS AND METHODS
Sub-cloning ET-traps into pIT2 phagemid vector
Previous work identified a strong binder to ET-1[31]. In this study, we tested different sequence combinations of ET-traps that could also bind ET-1. We performed phage display experiments to ascertain this.
Digested inserts were ligated into pIT2 phagemid vector and chemically competent
TG1 were transformed with the resulting recombinant phagemids with the heat-shock method. Phages were amplified and rescued by superinfection with KM13 helper phage. PEG/NaCl was used to precipitate and isolate phage clones which were spectrophotometrically quantified with NanoDrop 1000. Phage titers were calculated using equation 1 (derived by Day and Wiseman)[30] and subjected to phage enzyme-linked immunosorbent assay (ELISA).
Verifying ET-traps construct display
MaxiSorp microtiter plate (Nunc) wells were coated with anti-cMyc antibodies (1 µg/mL in PBS) overnight at 4 °C. Wells were blocked with 5% skimmed milk and 100 µL of 5 × 10
phage clone virions in 0.5% milk/0.1% PBST were added and incubated for 1 h with gentle agitation. After extensive washing, bound phages were detected with anti-M13 monoclonal antibodies conjugated with horseradish peroxidase (GE Healthcare) and chromogenic substrate (3,3’,5,5’- tetramethylbenzidine). Reaction was terminated with 2 M H
SO
and absorbance was measured at 450 nm. The signals generated were later used for normalization of ET-1 binding activity.
The time that I ve wasted is my biggest regretSpent in these places I will never forget.Just sitting and thinking about the things that I ve doneThe crying, the laughing, the hurt and the fun.
Analysis of ET- traps:ET-1 binding
To increase the adsorption surface area, and thus the detection signal, N-biotin-ET-1 (Phoenix pharma) was coupled to paramagnetic streptavidin beads (MyOne Streptavidin T1, Thermo Fisher Scientific; 10 mg/mL beads) as an alternative to the conventional phage ELISA on 96-well plates. 5 µL of beads were incubated with 2.5 × 10
ET-traps-displaying phages for 1 h in 500 µL of 0.5% milk/0.1% PBST. After washing and detection reaction, paramagnetic beads were captured on a magnet, and the supernatants were transferred to a 96-well plate for absorbance A
measurements. In parallel, binding of ET-trap constructs to streptavidin beads in absence of ET-1 was analyzed. Absorbance signals were blanksubtracted and normalized according to the relative display levels as determined in anti-cMyc phage ELISA assay.
Creating an Fc-fusion construct and measuring binding affinity to ET-1
The gene for the ET-traps was designed and optimized for expression in mammalian cells (HEK293) prior to being synthesised. The sequence was then sub-cloned into a cloning and expression vector for human Fc fusion proteins.
In brief, HEK293 cells were passaged to the optimum stage for transient transfection. Cells were transiently transfected with the appropriate expression vector and cultured for a further 6-14 d. An appropriate volume of cells was transfected with the aim of obtaining 1-5 mg of purified Fc fusion protein. Cultures were harvested and one-step purification performed using affinity chromatography. For this, culture supernatant containing Fc fusion protein was loaded onto a MabSelect SuRe Protein A column at 4 mL/min and washed with PBS pH 7.2. A step elution was performed with sodium citrate buffer (pH 3.0). Eluted protein was neutralised with 10% (v/v) Tris buffer (pH 9.0). Upon purification, the Fc fusion protein was buffer exchanged into PBS pH 7.4. The protein was analysed for purity by SDS-PAGE and concentration determined by UV spectroscopy (at 280 nm).
A new target in drug discovery and development.
RESULTS
ET-traps constructs were cloned and displayed in a monovalent phage display system using the 3 + 3 display approach (Figure 1). Each codon-optimized construct (Genscript) was amplified by polymerase chain reaction from its parent plasmid pUC18 using forward and reverse primers with overhangs harboring NcoI and NotI restriction sites. Reaction mixtures were subjected to agarose gel electrophoresis, amplicons were purified with QIAEX II gel extraction kit (Qiagen), and subsequently digested with NcoI/NotI restriction enzymes alongside pIT2 phagemid vector.
Phage display experiments
In this study, we first performed phage display experiments to ascertain the binding of the ET-traps to ET-1. Phage display is one of the most powerful and widely used laboratory techniques for the study of protein-protein, protein-peptide and protein-DNA interactions[29]. This technology is based on expressing the protein or peptide of interest on bacterial virus protein coat, allowing the study of molecular interaction between the virion-displayed ligand (in this case ET-traps) and an immobilized target (
ET-1).
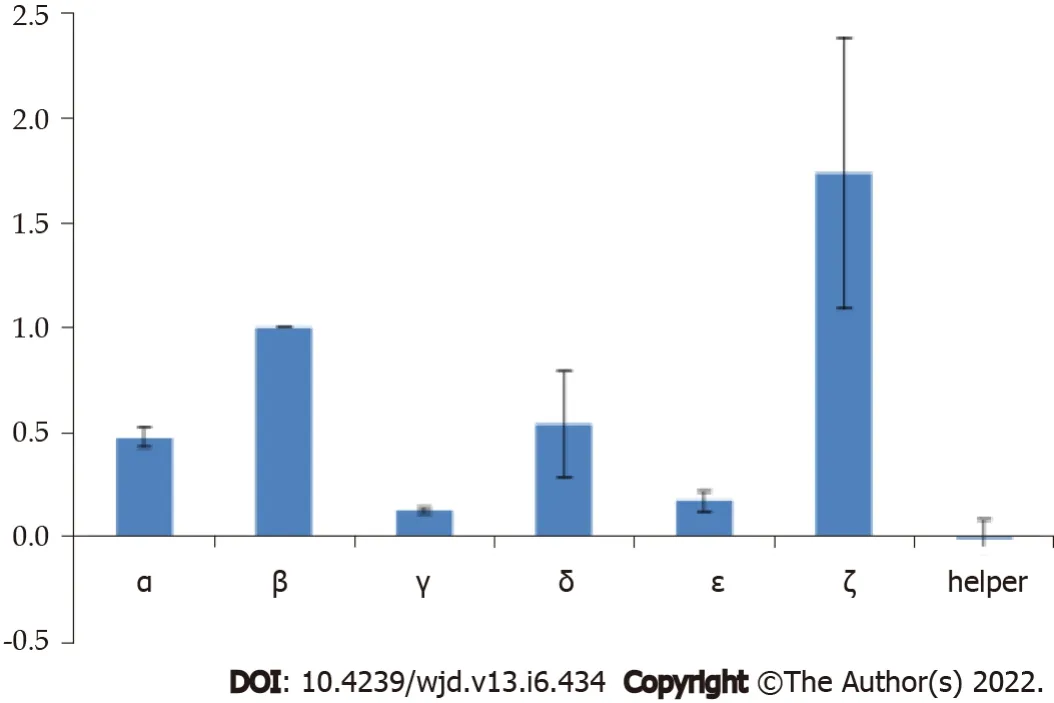
The construct ζ gave strong binding to ET-1 in phage display experiments, but the Fc-fusion molecule was not stable and the results could not be replicated with the soluble fusion protein. The phage experiments confirmed that construct β indeed gave consistent, high binding. It was further observed that in the form of an Fc-fusion construct β showed high binding affinity to ET-1 consistently (Figure 3).
DlSCUSSlON
This paper discusses the characterization of ET-traps that might be useful in the treatment of ET-1 related diseases or disorders, such as preeclampsia, cardiovascular diseases, chronic kidney disease, diabetes or neurodegenerative disorders[11,18,19,33-36]. Previous work has shown that the use of the selected ET-traps gave a therapeutic effect on reducing different markers of diabetes-induced disease pathology[12,13]. The ET-traps helped reduce different markers of diabetes disease pathology, such as over-expression of ECM proteins, proteinuria and tissue damage to kidneys and heart. This effect was found to be statistically significant both
and
. The ET-traps were designed based on a previous study[22]. The purpose of this study was to ascertain our selection of the ET-traps. Both ECL2 and ECL3, including the flanking transmembrane regions, were found to play an important role in ligand selection[22]. Further, residues in the intracellular loop and of the C-terminus are important for ET-1 binding[22]. These domains were used to create the final ET-traps that gave an efficacious, therapeutic effect in our proof-of-concept studies done both at the
and
levels in the diabetes disease space[12,13]. The final design of our ET-traps ensured that the selected ET-traps do not bind the ET-1 paralogs, which is important for selective activity and thus fewer potential adverse effects.
We began with phage display experiments to test different combinations of potential ET-traps, including the previously selected ET-traps that were used to perform the proof of concept (PoC) work[12,13]. These experiments confirmed the selection of the most efficient ET-traps. Phage experiments were performed on different combinations of sequences (Figure 3). These experiments allowed us to confirm our final ET-traps selection and we then proceeded to test the binding of its cognate Fc fusion to ET-1.
As ET-1 is abundant in the body while ET-2 is almost undetectable, ET-1 was more convenient to research; this assumption has meant ET-2 is relatively under-researched[26]. However, recent research evidence suggests distinct roles and features of ET-2. In mice with the ET-2 gene knocked-out, the animals displayed growth retardation, and were hypothermic and hypoglycemic, which resulted in early mortality[37].
Developed a potent tool targeting ET-1.
ET receptor antagonists (ERAs) can have a deleterious effect on physiological ET-2 functions by completely blocking the receptors and thereby inhibiting the physiological actions of ET-2. With our ETtraps, we would overcome this; the ET-traps specifically bind to ET-1 and do not block the receptors to effect ET-2 actions like an inhibitor to the ET system might do. The ET-traps have been designed to specifically bind ET-1.
Then the two brothers put on their armour, buckled10 their swords, and pocketed their purees; and, after taking a tender farewell of each other, they mounted their horses and went their various ways
Aberrations in the
gene that is responsible for producing ET-3 have been associated with congenital disorders involving neural crest-derived cells, like Hirschsprung disease and Waardenburg syndrome[38,39]. This shows that ET-3 is one of the important peptides of the ET family, which is involved in various developmental processes. Further, use of ERAs would essentially block the function of this molecule thereby potentially causing serious birth defects. This again precludes completely blocking the physiological functions of ET-3, as it is one of the important factors for essential developmental processes. Again, with the ET-traps, we would not completely be blocking the ET system, rather just targeting elevated ET-1 Levels upstream. As found in this study, the selected ET-traps bind ET-1 with a high binding affinity in the double-digit picomolar range (an average of 73.8 rM,
= 6). This was also previously found and reported by Jain
[12] in their diabetes PoC study. This work showed that the selected ET-traps have an efficacious, therapeutic effect in ameliorating diabetes disease pathology[12,13]. This was not associated with any toxic effects as evinced by the toxicology data. This corroborates that the selected ET-traps are efficacious at the working concentration and specific to just ET-1.
CONCLUSlON
The ET-traps were designed to specifically bind ET-1. The results of this study confirm that our selected ET-traps specifically bind to ET-1. This is in agreement with previous PoC studies that detected no toxic effects of the selected ET-traps at the working concentration. This is an important factor for the potential use of ET-traps as a therapeutic.
ARTlCLE HlGHLlGHTS
Research background
Endothelin (ET)-1 is a very potent vasoactive peptide that is significantly elevated in different diseases.
Research motivation
We wanted to develop a cure that would target this peptide and would help save millions of lives around the world.
Research objectives
To develop a tool that specifically targets ET-1.
Research methods
We employed phage display and binding assays.
Research results
A very high binding affinity was observed for our selected tool.
Research conclusions
The waiting-maid now mounted Falada, and the real bride the worse horse, and so they continued their journey till at length they arrived at the palace yard. There was great rejoicing over the arrival, and the Prince sprang forward to meet them, and taking the waiting-maid for his bride, he lifted her down from her horse and led her upstairs to the royal chamber13. In the meantime the real Princess was left standing14 below in the courtyard. The old King,25 who was looking out of his window, beheld15 her in this plight16, and it struck him how sweet and gentle, even beautiful, she looked. He went at once to the royal chamber, and asked the bride who it was she had brought with her and had left thus standing in the court below.
Research perspectives
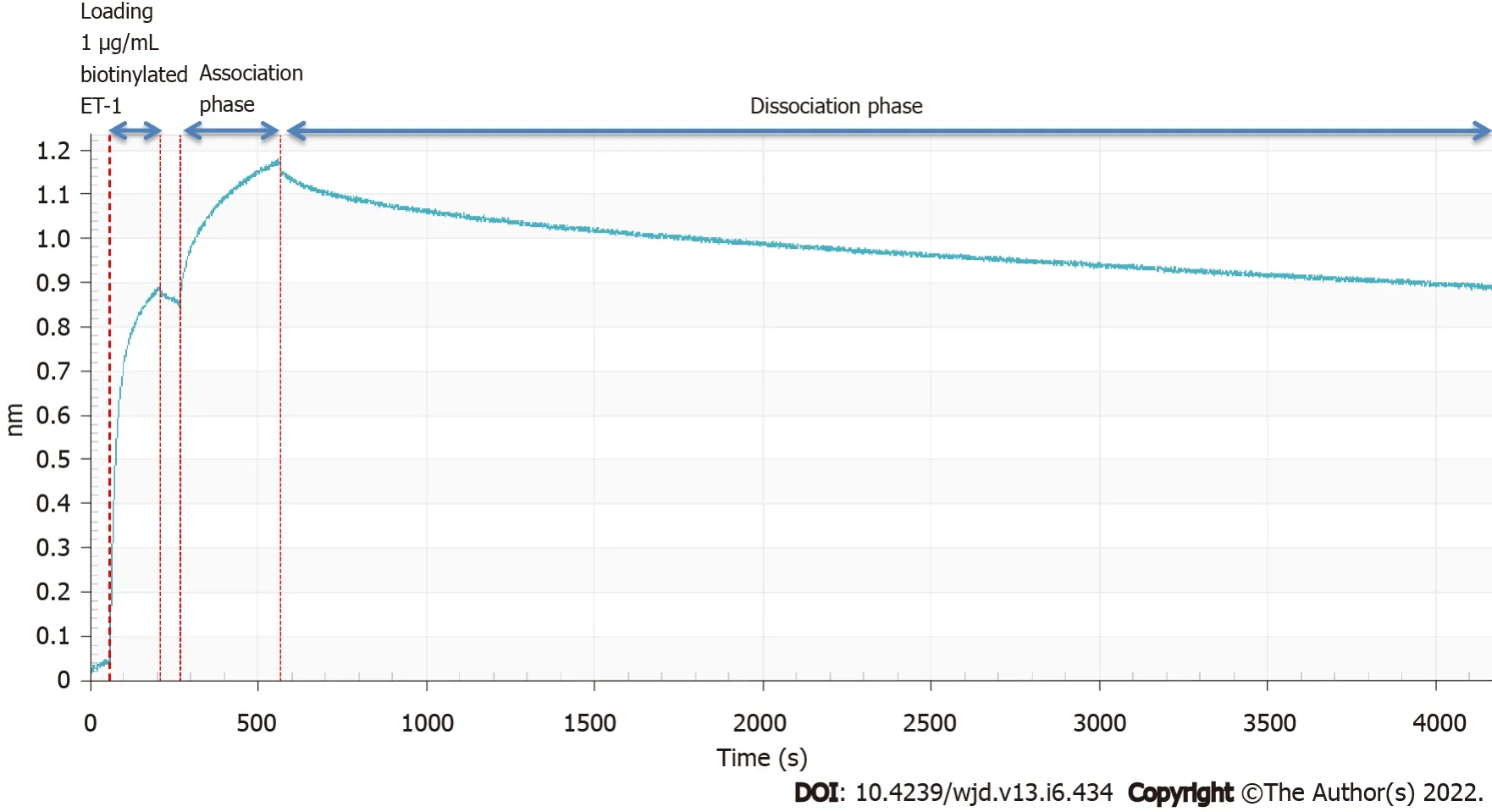
For the binding affinity measurement, we employed the use of the Octet Red96 system (Patel
2013). In brief, the kinetics of the selected ET-traps binding to biotinylated ET-1 (Phoenix Pharmaceuticals) was determined using the Octet Red96 system (ForteBio, Menlo Park, CA). The buffer for the assays was PBS with 0.01% (w/v) bovine serum albumin and 0.002% Tween20. The measurements were carried out at 30 °C. 1 µg/mL bio-ET-1 was captured on dip-and-read streptavidin sensors, followed by binding of the selected ET-traps at 500 nM concentration. The ForteBio Octet analysis software (ForteBio, Menlo Park, CA) was used to generate the sensorgram.
Then she stepped to the window and opened the lattice, saying, The room is so hot, but she looked after him as long as she could see the white plumes9 in his hat
This paper is dedicated to Dr. M.L. Mehrotra. We would also like to thank Mr. Ashok Jain for all his dedication and support during this difficult time.
And when the young man stood before the king, the king spoke6 to him and said, By your tricks and the pranks7 that you have played on other people, you have, in the eye of the law, forfeited8 your life
Jain A and Bozovičar K contributed equally to this work; Jain A, Bozovičar K, Mehrotra V, Bratkovič T performed the experimental analyses and contributed towards writing the article and Johnson M and Jha I revised it critically for important intellectual content.
This study was approved by ET-traps Limited.
The authors have no conflict of interest.
Tsar Dolmat hastened to the Palace and sent many soldiers in pursuit, but they could not catch the Gray Wolf, who soon overtook the Horse with the Golden Mane that bore Tsarevitch Ivan and the Tsarevna.
Data available from corresponding author, Dr. Arjun Jain at arjun@et-traps.co.uk.
For many moments, there is only silence. We cannot take our eyes from each other, and as the veils of time lift, we recognize the soul behind the eyes, the dear friend we once loved so much, whom we have never stopped loving, whom we have never stopped remembering.
This article is an open-access article that was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution NonCommercial (CC BYNC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is noncommercial. See:https://creativecommons.org/Licenses/by-nc/4.0/
Prince Milan loosened from his neck the little gold cross his mother had given him, and as soon as Hyacinthia grasped it, she had changed herself into a church, Milan into a monk45, and the horse into a belfry
And after this the real nightingale was banished from the empire, and the artificial bird placed on a silk cushion close to the emperor’s bed. The presents of gold and precious stones which had been received with it were round the bird, and it was now advanced to the title of “Little Imperial Toilet Singer,” and to the rank of No. 1 on the left hand; for the emperor considered the left side, on which the heart lies, as the most noble, and the heart of an emperor is in the same place as that of other people.
United Kingdom
Arjun Jain 0000-0003-0396-9290; Kristof Bozovicar 0000-0003-0025-1734; Vidhi Mehrotra 0000-0002-2037-4355; Tomaz Bratkovic 0000-0001-8367-5465; Martin H Johnson 0000-0002-5906-5883; Ira Jha 0000-0002-1067-8208.
Zhang H
A
Zhang H
1 Nelson J, Bagnato A, Battistini B, Nisen P. The endothelin axis:emerging role in cancer.
2003; 3:110-116 [PMID:12563310 DOI:10.1038/nrc990]
2 Böhm F, Pernow J. The importance of endothelin-1 for vascular dysfunction in cardiovascular disease.
2007; 76:8-18 [PMID:17617392 DOI:10.1016/j.cardiores.2007.06.004]
3 Wernly B, Jung C. Trapping endothelin-1 to hunt down cardiovascular disease?
2019; 24:2108-2110[PMID:31654816 DOI:10.1016/j.drudis.2019.10.007]
4 Ergul A. Endothelin-1 and endothelin receptor antagonists as potential cardiovascular therapeutic agents.
2002; 22:54-65 [PMID:11794430 DOI:10.1592/phco.22.1.54.33505]
5 Ranno E, D'Antoni S, Spatuzza M, Berretta A, Laureanti F, Bonaccorso CM, Pellitteri R, Longone P, Spalloni A, Iyer AM,Aronica E, Catania MV. Endothelin-1 is over-expressed in amyotrophic lateral sclerosis and induces motor neuron cell death.
2014; 65:160-171 [PMID:24423643 DOI:10.1016/j.nbd.2014.01.002]
6 Ferrari CC, Tarelli R. Parkinson's disease and systemic inflammation.
2011; 2011:436813 [PMID:21403862 DOI:10.4061/2011/436813]
7 Jain A. Endothelin-1-induced endoplasmic reticulum stress in disease.
2013; 346:163-172 [PMID:23740603 DOI:10.1124/jpet.113.205567]
8 Jain A, Coffey C, Mehrotra V, Flammer J. Endothelin-1 traps as a potential therapeutic tool:from diabetes to beyond?
2019; 24:1937-1942 [PMID:31394173 DOI:10.1016/j.drudis.2019.07.008]
9 Nelson JB, Hedican SP, George DJ, Reddi AH, Piantadosi S, Eisenberger MA, Simons JW. Identification of endothelin-1 in the pathophysiology of metastatic adenocarcinoma of the prostate.
1995; 1:944-949 [PMID:7585222 DOI:10.1038/nm0995-944]
10 Chiao JW, Moonga BS, Yang YM, Kancherla R, Mittelman A, Wu-Wong JR, Ahmed T. Endothelin-1 from prostate cancer cells is enhanced by bone contact which blocks osteoclastic bone resorption.
2000; 83:360-365[PMID:10917552 DOI:10.1054/bjoc.2000.1261]
11 Jain A. Endothelin-1:a key pathological factor in pre-eclampsia?
2012; 25:443-449 [PMID:22995748 DOI:10.1016/j.rbmo.2012.07.014]
12 Jain A, Chen S, Yong H, Chakrabarti S. Endothelin-1 traps potently reduce pathologic markers back to basal levels in an in vitro model of diabetes.
2018; 17:189-195 [PMID:30918854 DOI:10.1007/s40200-018-0360-8]
13 Jain A, Mehrotra V, Jha I, Jain A. In vivo studies demonstrate that endothelin-1 traps are a potential therapy for type I diabetes.
2019; 18:133-143 [PMID:31275884 DOI:10.1007/s40200-019-00400-7]
14 Emerging Risk Factors Collaboration, Sarwar N, Gao P, Seshasai SR, Gobin R, Kaptoge S, Di Angelantonio E,Ingelsson E, Lawlor DA, Selvin E, Stampfer M, Stehouwer CD, Lewington S, Pennells L, Thompson A, Sattar N, White IR, Ray KK, Danesh J. Diabetes mellitus, fasting blood glucose concentration, and risk of vascular disease:a collaborative meta-analysis of 102 prospective studies.
2010; 375:2215-2222 [PMID:20609967 DOI:10.1016/S0140-6736(10)60484-9]
15 Chen S, Mukherjee S, Chakraborty C, Chakrabarti S. High glucose-induced, endothelin-dependent fibronectin synthesis is mediated
NF-kappa B and AP-1.
2003; 284:C263-C272 [PMID:12388107 DOI:10.1152/ajpcell.00192.2002]
16 Chen S, Feng B, Thomas AA, Chakrabarti S. miR-146a regulates glucose induced upregulation of inflammatory cytokines extracellular matrix proteins in the retina and kidney in diabetes.
2017; 12:e0173918 [PMID:28301595 DOI:10.1371/journal.pone.0173918]
17 Law B, Fowlkes V, Goldsmith JG, Carver W, Goldsmith EC. Diabetes-induced alterations in the extracellular matrix and their impact on myocardial function.
2012; 18:22-34 [PMID:22221857 DOI:10.1017/S1431927611012256]
18 Seligman BG, Biolo A, Polanczyk CA, Gross JL, Clausell N. Increased plasma levels of endothelin 1 and von Willebrand factor in patients with type 2 diabetes and dyslipidemia.
2000; 23:1395-1400 [PMID:10977040 DOI:10.2337/diacare.23.9.1395]
19 Schneider JG, Tilly N, Hierl T, Sommer U, Hamann A, Dugi K, Leidig-Bruckner G, Kasperk C. Elevated plasma endothelin-1 levels in diabetes mellitus.
2002; 15:967-972 [PMID:12441216 DOI:10.1016/s0895-7061(02)03060-1]
20 Karet FE, Davenport AP. Endothelin and the human kidney:a potential target for new drugs.
1994; 9:465-468 [PMID:8090323 DOI:10.1093/ndt/9.5.465]
21 Orry AJ, Wallace BA. Modeling and docking the endothelin G-protein-coupled receptor.
2000; 79:3083-3094[PMID:11106614 DOI:10.1016/S0006-3495(00)76543-X]
22 Adachi M, Hashido K, Trzeciak A, Watanabe T, Furuichi Y, Miyamoto C. Functional domains of human endothelin receptor.
1993; 22 Suppl 8:S121-S124 [PMID:7509921 DOI:10.1097/00005344-199322008-00033]
23 Davenport AP, Hyndman KA, Dhaun N, Southan C, Kohan DE, Pollock JS, Pollock DM, Webb DJ, Maguire JJ.Endothelin.
2016; 68:357-418 [PMID:26956245 DOI:10.1124/pr.115.011833]
24 Yanagisawa M, Kurihara H, Kimura S, Tomobe Y, Kobayashi M, Mitsui Y, Yazaki Y, Goto K, Masaki T. A novel potent vasoconstrictor peptide produced by vascular endothelial cells.
1988; 332:411-415 [PMID:2451132 DOI:10.1038/332411a0]
25 Inoue A, Yanagisawa M, Kimura S, Kasuya Y, Miyauchi T, Goto K, Masaki T. The human endothelin family:three structurally and pharmacologically distinct isopeptides predicted by three separate genes.
1989;86:2863-2867 [PMID:2649896 DOI:10.1073/pnas.86.8.2863]
26 Ling L, Maguire JJ, Davenport AP. Endothelin-2, the forgotten isoform:emerging role in the cardiovascular system,ovarian development, immunology and cancer.
2013; 168:283-295 [PMID:22118774 DOI:10.1111/j.1476-5381.2011.01786.x]
27 Hofstra RM, Osinga J, Tan-Sindhunata G, Wu Y, Kamsteeg EJ, Stulp RP, van Ravenswaaij-Arts C, Majoor-Krakauer D,Angrist M, Chakravarti A, Meijers C, Buys CH. A homozygous mutation in the endothelin-3 gene associated with a combined Waardenburg type 2 and Hirschsprung phenotype (Shah-Waardenburg syndrome).
1996; 12:445-447[PMID:8630503 DOI:10.1038/ng0496-445]
28 Baynash AG, Hosoda K, Giaid A, Richardson JA, Emoto N, Hammer RE, Yanagisawa M. Interaction of endothelin-3 with endothelin-B receptor is essential for development of epidermal melanocytes and enteric neurons.
1994; 79:1277-1285[PMID:8001160 DOI:10.1016/0092-8674(94)90018-3]
29 Wu CH, Liu IJ, Lu RM, Wu HC. Advancement and applications of peptide phage display technology in biomedical science.
2016; 23:8 [PMID:26786672 DOI:10.1186/s12929-016-0223-x]
30 Day LA, Wiseman L. A Comparison of DNA Packaging in the Virions of fd, Xf, and Pf1. Cold Spring Harbor:New York, 1978:605-625 [DOI:10.1016/0022-2836(77)90088-2]
31 Jain A, Mehrotra V, Yong H, Hiremath K, Jain A, Johnson M, Jha I. Creating a Soluble Binder to Endothelin-1 based on the natural ligand binding domains of the endothelin-1 (G-protein-coupled) receptor.
2019; 25:107-114 [DOI:10.1007/s10989-017-9653-x]
32 Smith GP, Petrenko VA. Phage Display.
1997; 97:391-410 [PMID:11848876 DOI:10.1021/cr960065d]
33 Zeiher AM, Goebel H, Schächinger V, Ihling C. Tissue endothelin-1 immunoreactivity in the active coronary atherosclerotic plaque. A clue to the mechanism of increased vasoreactivity of the culprit lesion in unstable angina.
1995; 91:941-947 [PMID:7850978 DOI:10.1161/01.cir.91.4.941]
34 Haufschild T, Shaw SG, Kesselring J, Flammer J. Increased endothelin-1 plasma levels in patients with multiple sclerosis.
2001; 21:37-38 [PMID:11315981 DOI:10.1097/00041327-200103000-00011]
35 Pache M, Kaiser HJ, Akhalbedashvili N, Lienert C, Dubler B, Kappos L, Flammer J. Extraocular blood flow and endothelin-1 plasma levels in patients with multiple sclerosis.
2003; 49:164-168 [PMID:12646761 DOI:10.1159/000069085]
36 Sethi AS, Lees DM, Douthwaite JA, Dawnay AB, Corder R. Homocysteine-induced endothelin-1 release is dependent on hyperglycaemia and reactive oxygen species production in bovine aortic endothelial cells.
2006; 43:175-183[PMID:16410680 DOI:10.1159/000090947]
37 Chang I, Bramall AN, Baynash AG, Rattner A, Rakheja D, Post M, Joza S, McKerlie C, Stewart DJ, McInnes RR,Yanagisawa M. Endothelin-2 deficiency causes growth retardation, hypothermia, and emphysema in mice.
2013; 123:2643-2653 [PMID:23676500 DOI:10.1172/JCI66735]
38 Liu Q, Cheng J, Lu Y, Zhou J, Wang L, Yang C, Yang G, Yang H, Cao J, Zhang Z, Sun Y. The clinical and genetic research of Waardenburg syndrome type I and II in Chinese families.
2020; 130:109806[PMID:31812001 DOI:10.1016/j.ijporl.2019.109806]
39 Sánchez-Mejías A, Fernández RM, López-Alonso M, Antiñolo G, Borrego S. New roles of EDNRB and EDN3 in the pathogenesis of Hirschsprung disease.
2010; 12:39-43 [PMID:20009762 DOI:10.1097/GIM.0b013e3181c371b0]
