Na+与Cl-浓度平衡供应促进小白菜干物质积累及营养品质提升
2022-06-17张维霞王俊玲陈倩倩薛占军高志奎
张维霞,高 爽,王俊玲,陈倩倩,薛占军,高志奎*
(1 河北农业大学园艺学院,河北保定 071001;2 河北农业大学食品科技学院,河北保定 071001;3 河北农业大学生命科学学院,河北保定 071001)
低浓度的 Na+和Cl-有益于植物的生长[1-4]。喷施低浓度NaCl也促进了黄瓜幼苗的生长[5]。虽然NaCl的研究多年来主要集中在土壤或根际高浓度NaCl(50~300 mmol/L)带来的盐胁迫效应[6-9]以及作物的耐盐机制[10-11]上,但是近年来低浓度(1~5 mmol/L)NaCl对植物生长的有益效应[4-5,12-14]也逐渐受到关注。
Franco-Navarro等[4]在根际营养液中的K+、Ca2+、Mg2+阳离子保持一致的条件下,发现Cl-在1~5 mmol/L低浓度范围内,可提高作物的干物质累积和含水量,增加叶面积和叶片厚度,提高叶片光合速率(Pn)。王宝增等[12]在Hoagland营养液中添加2.5~5.0 mmol/L NaCl促进了小麦植株干物质累积和光合速率。孟闯等[5]在叶面喷施5~10 mmol/L NaCl提高了黄瓜幼苗干物质量、叶面积、含水量、Pn、气孔导度(Gs)以及植株的可溶性蛋白质、可溶性糖和游离氨基酸含量。在水培番茄[13]、水培韭菜[2,14]、基质培苋菜[3]上也有低浓度NaCl的促生效应。然而,较少研究区分Cl-和Na+的效应,以及Na+与Cl-之间的相互关系。在Na+与Cl-共同存在时,NaCl盐胁迫效应的研究中,Srivastava[15]认为NaCl对柑橘的盐胁迫主要由于Cl-(30 mmol/L)引起,而许兴等[16]认为小麦的盐胁迫主要由Na+(340 mmol/L)所致;鲁少尉等[17]则认为Na+(50 mmol/L)对番茄叶片中糖代谢的影响显著大于Cl-(50 mmol/L)。低浓度时,Na+与Cl-离子的摩尔质量相等,导致难以辨析Na+和Cl-的效应。因此,本试验在[Na+]=[Cl-]、[Na+]<[Cl-]和[Na+]>[Cl-] 3种离子浓度条件下,观察了小白菜的生长以及叶片光合效率等指标,探索Na+和Cl-的单独效应、复合效应以及交互效应。
1 材料与方法
1.1 试验材料
供试验材料为小白菜(Brassica campestrisL. ssp.chinensisvar.communisTsen et Lee )‘寒月F1’,于2021年3月1日在河北农业大学东校区日光温室内播种育苗,穴盘为50孔,每穴3~5粒种子,苗期间苗2次,留长势整体一致、生长健壮的秧苗。基质为硕丰园艺(N+P2O5+K2O≥2%,有机质≥40%)有机通用基质。在小白菜2叶1心时挑选长势一致的幼苗移栽到营养钵(直径13 cm,高12 cm)中;当小白菜生长至3叶1心时,每日上午8:00~8:30进行1次叶面喷施。以叶面均匀铺满溶液且叶缘开始有液珠滴落为宜。喷施于3月26日开始,到4月15日结束,同时以叶面喷施无离子水为对照CK,连续叶施处理21天后取样测定相关指标。
1.2 试验设计
以Hoagland营养液为基础配制处理溶液,基础组成为:K+2.121 mmol、Ca2+2 mmol、Mg2+1 mmol、NO3-5.25 mmol、PO43-0.798 mmol、SO42-1.13 mmol。Franco-Navarro 等[4]为了辨析 Cl-的有益效应,在基础营养液中添加了3种氯化物(KCl 2.5 mmol、MgCl20.625 mmol、CaCl20.625 mmol)来凸显Cl-阴离子的效应,并弱化伴随阳离子的营养效应。鲁少尉等[17]在辨析Na+和Cl-的单盐胁迫效应方案中,采用3种含钠化合物进行了单Na+盐处理(NaNO3∶NaHCO3∶NaH2PO4=5∶4∶1),以及 2 种氯化物进行了单Cl-盐处理(KCl∶NH4Cl=1∶1)。本试验为了辨析Na+或Cl-促进小白菜生长效应的差异性,在氯化钠中添加3种氯化物或3种含钠化合物进行了Na+与Cl-平衡及非平衡配制。试验共设了3 个 [Na+]=[Cl-]的离子平衡处理 (B1、B2、B3),3 个[Na+]<[Cl-]处理 (A1、A2、A3),即阴离子 Cl-占优势;3个[Na+]>[Cl-]处理(C1、C2、C3),即阳离子Na+占优势,具体处理的离子浓度见表1,试验采用随机区组设计,共设5次重复,每个重复20株。
值得注意的是,在A组处理的[Na+]<[Cl-]中,为了凸显Cl-的效应,采用3种伴随阳离子K+、Ca2+、Mg2+,并保持较低浓度水平,分别为Hoagland营养液中的50%、25%、25%,来弱化伴随阳离子对小白菜的营养效应[4,17]。相应地,在C组处理的[Na+]>[Cl-]中,为了凸显Na+的效应,采用3种伴随阴离子NO3-、PO43-、SO42-,并保持较低浓度水平,分别为Hoagland营养液中的29%、33%、25%,来弱化伴随阴离子对小白菜的营养效应[17]。无论如何,3种相伴随的阳离子或阴离子仍然会有微弱的营养效应。
1.3 测定项目与方法
1.3.1 生长及形态指标的测定 小白菜株高采用直尺测量;茎粗采用数显游标卡尺测定;叶面积采用LI-3000叶面积仪测量。将小白菜的叶片、叶柄、茎、根分开,用电子天平分别测定鲜重后,置于105℃的烘箱中杀青0.5 h,继后在80℃烘干至恒重并称量其干重。每个处理3次重复,每个重复随机测定5株。
1.3.2 营养物质含量的测定 小白菜可溶性糖、淀粉的测定采用硫酸蒽酮法[18],游离氨基酸总量的测定采用水合茚三酮法[18],可溶性蛋白的测定采用考马斯亮蓝G-520法[19],3次重复测定。
1.3.3 叶片组织结构的测定 选择每株小白菜自下而上第5片叶片的同一部位,用透明胶带-指甲油印迹法[20]制成叶片气孔切片,测量小白菜气孔长度和气孔开度及数量,计算叶片下表皮气孔密度;用刀片横切叶片制成切片,测量叶片、栅栏组织、海绵组织、表皮细胞的厚度,3次重复测定。
1.3.4 光合碳吸收的测定 小白菜采用CIRAS-2光合仪测定自下而上第5片叶的光合气体交换参数。测定时光源为红蓝3∶1复合光源,光照强度为1500 μmol/(m2·s),叶室温度为 20℃,CO2浓度为380 mg/L,3 次重复测定。
1.4 数据处理与分析
用软件Excel 2010将试验数据录入、处理并作图,用SPSS 26.0软件对数据进行分析并进行2因素主体间效应检验,按照Duncan’s法P<0.05为差异显著进行多重比较。
2 结果与分析
2.1 喷施不同Na+和Cl-平衡状态溶液对小白菜生长及形态的影响
无论喷施溶液的[Na+]和[Cl-]是否相等,与清水对照相比,均显著提高小白菜的株高、茎粗、叶面积、叶片肉质性、叶片持水性和植株鲜重(图1和表2)。在[Na+]=[Cl-]的B组中,植株鲜重与叶面积增加最明显,平均较CK分别增加了38.4%和30.3%,B3的比叶重增加了12.1%;在 [Na+]<[Cl-]的A组中,A2处理的株高、茎粗、叶片持水性和植株鲜重增加最大,分别较CK增加了27.7%、35.1%、21.1%和38.7%;在[Na+]>[Cl-]的C组中,C1处理的比叶重与CK相比显著增加了8.6%,C2和C3处理的叶片肉质性分别比CK增加了30.4%和29.4%,C3处理的茎粗比CK显著增加了23.9%。

图1 不同Na+和Cl-平衡状态处理下小白菜植株形态Fig. 1 Morphology of pakchoi under equal and unequal Na+ and Cl- concentration
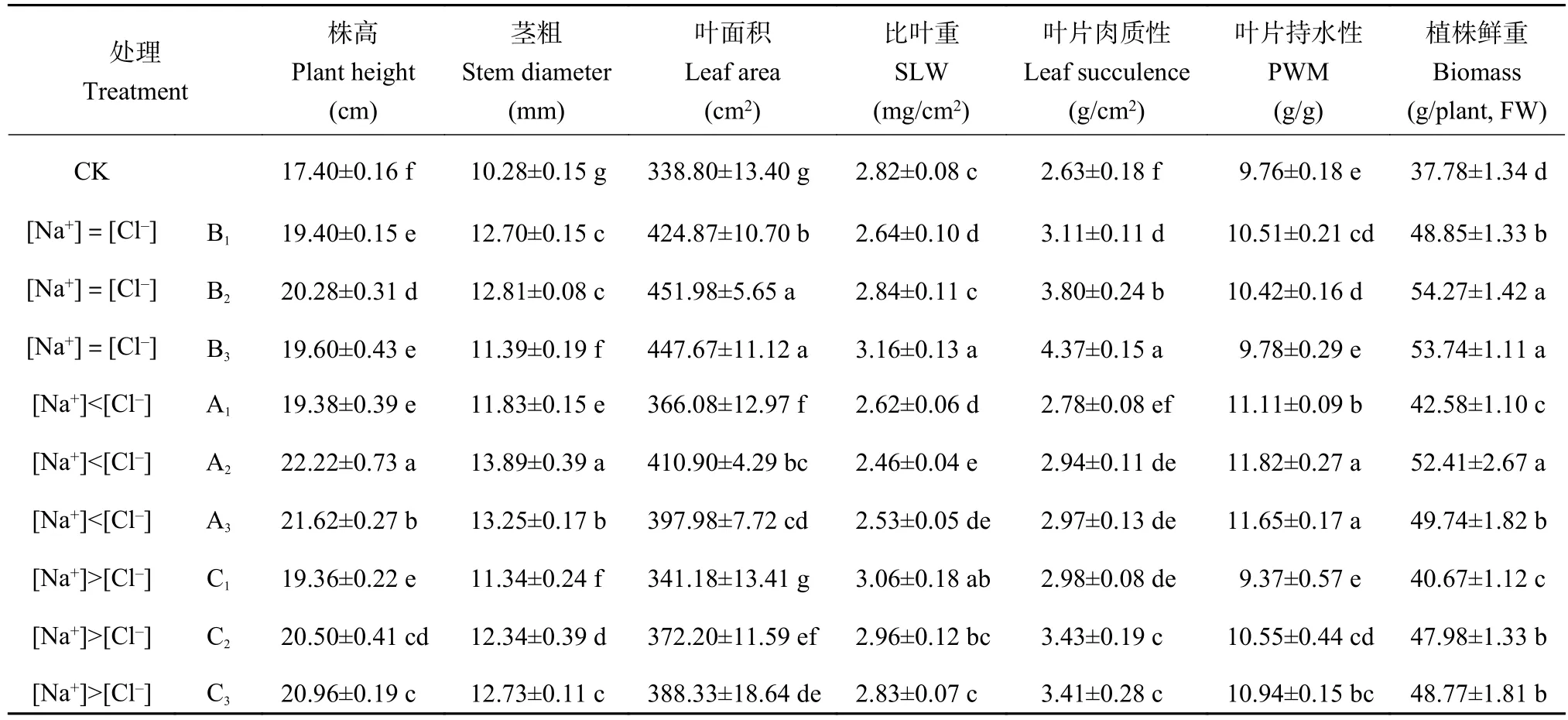
表2 Na+和Cl-平衡与非平衡处理下小白菜的生长及形态指标Table 2 Index of growth and morphology of pakchoi under equal and unequal Na+ and Cl- concentration
因此,当[Na+]=[Cl-]时对小白菜的生长及形态尤其是植株鲜重和叶面积的促进效应较大;当[Na+]<[Cl-]时对小白菜的株高、茎粗、叶片持水性和植株鲜重的促进效应较大;当[Na+]>[Cl-]时对小白菜的比叶重、叶片肉质性、茎粗促进效应较大。
2.2 不同Na+和Cl-平衡态处理对小白菜植株干物质累积与含水量的影响
与CK 相比,B1与C1处理的植株干物质分别增加了21.5%与11.0%,含水量显著增加了30.3% 和7.3%,而A1对植株干物质无显著影响,但A1植株含水量显著增加了14.3% (图2)。另外,B2处理的植株干物质和含水量比CK分别增加了35.1%和44.4%,且B组处理的植株干物质和含水量增加最显著,平均比CK分别增加了33.5%和38.9%。
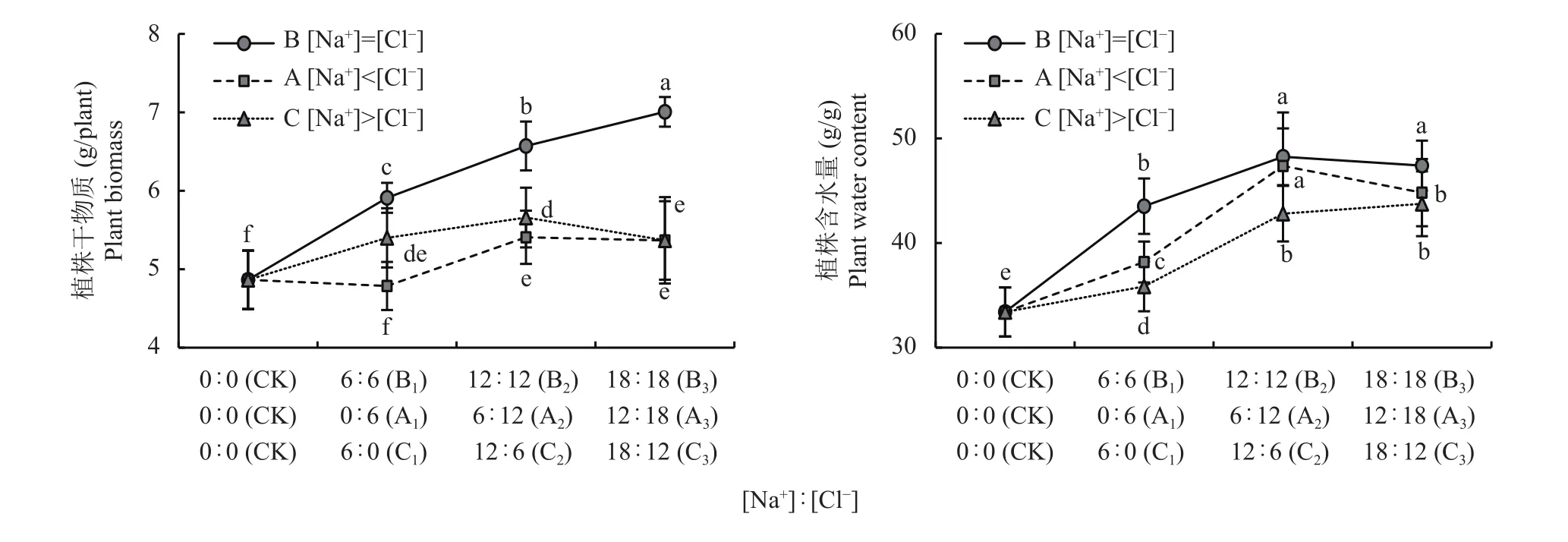
图2 相同和不同Na+和Cl-喷施浓度下小白菜植株干物质重和含水量Fig. 2 Biomass and water content in pakchoi plant under equal and unequal Na+ and Cl- concentration
各组内3个处理的植株干物质与含水量表现为,与B1处理相比,B2和B3处理平均增加了18.1%与12.9%;与A1处理相比,A2和A3处理分别平均增加了12.3%与23.7%;与C1处理相比,C2和C3分别平均增加了2.4%与22.3%。植株干物质量增幅以B组最大,其次是A组,最低为C组,而含水量增幅也以B组最大、A组次之,再次是C组。
这表明,[Na+]=[Cl-]平衡时对干物质累积和水分累积的促进效应最大;而在其他阳离子(K+、Ca2+、Mg2+)或阴离子(NO3-、PO43-、SO42-)伴随下,无Cl-时Na+有利于干物质累积,有Cl-时Cl-比Na+更有利于干物质累积;无Na+时Cl-有利于水分累积,有Na+时Na+和Cl-均有利于水分累积。
2.3 不同 Na+和Cl-的平衡态处理对小白菜植株营养物质含量的影响
从图3可见,与CK相比,植株可溶性糖和植株淀粉含量随Na+和Cl-的增加均呈降低趋势,但其中A1和C1的可溶性糖有显著增加,分别增加7.2%和12.3%;同时,植株可溶性蛋白质随Na+和Cl-的增加呈轻微降低或保持不变的趋势。
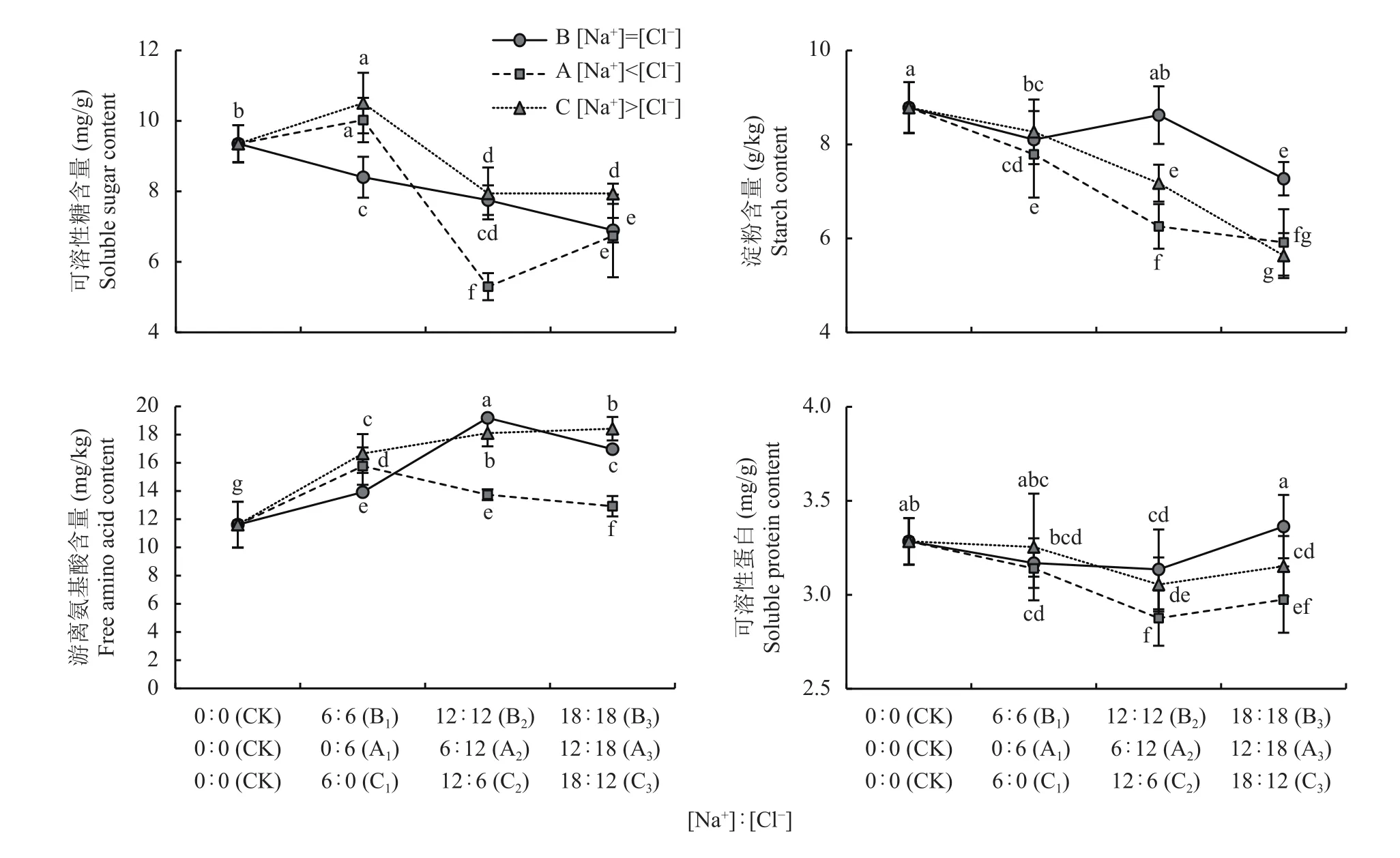
图3 相同和不同Na+和Cl-喷施浓度下小白菜植株可溶性糖、游离氨基酸、可溶性蛋白质和淀粉含量Fig. 3 Contents of soluble sugar, free amino acid, soluble protein and starch in pakchoi plant under equal and unequal Na+ and Cl- concentration
无论是 B组 (即 [Na+]=[Cl-])还是 A组 (即[Na+]<[Cl-])和C组(即[Na+]>[Cl-])处理,植株游离氨基酸含量与CK相比均有显著增加。其中,[Na+]=[Cl-]和[Na+]<[Cl-]处理的植株游离氨基酸含量随Na+和Cl-的增加呈现峰值变化趋势,B1、B2、B3处理的植株游离氨基酸含量分别比CK增加了19.8%、65.2%、46.1%,A1处理的植株游离氨基酸含量在A组中增加最多,比CK增加了35.7%。[Na+]>[Cl-]处理的C组植株游离氨基酸含量随Na+和Cl-的增加呈现趋饱和变化趋势,平均比CK增加了52.6%。
因此,Na+和Cl-对游离氨基酸含量的累积均有促进作用,Na+的促进效应明显大于Cl-的促进效应,且Na+与Cl-平衡时(B2)的促进效应最大。
2.4 不同Na+和Cl-平衡态处理对小白菜不同叶位单叶面积和单叶干物质的影响
在小白菜长到3叶1心时进行叶面喷施处理后表明,与CK相比,B1、B2、B3处理(即[Na+]=[Cl-])的下位叶(1~3片真叶)单叶面积和单叶干物质含量均有轻微的变化(图4,[Na+]=[Cl-]);中位叶(4~6片真叶)、上位叶(7~9片真叶)的单叶面积和单叶干物质含量均有显著增加,分别增加26.1%、55.5%和19.0%、64.2%。
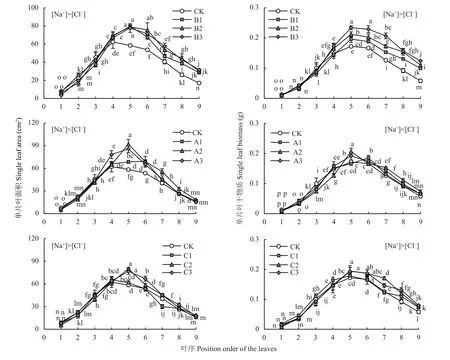
图4 相同和不同Na+和Cl-喷施浓度下小白菜不同叶位叶片面积和单片叶干物质量Fig. 4 Leaf area and biomass of pakchoi at different positions under equal and unequal Na+ and Cl- concentration
从图4中[Na+]<[Cl-]组可见,与CK相比,A1、A2、A3处理(即[Na+]<[Cl-])的下位叶的单叶面积和单叶干物质含量均有轻微变化;中位叶的单叶面积显著增加26.3%,而中位叶的单叶干物质含量均有轻微地增加,增加4.4%;上位叶的单叶面积和单叶干物质含量均有轻微地增加,分别增加9.4%和13.5%。
从图4中[Na+]>[Cl-]组可见,与CK相比,C1、C2、C3处理([即Na+]>[Cl-])仅中位叶的单叶面积显著增加12.8%,而中位叶和上位叶的单叶干物质含量均有显著增加,分别增加6.8%、23.5%。
因此,当[Na+]=[Cl-]时主要促进了中上位单叶面积的增加和单叶干物质的累积;当[Na+]<[Cl-]时主要促进了中位单叶面积的增加;当[Na+]>[Cl-]时主要促进了中位单叶面积的增加和中上位单叶干物质的累积。
2.5 不同Na+和Cl-平衡态处理对小白菜叶片组织结构的影响
从图5可见,无论是B组[Na+]=[Cl-]处理还是A组[Na+]<[Cl-]处理和C组[Na+]>[Cl-]处理,与CK相比,不仅增加了小白菜的叶片厚度,还主要增加了表皮细胞厚度和栅栏组织厚度(A1、C1除外)以及栅海比(栅栏组织与海绵组织厚度的比值,A1除外);也显著增加了气孔长度和气孔开度(A1除外)。另外,A1处理的海绵组织厚度增加最大;仅有B2和C1、C2处理气孔密度高于对照CK。
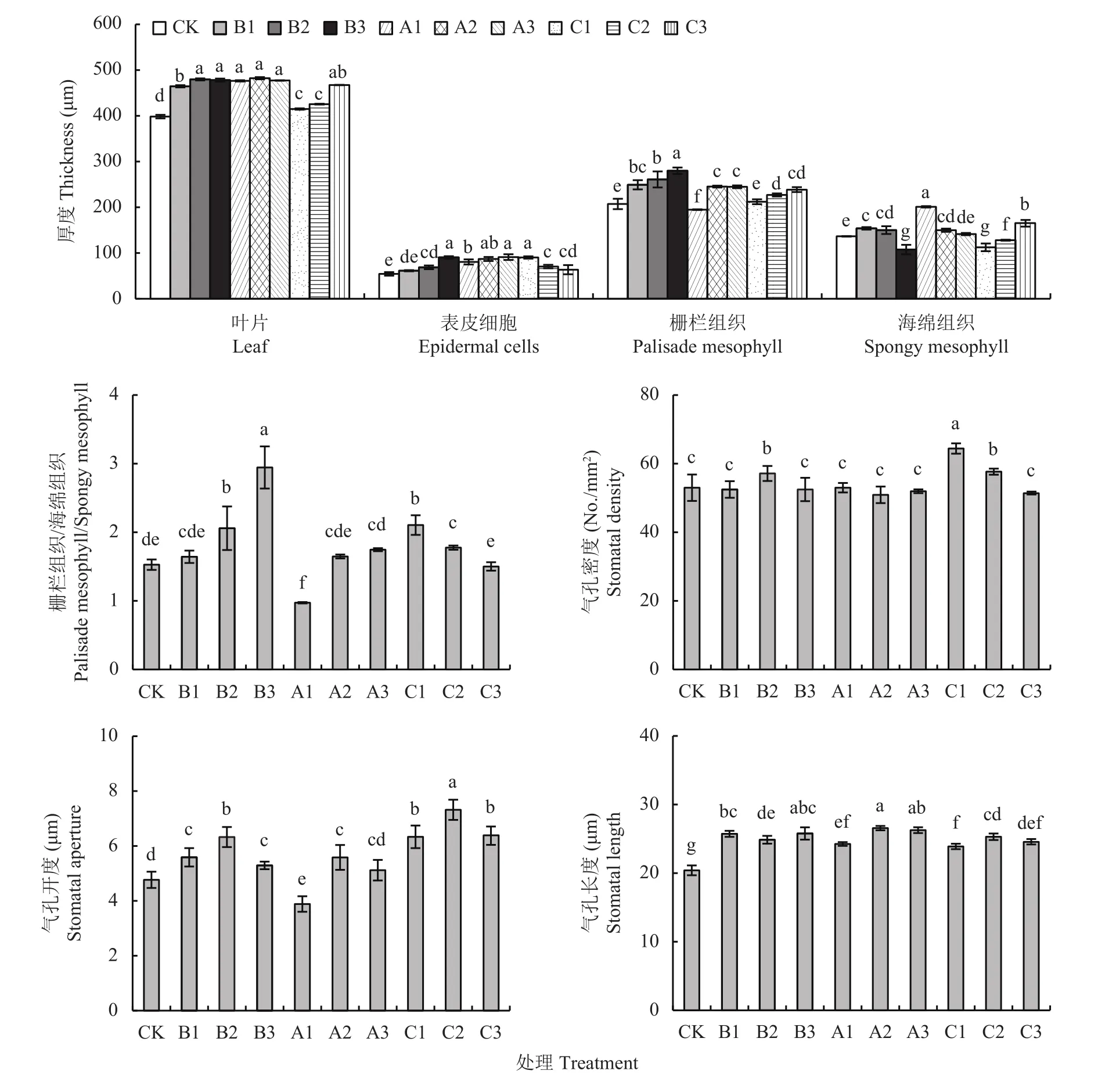
图5 相同和不同Na+和Cl-喷施浓度对小白菜叶片组织结构的影响Fig. 5 Leaf tissue structure of pakchoi under equal and unequal Na+ and Cl- concentration
综合来看,Na+尤其是Cl-促进了小白菜栅栏组织和海绵组织厚度以及气孔长度,而Na+更有利于促进气孔开度和气孔密度;[Na+]=[Cl-]组对栅栏组织、栅海比的促进效应最大。
2.6 不同Na+和Cl-平衡态处理对小白菜叶片光合气体交换活性的影响
小白菜进行叶面喷施处理后,B1、A1和C1处理的Pn和Gs与CK相比均显著增加,其中[Na+]<[Cl-]组中A1处理增加最大,分别增加了19.7%和140.9%。
在B1、A1、C1处理的基础上继续添加NaCl后(图6),B 组 [Na+]=[Cl-]处理的Pn和Gs随着 Na+和Cl-的增加呈现逐渐增加趋势,其中B3处理的Pn和Gs比B1处理分别增加了11.8%和78.8%;A组[Na+]<[Cl-]处理Pn随着Na+和Cl-的增加呈现峰值变化趋势,平均比 CK 增加了 17.3%,而Gs 则呈现趋饱和变化趋势,平均比 CK 增加了 145.3%;C组[Na+]>[Cl-]处理中随着Na+和Cl-的增加Pn无明显变化,但Gs呈下降趋势。
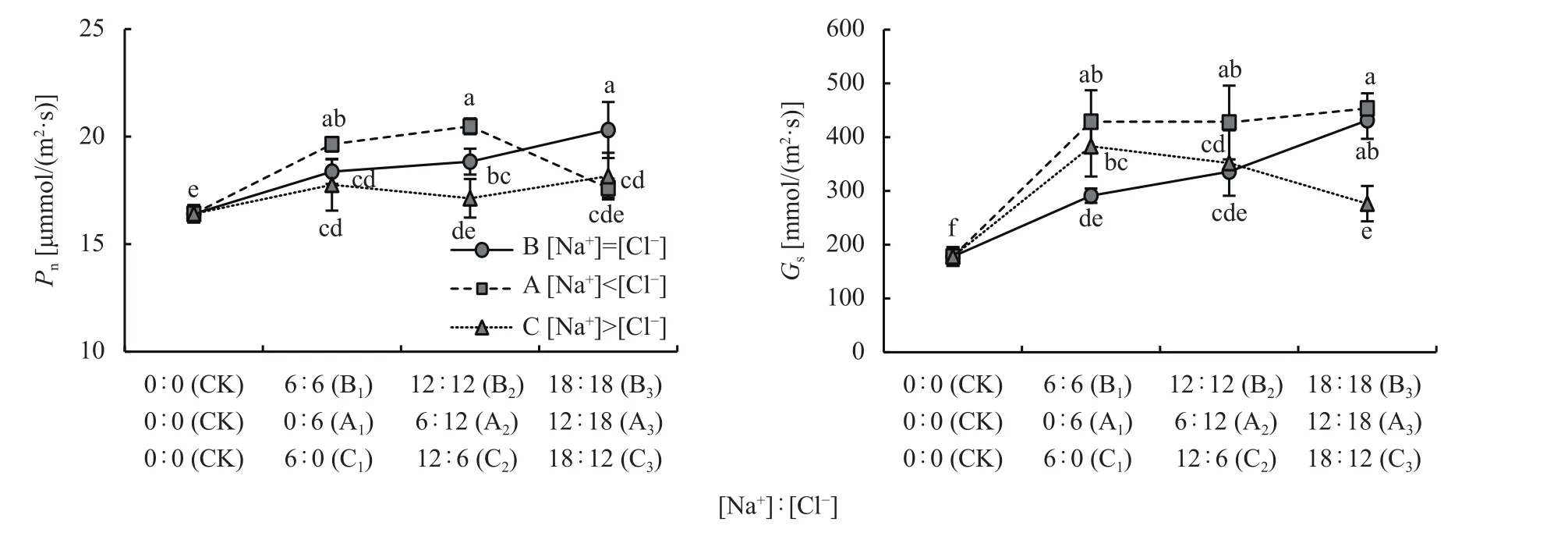
图6 相同和不同Na+和Cl-喷施浓度对小白菜叶片光合气体交换活性的影响Fig. 6 Photosynthetic gas exchange activity of pakchoi under equal and unequal Na+ and Cl- concentration
这表明,[Na+]=[Cl-]平衡及[Na+]<[Cl-]非平衡时对小白菜叶片光合气体交换活性的促进效应较大;而[Na+]>[Cl-]非平衡时促进效应较小。
2.7 主体间效应检验
由两因素主体间效应检验分析可知,小白菜的株高、茎粗、叶面积、比叶重、叶片肉质性、叶片持水性、全株鲜重、植株干物质累积、水分累积、营养物质含量、中上叶位单叶面积、单叶干物质、叶片组织结构和叶片光合气体交换活性,既受Na+(单叶6和4除外)、Cl-(叶片肉质性除外)单独离子效应的影响,又受Na+与Cl-交互效应的影响(表3、表4)。
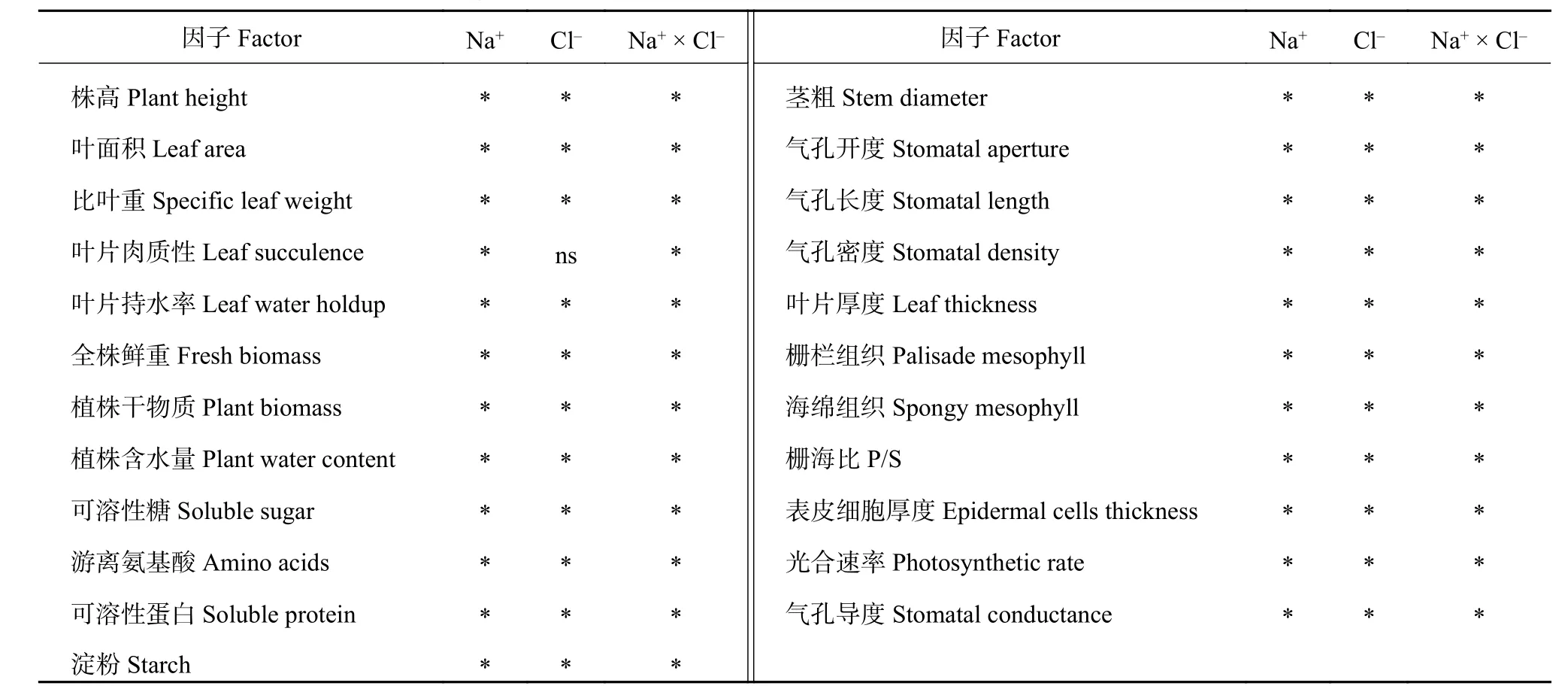
表3 Na+和Cl-平衡与非平衡处理下小白菜的主体间效应检验Table 3 Test of inter subjectivity effects of pakchoi under equal and unequal Na+ and Cl- concentration

表4 Na+和Cl-对小白菜不同部位叶片面积和单片叶干物质影响的效应检验Table 4 Test of Na+ and Cl- effects on area and biomass of pakchoi leaves at different positions
3 讨论
3.1 低浓度Na+和Cl-的生长有益效应呈现差异性
关于低浓度Cl-的生长有益效应试验表明,在根际营养液中伴随阳离子(K+、Ca2+、Mg2+)保持一致的条件下,通过阴离子(NO3-、SO42-、PO43-、Cl-)的对比试验观察到,5 mmol/L Cl-处理的烟草植株干物质、叶面积、Pn分别增加35%、45%、40%[4]。与之相类似,低浓度NaCl试验效应也呈现出植株干物质、植株形态、营养物质含量、Pn等指标的增加。王宝增等[12]将低浓度的NaCl (2.5和5 mmol/L)添加至Hoagland营养液中,小麦干物质量与Pn分别比对照增加了15.0%和10.0%与131.0%和123%。孟闯等[5]将低浓度的NaCl (5~10 mmol/L)喷施于叶片发现,黄瓜幼苗不仅干物质量(38.6%~50.2%)、植株含水量(20.8%~52.2%)、叶面积(12.6%~30.4%)和Pn(55.0%~35.0%)增加,而且全株可溶性糖(14.4%~38.3%)、游离氨基酸(30.2%~43.9%)含量提高。一些试验还观察到,根际营养液中添加低浓度NaCl处理下,番茄的单株果数增加[13],韭菜[2,14]和苋菜[3]的干鲜物质及营养物质、产量等均有显著增加。
本研究无论是采用[Na+]=[Cl-]的平衡处理还是[Na+]<[Cl-],或是[Na+]>[Cl-]的非平衡处理,对小白菜生长均呈现出与上述文献相类似的有益效应。然而,在阳离子(K+、Ca2+、Mg2+)的伴随下,单独Cl-处理(A1)有利于小白菜Pn和Gs提高;在阴离子(NO3-、PO43-、SO42-)的伴随下,单独Na+处理(C1)可显著提高小白菜可溶性糖(12.3%)与游离氨基酸(43.4%)含量。[Na+]与[Cl-]浓度平衡处理对小白菜的植株干物质(33.5%)、叶面积(30.3%)以及中上位单叶面积(26.1%~55.5%)等指标的促进效应最大;[Na+]<[Cl-]对小白菜光合效率Pn(17.3%)和Gs(145.3%)的促进效应最大,[Na+]>[Cl-]对小白菜游离氨基酸累积(52.6%)的促进效应最大。
3.2 低浓度Na+和Cl-的外源供给平衡性影响植株的水分积累和游离氨基酸累积
在本研究的预试验中,测定了叶施低浓度[Na+]=[Cl-]状态下全株各个器官Na+和Cl-的吸收量及Cl-/Na+值,初步观察到,虽然小白菜对Cl-吸收量的增幅比Na+更大,但是依然保持着对Na+的吸收量大于对Cl-的吸收量的状态。一般来说,植物吸收的Na+和Cl-会分布在各组织的质外体、共质体和液泡3个区域内[4,21-22]。在NaCl耐盐机制研究中,高浓度盐胁迫下(>100 mmol/L)植物体内通过保持细胞质内低的Na+浓度和高的K+/Na+值来保持Na+离子的内稳态[23]。一方面通过K+与Na+竞争降低Na+的吸收,另一方面通过Na+/H+逆向转运体提高Na+的外排或者将Na+区隔化到液泡中[24]。然而,Qiu等[25]在低浓度NaCl (10~30 mmol/L)试验中未观察到Na+/H+逆转运活性。在根际NaCl胁迫试验中,根系吸收的Cl-通过蒸腾拉力进入叶片[26],主要累积在液泡[27-28]或积累在叶绿体基质和类囊体内腔[29]。根际吸收Cl-和NO3-、SO42-存在拮抗作用[30],还在根细胞膜上发现Cl-和NO3-的共转运蛋白[31],也发现了H+/Cl-的共转运蛋白[22]。叶肉细胞中,Cl-则也会经MATE、CLC转运体的阴离子通道被区域化在液泡中[21]。在钠盐胁迫的调控机制和信号转导的大量研究中,关于Na+进出细胞质膜和液泡膜的转运蛋白(质膜Na+/H+逆向转运体SOS1、液泡膜Na+/H+逆向转运体NHX)及其基因表达,以及跨膜质子梯度驱动力(Na+外排的动力P型ATPase、Na+的区域化动力V型ATPase)等方面已有大量报道[6]。最近,介导Cl-流入(ZmNPF6.4和ZmNPF6.6)、Cl-流出 (AtSLAH3和AtSLAH1)和 Cl-区域化(AtDTX33、AtDTX35、AtALMT4和GsCLC2)转运蛋白的编码基因也逐步被证实[21]。
盐生植物通过维持细胞的渗透势,维持细胞质和液泡的水分用以提高生理耐旱能力[32]。在供水不受限制时,低浓度NaCl盐处理下,非盐生植物会通过降低细胞的渗透势,有利于液泡吸水以及细胞原生质体吸水膨胀甚至肿胀,增加细胞体积和扩展叶面积,表现出促进植物生长的有益效应[4,21]。
研究表明,植物体内K+的束缚水量较少,Na+的束缚水量比对照增加了2倍,Ca2+的束缚水量比对照增加了4.8倍;3个阳离子(K+、Na+、Ca2+)的效应比2个阴离子(PO43-和NO3-)对自由水和束缚水之比的影响更大[33]。Warne等[33]观察到,营养液中的NaCl在提高叶片肉质性方面比Na2SO4更有效,这说明Cl-的持水性更强。液泡内积聚有K+、Ca2+、Na+、H+等阳离子和NO3-、SO42-、PO43-、Cl-、有机酸 (如苹果酸)等阴离子。而Cl-作为非同化离子,其高浓度累积量与K+相近,是NO3-的4倍,是SO42-和PO43-的3倍[34]。这与Cl-激活液泡膜V型H+-ATPase活性[35-37]促进H+进入液泡所建立的电势梯度[38]下,有利于Cl-伴随进入液泡[39]的分室区域化累积有关。从而在液泡内Cl-累积形成更低的渗透势,促进水进入液泡[39],增加细胞膨压[40]。值得注意的是,Franco-Navarro等[4]和 Colmenero-Flores等[21]认为,与 NO3-、SO42-、PO43-等阴离子相比,Cl-在细胞渗透压调节、水分积累和细胞膨胀方面更有效;同时指出,Cl-在植物体内的水分动态平衡调节类似于大量元素K+,而不是Na+。Cl-与水的聚合度高,Cl-在提供细胞渗透压、水分积累和膨胀(植物细胞伸长的驱动力)方面更有效[22]。本研究在叶施低浓度Na+与Cl-平衡或非平衡处理下,观察到无Na+时Cl-有利于植株水分的累积,有Na+时Na+和Cl-均有利于植株水分的累积 (图2)。
另外,不仅NaCl盐胁迫下观察到叶片中脯氨酸等主要游离氨基酸积累[41-42];而且在12 mmol/L 低浓度NaCl盐处理下,游离氨基酸尤其是水解氨基酸累积,同时还观察到氮代谢中初级同化活性(谷氨酰胺合成酶Gs)大幅增加的现象[14]。孟闯等[5]在叶施低浓度NaCl处理时,观察到黄瓜幼苗全株及各器官游离氨基酸的累积。研究表明,Cl-作为微量元素直接参与天冬酰胺合成酶、淀粉酶活性的调节[43]。Colmenero-Flores等[21]推测Cl-似乎在韧皮部糖和可溶性氮(天冬酰胺)远距离转运的装载和卸载中起间接作用。无论如何,在Na+和Cl-从外部供给→吸收进入→体内转运→体内分布的物质流中,叶施低浓度Na+与Cl-的离子平衡及非平衡的外源供给,对相关转运蛋白的表达以及氮代谢的关键酶数量及活性的调节效应,尤其是对植物体内的“Na+与Cl-的代谢平衡态”的调节效应机制有待深入研究。
3.3 低浓度Na+和Cl-的外源供给平衡性影响着植株的干物质累积
一般来说,植物的干物质累积量与Pn、光合面积和光合时间密切关联[44]。Franco-Navarro等[4]采用根际营养液培养烟草观察到Cl-促进Pn和叶片面积增大效应。一般来说,根际供给的低浓度Na+或Cl-会随着蒸腾拉力经质外体长距离运输,到达叶片叶肉细胞及其液泡中累积,引起叶肉细胞充分吸水膨胀、体积增大和叶面积扩展增大[4,21-22]。然而,孟闯等[5]采用叶面喷洒低浓度NaCl的试验,也观察到促进黄瓜幼苗Pn和叶片面积的增加。本试验也采用叶施Na+或Cl-的处理方式,观察到类似的效应,且呈现出小白菜的植株干物质与叶面积的极显著相关(r=0.8226**)。值得注意的是,[Na+]=[Cl-]、[Na+]<[Cl-]和[Na+]>[Cl-]处理组的叶面积增加主要是中上位叶面积的增加(图4)。
高等植物的叶片,从茎顶端的叶原基分化之后,首先通过细胞分裂增加叶细胞的数量,至叶片展开后主要是细胞体积的增大,叶面积扩展会经历扩展初期(小叶)→扩展中期(叶面积迅速增加)→充分扩展期(功能叶)→叶片衰老的过程。本试验是在小白菜生长至3叶1心时开始叶施Na+或Cl-的处理。此时幼苗植株上叶片正处于已经展开后的充分展开期(从底数第1片叶)、扩展中初期(从底数第2~3片)和心叶未展开期。连续叶施Na+或Cl-处理21天后可见,下位叶(1~3片真叶)单叶面积均有轻微地变化;中位叶(4~6片真叶)、上位叶(7~9片真叶)的单叶面积均有显著增加(图4)。由此可见,叶施Na+或Cl-促进叶面积扩展效应,主要是有利于未展开期心叶的叶肉细胞体积增大。
显然,与根施处理方式不同的是,叶部喷洒NaCl处理方式下,Na+和Cl-会通过气孔渗入气孔腔室,通过质膜上的转运体进入气孔保卫细胞、叶表皮细胞和叶肉细胞,再通过液泡膜上的转运体进入液泡。
气孔的开闭与水分进出保卫细胞有关。保卫细胞从质外体吸收水分受K+、Ca2+、Cl-、NO3-、苹果酸根离子等流入的介导[44-45]。Franco-Navarro等[4]发现,根施Cl-处理方式下的气孔导度Gs会降低20%~36%,并认为这与叶片表皮的气孔密度降低有关。根施NaCl处理下,Na+或Cl-进入根系后,经过质外体长距离运输,到达最远端的叶片乃至叶片表皮的气孔保卫细胞,并且Na+或Cl-通过共质体内运输而分布在植物体的组织器官中。显然,根部处理下,Na+或Cl-优先累积在根和茎部的导管。Sotiras等[46]采用添加12、24和48 mmol/L的NaCl营养液培养枇杷幼苗42 天,观察到Na+在叶和根中积累,而Cl-在根和茎中积累。相比之下,本试验采用的叶部喷洒NaCl处理方式,Na+或Cl-首先接触的是叶片表皮及气孔保卫细胞。这种处理位点的变化,会引起Na+或Cl-的优先累积位点在叶片尤其是保卫细胞。有试验观察到,单一增加保卫细胞的Cl-,会直接增加保卫细胞的水进入引起膨压增大[21]。孟闯等[5]在叶施低浓度NaCl处理时,观察到黄瓜幼苗叶片气孔导度的增加效应。本试验在叶施低浓度Na+与Cl-的平衡及非平衡处理下,观察到Cl-比Na+更有利于小白菜叶片气孔导度的增加(图6),可是与之相反的是Cl-尤其是Na+更有利于促进气孔开度(图5)。这是否与非平衡处理中与Cl-伴随的阳离子K+、Ca2+、Mg2+或与Na+伴随的阴离子NO3-、PO43-、SO42-的差异有关,尚待研究。
4 结论
Na+与Cl-供应浓度平衡与否都对小白菜的生长、干鲜物质累积、叶片光合活性具有促进效应。较高的Na+更有利于游离氨基酸的累积,较高的Cl-更有利于干鲜物质的累积以及光合活性的增加。在Na+与Cl-供应浓度相同时,最有利于叶片主要是中上位叶片面积的扩展,因而提高植株干物质累积的效应最大。本试验中喷施Na+和Cl-浓度均为12 mmol/L对小白菜生长的促进效应最优。
