大豆和玉米影响后茬作物氮素供应的研究进展
2022-06-17耿赛男李岚涛苗玉红谭金芳王宜伦
耿赛男,李岚涛,苗玉红,谭金芳,王宜伦*
(1 河南农业大学资源与环境学院,河南郑州 450002;2 中山大学农学院,广东广州 510000)
小麦、玉米是广泛种植的粮食作物,世界上以小麦、玉米为主食的人口分别约有40%和33%,中国是小麦、玉米生产和消费大国[1-2],玉-麦轮作是我国重要的一年两熟集约种植模式[3]。氮是植物所必需的三大营养元素之一,外源氮肥投入是植物获取氮素的核心途径。小麦、玉米周年氮素需求量大,我国粮食作物的推荐平均施氮量不超过180 kg/hm2[4],但我国玉-麦轮作主产区周年氮肥平均用量达545 kg/hm2,部分高产粮区甚至超过600 kg/hm2[5],玉米生育期较短,但施氮量占周年氮肥用量的二分之一以上。中国集约化农田中高氮肥施用面积已达农田总面积的15%以上[6]。过量氮肥施用导致氮肥利用率低,土壤结构板结,土壤有机质含量下降,同时未被吸收的部分会进入土壤、大气和水体污染生态环境[5]。因此,在集约化农区,如何建立绿色可持续发展的轮作模式尤为重要。
大豆的蛋白质和脂肪含量丰富,是世界各国重要的粮食贸易作物,是我国五大粮食作物之一[7],大豆-小麦轮作也是我国重要轮作模式。豆科作物自身具有较强生物固氮能力,我国大豆种植区的生物固氮量最高可达150 kg/hm2[8]。Herridge等[9]认为,全球豆科作物年固氮量为1.3×108t,因此,大豆等豆科作物对氮肥依赖性低。此外,豆科作物还具有为后季作物提供氮素、降低其对氮肥的依赖性及提升其产量效益等。Vance[10]指出,后季作物中约有100 kg/hm2的氮来自前季豆科绿肥;Gan等[11]通过3年田间试验表明,与豆科作物轮作后,后季小麦产量、蛋白质含量和肥料利用率分别提高35.5%、50.9%和33.0%;减少豆科作物种植面积后,法国小麦产量15年内年平均降低0.035 t/hm2[12]。因此,深入探索轮作体系中豆科作物对后茬作物氮素供应机制的影响,对于降低氮素投入、提升氮素利用效率和实现作物优质高产等具有重要的理论和实践指导意义。综合国内外研究进展,以豆-麦和玉-麦轮作模式为例,从作物氮素需求特性、土壤物理特性,残留氮素转移和转化特性等方面,综合阐述豆科作物在轮作体系中的地位与作用,以期为进一步将豆科作物纳入轮作体系,实现集约种植模式氮肥优化管理和绿色可持续发展提供依据。
1 玉-麦和豆-麦轮作体系前茬作物氮素供需特征
1.1 玉米氮素需求、氮肥施用特征
2010—2020 年间我国粮食作物种植面积平均稳定在11563万hm2,玉米种植面积占1/3左右[13]。每生产100 kg玉米籽粒,需从土壤中吸收纯氮2.9 kg[5]。拔节—大喇叭口期、抽雄—灌浆期、乳熟—蜡熟期是玉米主要的氮素吸收高峰期[14],抽雄前营养器官中积累氮素的转移和根系的氮素吸收是玉米籽粒中氮的主要来源[15]。外源氮肥是保证玉米增产的重要手段,当前玉米生产中主要采用无追肥的一次性施肥技术,2008年调研玉米种植大省吉林,玉米一次性施肥技术最高推广地区已经占到了75%以上,且近年来在全国也迅速发展[16-17]。由此,肥料氮释放和玉米氮需求同步性不高是限制玉米增产与氮肥利用的一大障碍。由于土壤、气候和品种等因素影响,夏玉米适宜氮肥用量差异较大,我国粮食主产区黄淮海平原夏玉米适宜施氮量在113~180 kg/hm2[18-19],氮肥不合理施用将不利于玉米增产和养分高效利用。统计表明,1979—2016年间全国化肥、氮肥和复合肥施用量整体呈稳步增长趋势,但粮食产量并未同步持续增加(图1)。农业生产中氮肥施用过量现象普遍存在,有时甚至超过“作物特异性适应”范围的上限,造成“高氮胁迫”[20]。氮肥平均利用率世界范围内约为33%[21],西欧发达国家可达42.6%~57%[22],我国主要粮食作物氮肥当季利用率平均为27.5%,其中玉米约为26.1%[23]。施入过量氮肥不仅使增产效应降低,且损失的氮素会进入大气、土壤,造成面源污染[24]。在高施氮量条件下,玉米季残留氮除以微生物氮、有机氮形式存在外,以NO3--N形态存在的占比也较高,无形中增大了残留氮素淋溶损失的风险[25]。

图1 中国粮食、小麦、玉米、大豆产量及肥料施用量变化趋势[13]Fig. 1 Trends in grain, wheat, maize, and soybean yield and fertilizer application in China
1.2 大豆氮素需求、氮肥施用特征
近20年来,我国大豆种植面积约维持在830×104hm2[13]。大豆是高需氮量作物,生产100 kg籽粒需氮量为7.0~9.5 kg,是小麦、玉米的2~3倍[5,26]。大豆开花前需从土壤中吸收充足氮素以保证营养生长,对氮肥需求主要集中在结瘤之前,施氮量一般为18~90 kg/hm2[27],远低于玉米,因此豆科作物可直接减少轮作体系周年氮肥施用量[11,28]。根瘤固氮和土壤氮是大豆主要氮源,研究表明根据植株的需要,根瘤中的固氮酶会调整自身活性[29-30]。大豆、花生、豇豆、扁豆、豌豆等常见食用豆类作物的固氮量最高值可达植株总吸收氮的70%以上,平均固氮量可达200 kg/(hm2·a)以上,其中大豆固氮量最高可达450 kg/(hm2·a) (表1)。生物固氮占全球固氮总量的70%,其中豆科植物与根瘤菌形成的共生固氮占生物固氮总量的60%以上,是农业生产中的重要氮源[32]。因此,种植豆科作物氮肥投入很小,巴西大豆增产主要依靠生物固氮,早年间美国就靠种植苜蓿、大豆、花生等豆科作物所固定的氮间接减少了氮肥施用量[32-33]。

表1 常见豆科作物固氮量及其所占植株吸氮量的比例[31]Table 1 Amount of plant nitrogen derived from N2 fixation by leguminous crops and its proportion to plant nitrogen uptake
2 豆-麦轮作体系下豆科作物增产效应
豆科作物有提升后季谷物产量的效果,后季谷物产量增幅为3%~73% (表2)。在同一地块中,大豆茬口冬小麦产量较玉米茬口增加5.12%~17.05%,且冬小麦经济最佳施氮量和最适施氮量分别比玉米茬口减少19%和22%[54]。豆科作物对后茬谷物的增产优势,主要源于其促进了土壤氮素的增加[55-56]和氮素形态的转化[57]。Brophy等[57]研究证实豆科植物通过根系分泌相当数量的氮化合物,这些化合物以无机氮、小分子有机氮(氨基酸)、细胞溶解物、脱落物等形态在根际积淀[58-59],根际沉积物中的氮主要以化合物形态存在,有时也会以铵态氮、硝态氮等离子形式释放[60-61]。根际沉积氮是作物庞大的地下氮库,豆科作物根际沉积氮显著高于禾本科作物[62]。根际沉积氮易于为后茬作物吸收利用,研究发现,禾本科作物地上部超过30%的氮素来源于前茬大豆根际沉积氮[33]。在撒哈拉以南的非洲种植系统中,豇豆收获后留在土壤中的氮素高达25 kg/hm2,其中有40%存在于0—30 cm土壤中,可以为后茬作物吸收利用[63]。与玉米季残留氮素相比,豆科作物残留的氮素不容易损失,为后季作物的吸收提供了可能性[64]。因此,在以谷物为基础的种植体系中引入豆类作物,对提高生产力,实现粮食安全生产和可持续发展具有重要意义[65]。
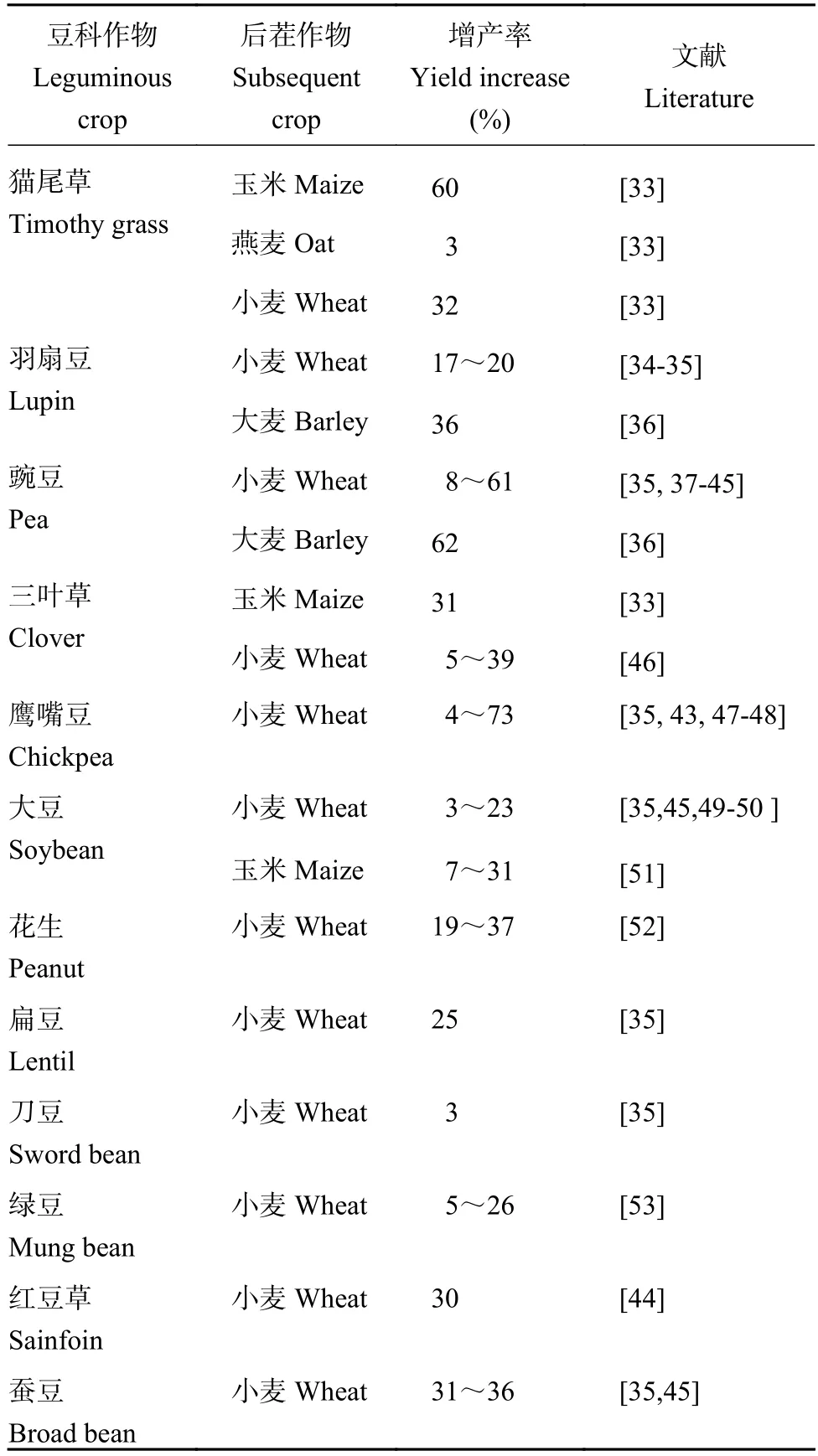
表2 轮作体系中不同豆科作物对后茬作物的增产率Table 2 Yield increasing effect of different legumes on the subsequent crops in rotational systems
3 大豆和玉米影响后茬小麦氮素供应差异分析
3.1 豆科作物有效改善土壤团聚度
大豆根系呈网状分布,主根有较多分枝侧根,生长过程中具有新老根频繁更替的特点。大豆收获后土壤颗粒排列疏松,土壤团聚体呈椭圆形、圆形,其形态由原来简单形态转变为多级复合形态,并且团聚体内部空隙明显增多,团聚体之间的空隙由原来裂隙状变为树枝状和网状[66-67]。大豆根系创造的多级空隙有利于减少后季作物根系伸展的机械阻力,且土壤有效水分的残留可为后茬小麦利用,促进其生长发育[68]。大豆根系生长过程中形成的多级复合团聚体是大豆根分泌物、腐解物和脱落物的吸附载体[67]。大豆收获后土壤中既有被粘粒吸附的也有未被吸附的腐解物,还有因挤压被封存在两个团聚体之间的腐解物,不同的粘粒吸附物分解速率不同,并且粘粒吸附的物质比未吸附的物质分解慢得多,因此大豆季土壤肥沃,潜在肥力可延续到后季作物生育期[68-69]。与大豆不同,玉米收获后土壤透光性差,土壤颗粒排列紧密,团聚体多为根系穿插挤压所形成的简单团聚体,以裂隙状为主要存在形态,团聚体内部空隙较少,彼此间连通性较差[70]。
Six等[71]和李鉴霖等[72]认为土壤中最好的团粒结构直径>0.25 mm,>0.25 mm的大团聚体含量越大,团聚体稳定性指数越高,越有利于土壤团聚体的分布和稳定。大豆生长过程中根系一方面能直接作用于土壤,影响其团聚体的形成特征与稳定性,另一方面大豆生长过程产生的根系分泌物也可以影响土壤团聚体。王国义等[73]将干筛法得到的1~2 mm团聚体用大豆根系代谢培养液进行处理,发现有14%可以成为水稳性团聚体,达到显著水平 (P<0.05)。另有研究指出与玉米相比种植大豆一年后,就可获得良好的耕层结构[67]。因此,豆科作物可有效改善土壤团聚度,研究表明土壤团聚体粒径分布与土壤矿化强度显著相关[74]。
3.2 豆科作物有效提高后茬土壤有效氮素供应
土壤氮素循环转化过程以氮素同化、异化、氧化和还原为中心的生物转化过程占据重要地位(图2)。土壤起始矿质氮和作物生长期间经氮素转化过程产生的矿化氮,决定土壤对作物的氮素供应能力。作物生长过程中无论施肥与否,其吸收的氮素有50%以上来自于土壤[75]。因此,作物生长期间土壤氮素供应水平与有机氮矿化产生无机氮的能力密切相关[76]。土壤有机氮主要分为酸解态氮与非酸解态氮。酸解态氮包含酸解未知态氮、氨基酸氮、氨态氮和氨基糖氮,有机氮不同分布形态是影响土壤氮素有效性的重要因子[77]。土壤中可矿化氮主要来源是盐酸可酸解态氮,尤其是铵态氮和氨基酸态氮[78]。土壤微生物氮是与土壤有机氮接触最为紧密的氮形态,土壤微生物不断将土壤有机氮矿化成铵态氮,在碳源充足情况下又可将氮用于繁殖构成新微生物体。因此,土壤微生物是土壤氮素固持和矿化的执行者,在土壤氮转化中起着重要作用[78]。研究发现0—10 cm耕层土壤氮总矿化量约为120 kg/hm2,净矿化量为59 kg/hm2,其中净矿化量69%来自土壤微生物氮库[64]。
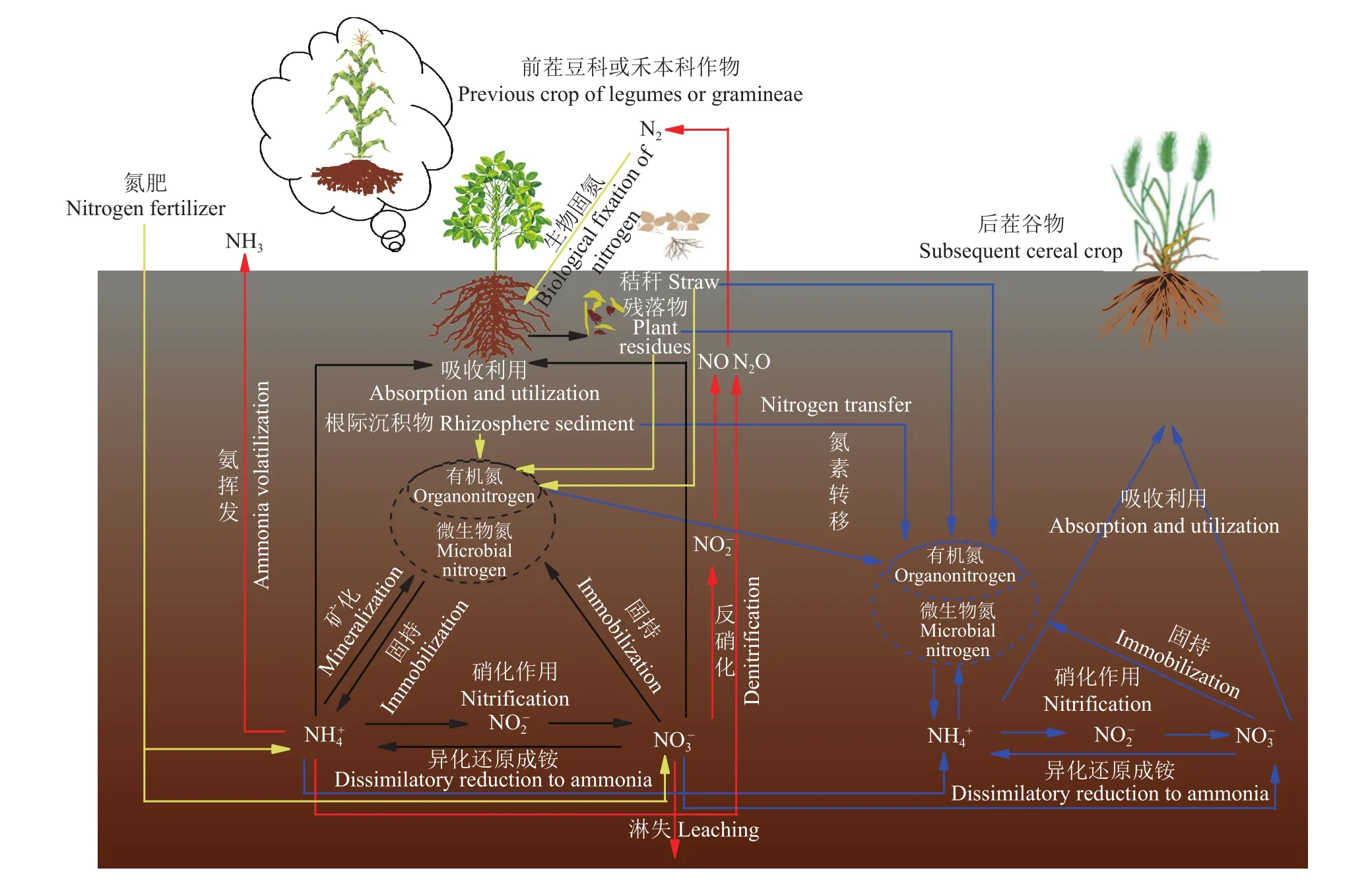
图2 豆科-禾本科轮作系统下土壤氮素的循环和转移示意图Fig. 2 Diagram of nitrogen cycling and transfer in farmland under legumes-cereal crop rotation system
农田生态系统氮素“源—流—库”变化显著受作物种类影响[78]。Nyfeler等[79]研究表明,种植三叶草可显著提高土壤氮初始矿化率,具有增加后茬作物有效氮库潜力。李可懿[80]研究证实当前茬栽种豆科作物时,次年土壤矿质氮含量显著提高,这主要源于前茬豆科作物有效提高了土壤中有机氮的矿化和转运[76]。Mayer等[81]研究表明,轮作体系中加入豆科作物可明显影响后茬作物的氮素吸收量,其吸收量高低取决于豆科植物的残留氮输入量。Plaza-Bonilla等[82]研究指出豌豆可为后季小麦作物提供40~49 kg/hm2的氮。Guinet等[35]研究认为,相比禾本科作物,豆科作物豌豆、羽扇豆、箭筈豌豆、扁豆、蚕豆可分别增加后季小麦地上部氮含量7.94%、9.52%、15.87%、20.63%和28.57%。来自澳大利亚的羽扇豆-小麦轮作试验表明,羽扇豆3.00%~10.00%的地下残余氮可被后季小麦有效利用,且后茬作物矿质氮的30%~75%来源于前茬豆科作物的积累[64,83]。综合分析前人研究进展发现,轮作体系中豆科作物残留氮素是土壤易矿化氮的主要来源,可显著增强土壤氮素矿化潜力,提高土壤矿质氮含量,增强后季作物氮素吸收利用能力。豆科作物不仅能提升后茬土壤氮素供应能力,而且豆科作物收获后残留物中碳的含量低且不稳定限制了反硝化作用,间接降低了N2O的周年排放强度[84],已有研究表明,反硝化作用是大多数农业土壤中N2O的主要来源[85]。此外,全球每年约60%的温室气体排放来自农业[86],主要是由化肥投入引起的[87]。因此,低氮肥投入的豆科作物直接降低了N2O的周年排放强度。Parkin等[88]研究表明,玉米季的N2O排放为大豆季的约两倍。Neto等[89]研究明确,玉米-小麦轮作体系中N2O的排放量比大豆-小麦轮作体系高25%。因此,轮作体系中引入豆科作物对农业生态系统健康可持续发展意义重大。
4 轮作体系氮素转移的机制研究
4.1 残留氮肥补充土壤氮库
氮肥施入土壤后的去向主要包括作物吸收、淋洗、挥发损失、土壤残留等途径(图2)。残留肥料氮通常占施氮量的12.0%~40.6%[90-91]。许多田间试验结果表明,随着施肥量增加,作物产量与氮肥当季利用率并没有同步提高,经常出现后茬作物施氮不增产或者低施氮量获得高产现象[92-93],表明了氮肥的后效。在轮作体系中,前茬作物种类不同,其施肥种类和量、氮素土壤残留量均有差异,氮肥后效也因此不同。利用15N标记尿素进行研究表明,作物收获后土壤中残留的氮肥以NO3--N、NH4+-N、固定态铵、微生物氮及其它多种有机形态存在于土壤中,在高施氮量条件下,残留氮肥主要以微生物氮、NO3--N和其他有机态氮存在,尽管微生物氮与NO3--N都是土壤中活性很强的氮库,但由于在土壤中不稳定,极易发生损失,因此,不利于后茬作物的吸收利用[25],后季作物对前茬残留氮肥利用率仅为2.4%~5.2%,一般不超过8%[25,94]。前茬氮肥残留在土壤中的氮素除通过淋失、挥发等途径损失外,还有部分补充到土壤氮库[90-91],短期内不易于被后茬作物直接吸收利用,但是经长期合理施肥后,这些氮素依然会对后季作物的氮素营养及产量产生正面的效应。
4.2 大豆秸秆还田促进土壤氮素循环
轮作体系中的氮素可以通过作物残落物、秸秆和根际沉积等途径转移(图2)。秸秆(残落物、残茬等)还田后,其释放的氮是土壤氮素重要的源和库[95]。秸秆等进入土壤后,其含有的可溶性有机碳、蛋白质、纤维素等一些易氧化有机物可被微生物快速矿化释放养分;残留的较难分解物质以及矿化过程产生的中间产物,在微生物作用下形成结构复杂难分解的腐殖质导致秸秆矿化速率减缓,这部分物质可以补充土壤有机质[96-97]。秸秆种类组成及环境因子影响秸秆腐解矿化过程,进而影响其对后季作物的氮素营养,最终影响作物产量[98]。大量研究表明,秸秆还田的后季作物产量增幅为0.90%~22.70%,土壤全氮增加幅度为2.61%~45.52%,土壤有机质提升1.60%~30.00% (表3)。因此,有效的秸秆还田可以成为提高耕地潜在氮库的有效方式[119]。

表3 不同秸秆还田对土壤和后季作物的效果Table 3 Effects of straw returning on soil and subsequent crops
C/N是影响微生物氮素转化的核心因子,低C/N的秸秆更易腐解[120-121]。还田秸秆C/N较大时,腐解初期释放的氮素首先进行生物固定,随着腐解进程推进,秸秆C/N逐渐降低,易被微生物获取的能源物质也逐渐被消耗殆尽,部分微生物死亡释放出被其固定的氮素。因此,C/N低的作物残体更利于土壤微生物生长,加快土壤氮素循环[122]。一般认为土壤微生物分解有机物质的最适宜C/N在25~30[116]。研究表明,C/N较低的大豆秸秆在翻压后前16周就已经达到较高腐解度,而谷物秸秆在翻压16周后仍然保持较高分解速率[123-124]。低C/N的大豆秸秆还田后有利于微生物活动,这是其在不适宜环境条件下仍保持较高分解率的主要原因[123]。因此,与谷物作物相比,大豆等豆科作物秸秆还田后,更有利于土壤氮素循环。
4.3 豆类作物根际沉积氮是后续作物重要氮源
龚振平等[125]研究指出,种植大豆也会造成土壤氮素含量下降,造成土壤氮素亏损的主要原因是大豆氮素收获指数高,成熟期转移到籽粒中的氮素为70.4%~88.6%。在大豆秸秆还田与不还田情况下,土壤氮素亏损量分别约为39.2 和 49.2 kg/hm2。持相反观点的研究认为,豆科作物能增加土壤矿质氮和全氮含量,减少后季作物氮肥使用量不会造成减产[10,54-55]。迟凤琴[126]研究表明,种植大豆后土壤有机质和全氮含量分别增加0.46%和0.6%,而种植玉米土壤有机质增加0.34% (较大豆降低26.09%),全氮减少5.2%。这些研究差异主要源于通常被忽略的根际沉积氮。研究表明,小麦对蚕豆和鹰嘴豆地下氮利用率分别为8%和15%,如果不考虑地下氮,小麦对蚕豆和鹰嘴豆可回收部分氮利用率只有3%[64]。常见豆科作物根际沉积氮为6.00~94.20 kg/hm2,占植物体总氮含量的4%~71% (图3)。根际沉积氮不受氮肥限制,与根生物量和根系氮含量呈正相关,根生物量增加1g,根际沉积氮增加53 mg左右[127]。根际沉积氮占地下氮素总量(根系氮素含量+根际沉积氮)的47%~85%,占总残留氮的35%~45%[63,81]。在澳大利亚的试验表明,小麦可利用前茬羽扇豆地下氮的3%~10%[128],在尼日利亚几内亚大草原和北欧草原上的研究表明,大豆等豆科作物地下氮的贡献是农业中氮的主要来源[129,133]。综合前人研究进展发现,小麦可利用13%~85%前茬豆类根际沉积氮,这占小麦吸氮总量的4%~20%[63,127-128]。

图3 常见豆科作物根际沉积氮量及占植柱总氮的百分比[33,62,81,127-132]Fig. 3 Amount and percentage of nitrogen deposition in the rhizosphere in total plant nitrogen of common legume crops
豆科作物根际沉积氮量是禾本科作物的2倍左右[33]。Fustec等[130]研究表明根沉积氮的主要途径是:1)根瘤和根细胞的衰老、死亡、分解和腐烂;2)可溶性化合物的渗出;3)根缘细胞脱落;4)粘液分泌。根瘤和脱落根系产生的氮是土壤沉积氮的重要来源[64]。大豆生长过程中具有频繁新老根更替特点,衰老、死亡的根量高[67]。此外,根系释放的氮大多为低分子量物质,如可溶性根系分泌物、氨基酸、激素和酶,发现的根系分泌物氨基酸中甘氨酸和丝氨酸的回收率通常很高,使用15N标记氨基酸表明,豆科植物根中甘氨酸和丝氨酸的排出量高于禾本科作物[130]。其次,根际沉积物C/N不同,豆科作物沉积物具有较高的氮含量和较低的C/N[76]。用丰度为99.0%的15N标记前茬根际沉积物,在后季小麦中测出来自前茬豆科和禾本科作物的根际沉积氮分别为14.4%和8.5%。因此,低C/N的豆科作物根际沉积物比谷物作物根际沉积物更易矿化,利于改善土壤氮素供应效能[134]。综上所述,豆科作物的根际沉积氮更容易被后茬作物吸收利用,是后续作物的重要氮源,具有推动农业可持续发展的潜力。
5 展望
与豆科作物轮作一直是农业绿色可持续发展的一个有效措施,影响轮作体系效益的土壤与作物因素很多,因此,要实现轮作体系的高效,还应从多方面去开展进一步研究。
5.1 轮作体系中土壤氮素的周年供应特征及主要驱动因素
目前国内外有关豆-麦轮作体系小麦产量效益的研究相对较多,而有关大豆残留氮在后季替代化肥氮比例的报道则相对较少。不同轮作体系会打破周年氮素循环与转化特征,探明豆-麦轮作体系中氮素有效化驱动因子的作用机制,构建相关氮素转化和氮素平衡模型,明确残留氮素在小麦季替代化肥氮当量对小麦产量和氮肥利用效率的影响,这对构建豆-麦轮作体系周年氮素高效综合管理策略意义重大,目前这方面的研究还很缺乏。
5.2 轮作体系中氮素高效利用的根-土-微生物机制
农田氮循环是由微生物驱动的一系列复杂过程,在探索大豆根系结构和根际沉积物多样性对氮素生物有效性和土壤微生物影响的基础上,深入揭示不同轮作体系下根-土-微生物调控氮素的互作机制,对阐明豆-麦轮作体系氮素高效利用的根际调控途径具有重要意义,而目前关于豆-麦轮作体系中小麦季氮素高效利用的微生物机制仍不明确。
5.3 豆-麦轮作降低温室气体排放的潜力
农田土壤是活跃的有机碳库,也是温室气体主要的“源”和“汇”。秸秆还田是提高土壤肥力与可持续生产力以及增加土壤固碳量的重要途径,然而土壤有机质积累会引起增加CO2等温室气体排放的风险,受研究技术限制,目前对秸秆还田“净排放”能力的研究主要借助模型估算,但估算结果和实际情况间存在差距。并且,由于农田土壤N2O等温室气体排放过程复杂,轮作系统间温室气体排放的差异机制,豆-麦轮作体系中土壤生物和理化性状的改善对温室气体排放的影响等系列问题也尚待解决。近年来,分子生物学和微生物学等创新性技术的发展,将有助于推进豆-麦轮作体系温室气体减排和作物增产高效的同步发展。
