不同根肿病抗性油菜根系分泌物的代谢组研究
2022-06-17陈宇红蒋学飞王茂林
陈宇红, 蒋学飞, 苏 滢, 王茂林
(四川大学生命科学学院 生物资源与生态环境教育部重点实验室, 成都 610064)
1 引 言
十字花科根肿病是由芸薹属根肿菌(PlasmodiophorabrassicaeWoronin)引起的土传性病害,包括拟南芥,白菜,油菜等在内的众多十字花科植物或经济作物均会罹患此病,每年都会造成巨大损失,而且随着现代农业的发展,根肿病的发病面积和危害程度也在逐年增加[1],因此对根肿菌的研究和对根肿病的防治已经到了刻不容缓的地步.
囿于根肿菌的寄生特性,现阶段对于根肿菌的认识水平仍较低,目前研究者普遍认同将根肿菌的生活史可分为三个阶段:根毛侵染,皮层侵染以及休眠孢子时期[2]. 首先,土壤中的休眠孢子在有合适寄主存在的情况下会被某些特定的刺激物诱导萌发,形成具两根不等长鞭毛的初级游动孢子[2]. 而后这些初级游动孢子泳动或吸附到寄主的根毛上,形成根毛感染. 在根毛中,初级孢子囊发育成熟为次级游动孢子并再次释放到环境中,等待进入皮层侵染阶段[3]. 皮层中柱的次级孢子囊不断分裂,扩增,最终形成休眠孢子充满寄主的根部细胞,待宿主腐烂重新释放到土壤中,直到下一轮侵染开始[4]. 初级游动孢子一旦侵入宿主,就可以隔绝外部的不利环境[5],在土壤中的休眠阶段,孢子又具有一层厚厚的几丁质,可保证其在各种恶劣的土壤环境中存活很长时间[6],因此,根肿菌的孢子在萌发之后但未成功入侵寄主的这个短暂时期对于根肿病的发病和预防而言都很关键. 已有证据表明根肿菌在这个阶段是最脆弱的[5],利用此特征的根肿病防治办法也有报道,例如,选择非寄主作物进行轮作,再次种植寄主植物发病率可显著降低,或种植易感的寄主作物,在发病期用药或处理土壤都可显著降低后续作物的发病率[7,8]. 因此,探究影响休眠孢子萌发的因素具有重要意义.
休眠孢子的萌发受许多因素的影响. 首先,休眠孢子在有宿主存在的情况下会大量萌发,但在没有宿主的情况下也会少量萌发,这种萌发可能是受到合适的土壤湿度和温度刺激. 休眠孢子的萌发也受到pH值影响,在pH值为5.0至7.0时,休眠孢子的萌发率较高,在pH值大于7.0时萌发率较低[9],而在更高pH值下萌发率被抑制得更加明显[10].休眠孢子萌发的最佳温度为24 ℃,最高致死温度为45 ℃,可见光会抑制萌发[5]. 另一方面,植物尤其是寄主植物对休眠孢子萌发的研究也由来已久,Macfarlane(1970)很早便发现根系分泌物能够刺激休眠孢子的萌发,且这种刺激作用与根系分泌物的来源密切相关[10]. 首先,十字花科宿主作物的根系分泌物比非宿主植物更有效,一些非十字花科植物宿主,如多年生黑麦草(LoliumperenneL.)、红三叶草(TrifoliumpratenseL.)、果岭草(DactylisglomerataL.)和翦股颖(AgrostisstoloniferaL.),根系分泌物比非宿主植物的作用强,有时甚至比十字花科宿主更有效[9,11]. 对于同一物种,易感材料的根系分泌物比抗性的更有效,例如,用来自易感甘蓝(B.oleraceavar.capitata)的根系分泌物处理后,休眠孢子的萌发率最高(75%)[12]. 这些研究表明根系分泌物中或有一些与根肿菌休眠孢子萌发密切相关的代谢物,它们的存在和含量高低直接决定了刺激休眠孢子的萌发的效率,但目前关于这种刺激物的研究却鲜有报道,而通过对比寄主非寄主,抗病和易感植株的根系分泌物组分可以帮助我们寻找这种关键的代谢物.
非靶向代谢组技术,是一种不局限于预先确定的已知代谢物,可以发现更多与植物代谢有关新物质的方法. 以前检测植物代谢物的常用方法是气相色谱质谱联用(GC-MS)[13,14],虽然这是分析小型极性代谢物的最佳方法,但由于需要对每个分子进行化学修饰以增加挥发性,因此无法检测大多数较大的植物次生代谢物,如复杂的生物碱和甘油磷酸脂等. 与GC-MS相比,超高效液相色谱与质谱联用(UPLC-MS)是一种非常适合大规模调查生物系统代谢物的分析方法[15]. Kc84-1是在之前的工作中筛选出的具有较好的根肿病抗性的甘蓝型油菜材料,华双5R是已获广泛推广种植的抗根肿病材料,R-197是易感材料. 在本研究中,我们利用UPLC-QTOF-MSE技术,结合多变量数据分析模型,对三个不同根肿病抗性水平的甘蓝型油菜根系分泌物中的次级代谢物进行检测鉴定和比较分析,筛选出一些差异代谢物. 并进一步验证差异代谢物与孢子萌发之间的关系,以及萌发率和根肿病发病率之间的关系,为油菜根肿病发病机理的研究及病害防治提供了一定的资料.
2 材料与方法
2.1 材 料
甘蓝型油菜(BrassicanapusL.)材料:根肿病易感材料R-197;根肿病抗性材料华双5R(华中农业大学张椿雨老师赠予)和Kc84-1(本实验室保有).
2.2 方 法
2.2.1 休眠孢子的制备 取保存于-20 ℃的根肿搅碎,纱布过滤后离心,先3100 r/min离心15 min,弃上清液,沉淀用蒸馏水悬浮后再3100 r/min离心15 min,重复3次. 弃上清,用50%蔗糖溶液悬浮沉淀物,3100 r/min离心10 min,将上清转移至新的50 mL无菌离心管中,补无菌水至30 mL,离心,弃上清,洗涤2遍,用2%的新鲜氯胺-T处理20 min,而后用抗生素(1000 μg/mL硫酸粘杆菌素,1000 μg/mL盐酸万古霉素,6000 μg/mL头孢噻肟钠)处理过夜,无菌水洗3遍,检查除菌情况. 4 ℃保存备用.
2.2.2 根系分泌物的制备 甘蓝型油菜种子经1%的NaClO表面消毒8 min,无菌水洗3次,播于1/2 MS固体培养基中萌发5 d. 将石英砂淘洗干净,并180 ℃干热灭菌3 h,冷却后分装至培养皿,每个培养皿50 g石英砂,而后将装有石英砂的培养皿再置于蒸汽灭菌锅中121 ℃,30 min,冷却后每个培养皿加入30 mL无菌水(pH 6.3). 将萌发的油菜幼苗分装于装有石英砂的培养皿中,每个培养皿20株(3种材料,每种6个重复). 将此装置置于光照培养箱中(16 h/8 h;24 ℃/20 ℃;光照/黑暗;相对湿度50%). 10 d后,将植物移出,收集根系分泌物,经0.22 μm滤膜过滤. 部分立即用于休眠孢子的萌发实验,取部分(20 mL)用液氮速冻,储存于-80 ℃,用于超高效液相色谱-质谱联用(UHPLC-MS)分析.
2.2.3 休眠孢子的萌发及鉴定 将无菌的休眠孢子用新制备的根系分泌物或无菌水(pH 6.3)调整浓度为每毫升1×107个孢子,200 r/min 25 ℃摇床黑暗孵育. 每24 h对各处理的休眠孢子进行萌发率调查. 用醋酸地衣红染色,在Nikon DS-Ri2显微镜下对每个样品300个孢子进行计数分类. 每次测量重复3次. 使用OriginLab对数据进行方差分析,使用Duncan多重检验(P≤ 0.05)评估每个时间点处理之间的差异.
之前的研究已经建立了一个检验根肿菌休眠孢子萌发率的方法,即在显微镜视野中孢子萌发的第一个特征是休眠孢子所特有的折射球消失,孢子的萌发可以通过这种方式区分[10, 16]. 而用醋酸地衣红染色后这种差异更易区分,即萌发的孢子是不被着色的(图1a中白色箭头所示),而未萌发的孢子的会被染上紫红色(图1a中黑色箭头所示).
2.2.4 根系分泌物的UPLC-MS分析 样品的衍生和UHPLC-MS分析是基于Chen等的方案[17]. 根系分泌物的检测使用:AB Sciex Triple TOF 5600高分辨质谱仪与Agilent 1290超高效液相色谱仪和Waters ACQUITY UPLC BEH Amide柱(1.7 μm × 2.1 mm×100 mm). AB 5600 Triple TOF质谱仪能够在控制软件(Analyst TF 1.7, AB Sciex)控制下基于IDA功能进行一级、二级质谱数据采集. 在每个数据采集循环中,筛选出强度最强且大于100的分子离子进行采集对应的二级质谱数据. 轰击能量:30 eV,每50 ms 15张二级谱图. ESI离子源参数设置如下:雾化气压(GS1):60 Psi,辅助气压:60 Psi,气帘气压:35 Psi,温度:650 ℃,喷雾电压:5000 V(正离子模式)或-4000 V(负离子模式). 流动相包括溶剂A(25 mmol/L醋酸铵和25 mmol/L氨水)和溶剂B(乙腈). 以0.5 mL/min的流速梯度进行:溶剂A和B在0 min到0.5 min时分别保持5%和95%,溶剂A从0.5 min的5%线性增加到7 min的35%,到8 min的60%,并保持在60%直到9 min. 最后,溶剂A在9.1 min时降至5%,并保持在3%直到12 min. 溶剂B从0.5 min的95%线性下降到7 min的65%,到8 min的40%,并保持在40%直到9 min. 最后,溶剂A在9.1 min增加到95%,并保持在95%直到12 min. 使用ProteoWizard软件将质谱原始转成mzXML格式. 再使用XCMS做保留时间矫正、峰识别、峰提取、峰积分、峰对齐等工作,minfrac设为0,cutoff设为0.6. 与自建的二级质谱数据库对峰进行物质鉴定.
通过比较总离子色谱(TIC)进行数据质控,为了识别代谢物,我们对以下信息进行了综合分析. (1)使用ProteoWizard软件将质谱原始转成mzXML格式;(2)获取标准化合物的准确质荷比(m/z)和保留时间;(3)将上机的代谢物参数矫正和比对之后与数据库进行比对和物质鉴定;(4)ESI+和ESI-模式分别进行.
2.2.5 数据分析 对结果进行主成分分析(Principal Component Analysis,PCA)和正交偏最小二乘法-判别分析(Orthogonal Projections to Latent Structures- Discriminant Analysis, OPLS-DA). 同时,对代谢物数据进行了对数转换,以改善正态性,并进行了归一化处理,用于统计分析. 对三个油菜材料进行分组,对于差异性代谢物的筛选,采用了学生T检验的P值和OPLS-DA模型的VIP值.P< 0.05,VIP> 1.
2.2.6 差异性代谢物的验证 为了评估差异代谢物对休眠孢子萌发的影响,我们选择了一些代谢物,如L-isoleucine, Amentoflavone, 2′-Deoxyuridine, Trigonelline, Thymine, Coumarin, Pyridoxal 5′-phosphate, D-lyxose,参照根系分泌物的方案测试不同次级代谢物刺激休眠孢子萌发的能力. 同时选取萌发率差异较大的两种代谢物处理休眠孢子,进行根肿病表型实验. 两个试验中使用的这些代谢物的浓度分别为2 mg/L、5 mg/L、5 mg/L、100 nmol/L、5 mg/L、3 mg/L、5 μmol/L、5 mg/L和3 mg/L,每个处理重复3次.
3 结果与分析
3.1 不同抗性材料根系分泌物诱导休眠孢子萌发
不同抗性材料的根系分泌物刺激休眠孢子萌发的效果有明显差异. 在处理开始时(第0 d),所有处理中约有0.97%的孢子在萌发,各处理之间没有差异. 处理1 d后,R-197根系分泌物处理的孢子萌发率显著高于华双5R和Kc84-1根系分泌物处理及对照组(P≤ 0.05). 在第2~7 d,R-197根系渗出物刺激休眠孢子萌发的效率总是显著高于华双5R和Kc84-1根系分泌物处理及对照组(P≤ 0.05)(图1b). 这些结果表明,与抗性材料相比,易感材料的根系分泌物中或许存在一些特有的或共有但含量较高的物质导致了休眠孢子的大量萌发.
王俊[3]等报道中良性结节血流分级主要为0~Ⅰ级,恶性结节血流分级主要为Ⅱ~Ⅲ级;相较于恶性结节,良性结节的血流信号丰富比率、RI和钙化比率均更低,形态规则比率、边界清晰比和晕环比率均更高,差异具有统计学意义(P<0.05)。本次研究的诊断结果和其他报道基本一致,可行性高[4]。
3.2 代谢物的鉴定
从R-197、华双5R和Kc84-1的根系分泌物中共鉴定出1257(ESI+)和1079(ESI-)种次级代谢物(用飞行时间和质荷比(RT-m/z)表示). 为了区分三个样品组,将ESI+(ESI-)中的1,257(1079)个代谢物进行PCA分析(图2a, d),三组样品除R-197中的两个样品外都可明显分开. 在所有的变量中,ESI+中61.38%(ESI-中56.23%)的模式与第一成分有关,ESI+中9.28%(ESI-中13.43%)与第二成分有关,区分了A、B和C(A、B、C分别代表华双5R、Kc84-1和R-197). 为了可靠的组间差异和后续的可视化分析,采取了OPLS-DA,得分图显示三个样本组之间有明显的区别(图2b, 2c, 2e, 2f).
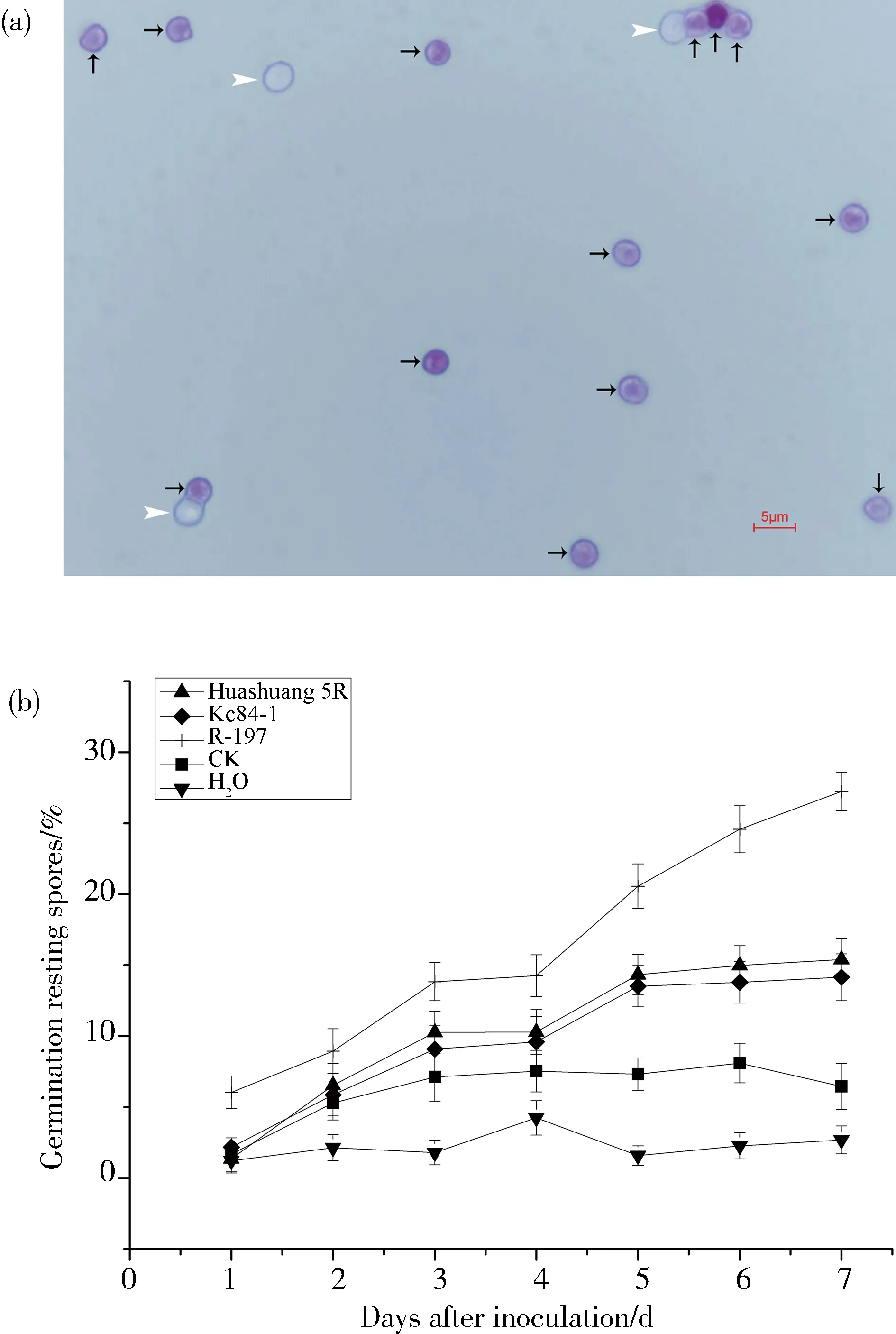
图1 根系分泌物对芸薹属根肿菌(P. brassicae)休眠孢子萌发的影响(a)水中培养1 d后,在显微镜下观察到的休眠孢子,黑色箭头和白色箭头分别表示未萌发和萌发的孢子; (b)不同抗性材料的根系分泌物处理组和对照组中休眠孢子的萌发情况,误差棒表示标准差Fig.1 Effect of root exudates on the germination of P. brassicae resting spores(a) Resting spores observed in the microscope after 1 day of incubation in water, black and white arrows indicate non-germinating and germinating spores, respectively; (b) germination of resting spores in the treated and control groups with different resistant materials root exudates, bars= SDs
3.3 差异性代谢物的筛选
为了寻找在刺激休眠孢子萌发中起重要作用的关键代谢物,分别对易感材料R-197与抗性材料华双5R和Kc84-1进行了代谢物的分组比较. 在ESI+模式下,与华双5R相比,R-197有120个代谢物含量上调,250个代谢物下调;与Kc84-1相比,中R-197有337个代谢物上调,180个代谢物下调. 在ESI-模式下,与华双5R相比,R-197中96个代谢物上调,243个代谢物下调;与Kc84-1相比,R-197中353个代谢物上调,142个代谢物下调(图3). 差异显著的次级代谢物如表1所示.
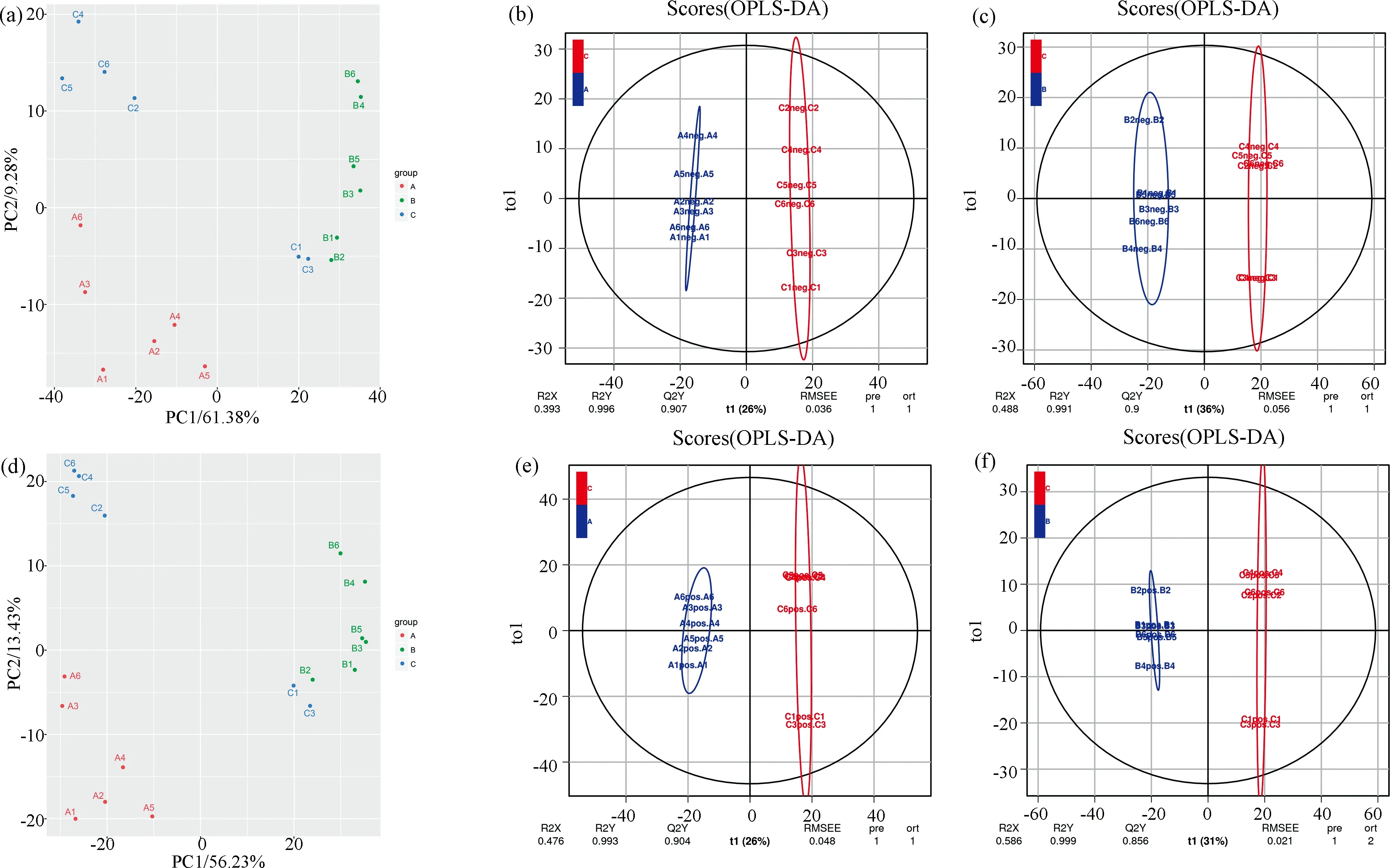
图2 主成分分析(PCA)及正交偏最小二乘法-判别分析(OPLS-DA)
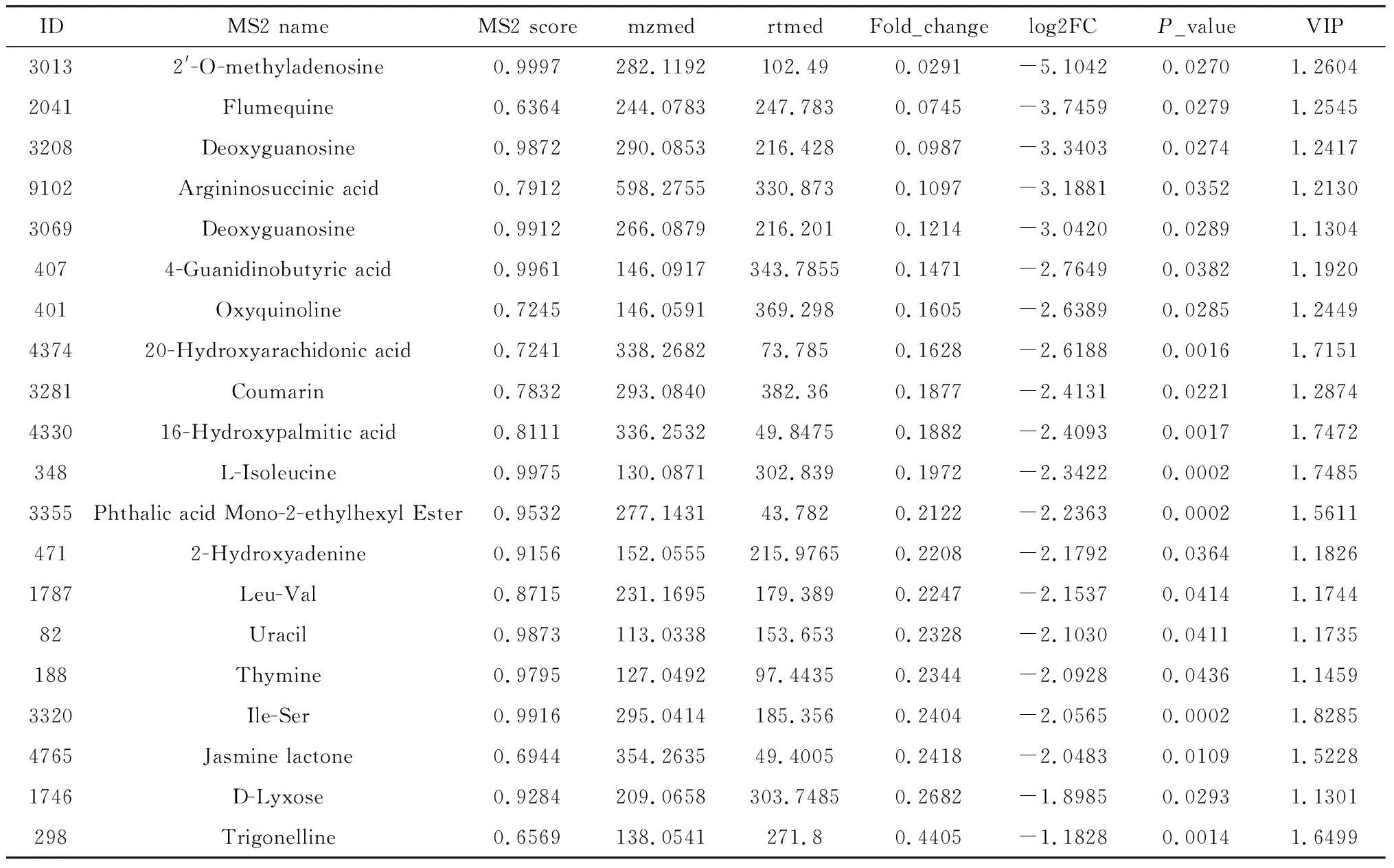
表1 部分差异代谢物
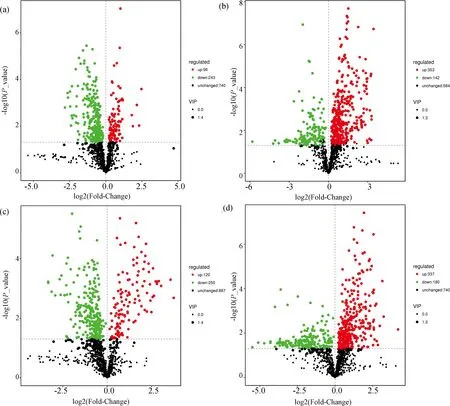
图3 差异代谢产物火山图(Volcano plot)
3.4 差异代谢物功能验证
为了验证差异代谢物在刺激休眠孢子萌发方面的作用,我们选择了一些差异代谢物单独进行萌发试验. 筛选原则如下:易感材料R-197与抗性材料华双5R和Kc84-1之间存在许多差异性代谢物. 易感材料分别与两种抗性材料进行比较得到两个差异组,而这两个组的差异代谢物分析表明,既有相同的,也有不同的,因此用于验证的差异代谢物涵盖了这两类. 在萌发实验中,孢子的初始萌发率为0.11%. 孵育前2 d,所有处理之间无明显差异. 处理第4 d,L-Ile(L-isoleucine)和Trigonelline处理组的萌发率显著升高(P≤ 0.05)(图4). 处理第6 d,用不同的差异性代谢物处理的休眠孢子的萌发率差异更为明显(P≤ 0.05). 除Arg组外,其他处理组的休眠孢子萌发率都与对照组均有显著差异,且不同处理组间也有很大差异,这些结果说明Thymine,2′-Deoxyuridine,L-isoleucine,Trigonelline,Pyridoxal 5′-phosphate,Coumarin,D-lyxose和Amentoflavon等差异代谢物可以刺激休眠孢子的萌发.
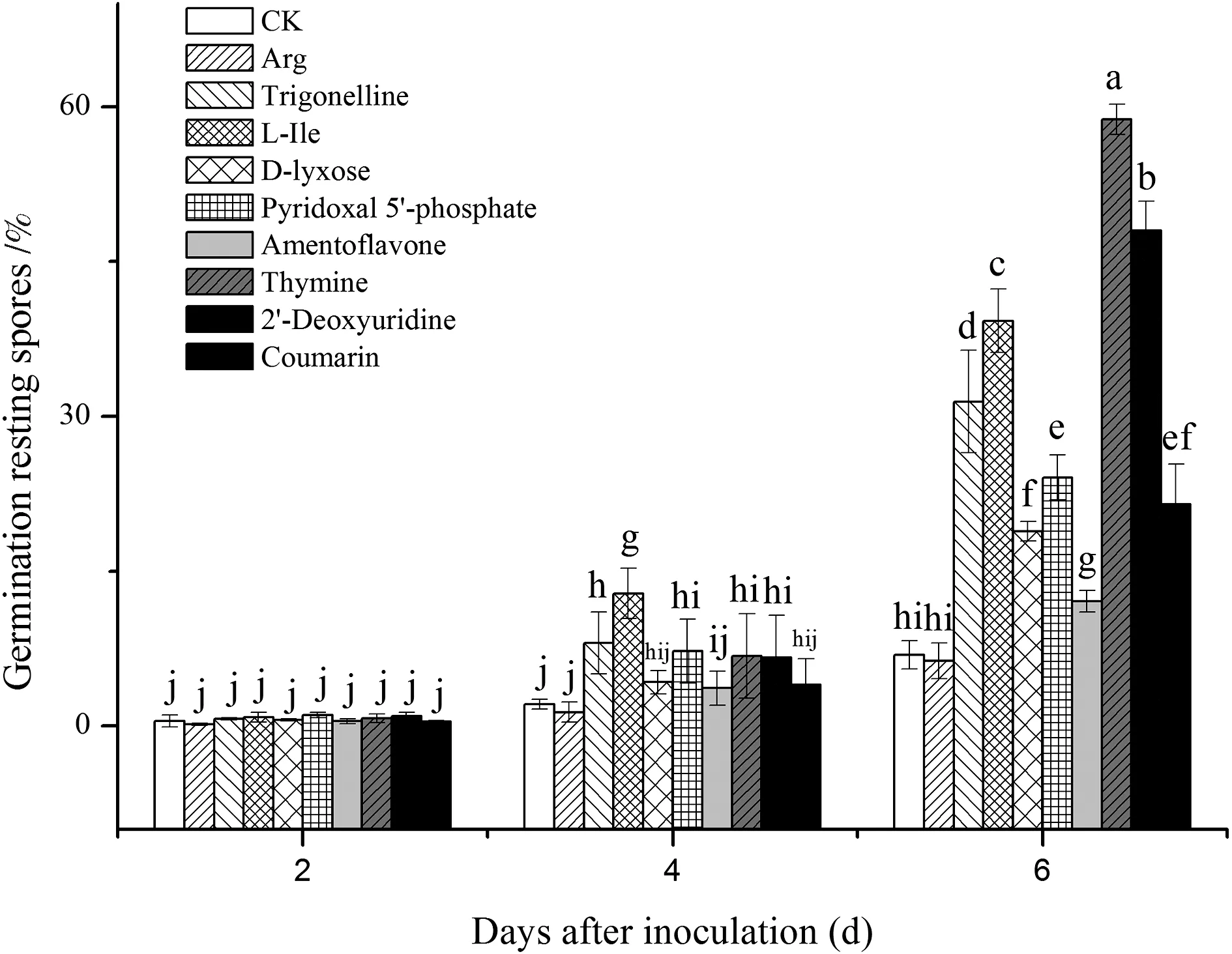
图4 差异代谢物处理组和对照组中休眠孢子的萌发情况误差棒表示标准差,显著性使用单因素方差分析,图基检验,P≤ 0.05,字母不同表示具有显著性差异Fig.4 Germination of resting spores in the differential metabolite treatment and control groupsBars= SDs, significance was taken with one-way ANOVA, Tukey′s test, P≤ 0.05, different letters indicate significant difference
3.5 休眠孢子萌发率对根肿病发病的影响
为了进一步确认不同萌发率的孢子对根肿病发病的影响,我们分别选择Arg和Thymine孵育到7 d时的休眠孢子悬液(萌发率分别是0.13%和71.2%)对抗性材料华双5R接种,30 d后调查发病情况. 结果表明,接种低萌发率孢子悬浮液的华双5R表现出较强的抗病特征,感病植株较少,且感病程度很低. 但使用高萌发率的孢子悬浮液接种,发病率达到了100%,且发病程度也很严重(图5a, 5b). 因此,我们推断,休眠孢子的萌发在根肿病发病过程中起重要作用:高萌发率导致高发病率. 另外,我们将易感材料R-197和抗性材料华双5R种植在一起,并保证根系相互接触,对照组则单独种植. 30 d后调查了发病率. 结果表明,与对照组相比,捆绑种植的抗性材料发病率显著增加. 这说明易感材料可以分泌出大量引起休眠孢子萌发的代谢物,致使根系周围的休眠孢子萌发为具有极强侵染能力的初级游动孢子进而提高了根肿病的发病率和患病程度(图5c).

图5 休眠孢子萌发率对抗性材料抗病表型的影响
4 讨 论
植物的根部可以合成、积累和分泌各种各样的化合物,如氨基酸、有机酸、多糖和蛋白质,这些化合物有的可以分泌到根系周围,被称为根系分泌物[18]. 遗传背景是影响根系分泌物含量和种类的一个重要因素. 在我们的研究中,通过鉴定,并分别比较了两个种不同的抗根肿病材料与易感根肿病材料的根系分泌物中的代谢物组分寻找可以刺激休眠孢子萌发的代谢物,如氨基酸、核苷、有机酸和激素等,这些代谢物也被称为萌发刺激因子(GSFs)[19]. 此外,在比较两组差异代谢物时,我们发现与易感材料相比,不同抗性水平的代谢物中存在一些特异的差异代谢物和一些相同的差异代谢物. 对于这些相同的差异性代谢物,两组的差异倍数是不同的. 我们猜想,这些相同的差异代谢物更有可能是关键的萌发刺激因素,根肿菌孢子萌发验证结果也证实了这一点. L-isoleucine, Amentoflavone, 2′-Deoxyuridine, Trigonelline和Thymine以及一些其他含量差异较大代谢物的作用或多或少都与微生物孢子或萌发作用有关. 茉莉酸(JA)似乎被一些真菌病原体利用作为效应物[20]. 2′-Deoxyuridine被确定为能够促进稻瘟病真菌分生孢子侵染的活性分子[21].Trigonelline可能是微生物-植物共生或致病系统中的信号分子,在许多豆科植物根部分生组织的早期生长中起细胞周期调节器的作用[22,23]. 根瘤菌可以利用由其宿主Medicagosativa[24]合成的Trigonelline作为渗透保护剂和能量来源.Thymine是生物的一个重要化学组分,可以促进Scsbamaaculeata的结节,但高浓度的Thymine具有杀菌作用[25,26]. 香豆素化合物是植物合成的重要次级代谢物,它具有许多重要的生物功能,首先具有抗菌活性,其次,它也可以作为共生细菌、病原体和其他植物的信号分子[27].
在本研究中,与易感材料一起种植,抗性材料也普遍感染根肿病,在一定条件下,抗性材料的发病甚至可以达到与易感材料相当的水平. 目前进行的抗性研究,大多是利用自然条件下筛选出的抗性材料,建立定位群体,定位主导抗病的抗性位点,而关于深层抗病机理的报道却很少. 另一方面,对于筛选出的抗性材料,不同实验室有不同甚至相反的结果. 原因很复杂,比如根肿菌繁多的生理小种、不同的接种方法、不同的培养条件等. 在我们的实验中,将病原体、接种方法和培养环境等因素控制在相同,结果表明不同萌发率根肿菌孢子处理抗病材料,植株抗病性存在显著差异. 由此推断,根肿病的发生与休眠孢子的萌发率密切相关. 一些研究发现,根肿病的发生也部分取决于土壤环境中休眠孢子的含量,也就是所谓的有效浓度,即只有根际的萌发孢子参与感染,易感材料的根可以分泌大量的物质,触发根肿菌休眠孢子的萌发,并导致有效孢子的含量增加,最终导致植株产生根肿. 而抗性材料的根系分泌物中这些物质的含量较低甚至没有,即使在休眠孢子含量极高的土壤中,由于萌发的有效孢子的含量较低,这些植株也能免于被侵染,表现出抗性. 当然,根肿病的抗性机制是多种多样的,我们的研究也只是提供一个新的视角.
本研究首次采用UPLC-MS方法对根肿病抗性和易感材料植株的根系分泌物进行分析,筛选出一些差异代谢物,并确认一些代谢物可以有效刺激休眠孢子萌发. 这些差异代谢物在很多方面都可能存在潜在的应用前景. 例如,利用根系分泌物可开发出稳定的化学药剂,刺激休眠孢子的萌发,对带菌田块进行预处理,防治根肿病;另一方面,一些差异代谢物也可以开发成抗性育种的生物标志物. 植物根际是一个复杂的环境,不同代谢物在体外的稳定性也是一个重要的影响因素,不同代谢物的性质及作用验证仍需要进一步研究.
