文山国家级自然保护区不同海拔地带性植被的土壤微生物生物量碳氮分布特征*
2022-06-15吕晶花杨志东任玉连
李 聪 吕晶花 陆 梅 杨志东 刘 攀 任玉连 杜 凡
(1.西南林业大学生态与环境学院 昆明 650224; 2.西南林业大学林学院 昆明 650224; 3.贵州大学生命科学学院 贵阳 550025)
土壤微生物生物量碳氮是土壤碳氮库中较为活跃的生物组分(王宁等, 2016),直接驱动土壤微生物参与的碳氮养分循环过程,并能调控土壤肥力形成、植物生长及群落演替(王慧颖等, 2018)。土壤微生物生物量碳氮含量对植被群落和土壤环境变化十分敏感(Xuetal., 2017; 涂志华等, 2018)。李文周等(2019)与张义凡等(2017)研究表明,土壤微生物生物量碳氮含量随海拔变化呈单峰型变化趋势,并与土壤养分含量变化显著正相关(P<0.05)。田琴等(2017)研究发现土壤微生物生物量碳氮不仅与土壤理化性质紧密相关,且受植被类型的显著影响(P<0.05)。马和平等(2013)研究发现,不同海拔水热条件的空间异质性可能会改变植被群落的垂直地带分布,引起土壤理化环境差异,进而导致土壤微生物生物量碳氮呈现不同的海拔分布格局。前人对不同海拔上典型植被的土壤微生物生物量碳氮分布开展了一定研究,一些学者研究发现,随着海拔上升,植被群落呈现出典型的地带性分布,土壤微生物生物量碳氮的含量沿海拔上升呈增加趋势(丛静等, 2014; 王琴等, 2013),而另一些学者研究结果则与之相反,在低海拔植被带向高海拔植被带的垂直演替过程中,土壤微生物生物量碳氮含量沿海拔上升呈减小趋势(赵盼盼等, 2019; Zhouetal., 2015)。上述研究表明土壤微生物生物量碳氮可能会因生境差异而对海拔上不同地带性植被群落产生不同的响应,因此,研究土壤微生物生物量碳氮沿不同海拔地带性植被的变化特征,有助于理解环境因子变化对森林土壤微生物生物量碳氮含量及土壤碳氮循环过程的调控机制,可为科学管理和经营森林提供参考。部分学者对土壤微生物生物量碳氮含量与不同海拔土壤因子间的相互关系开展了一些定性探讨,如赛牙热木·哈力甫等(2017)研究发现,微生物生物量碳氮含量与不同海拔下土壤密度、有机质含量和碱解氮含量显著正相关(P<0.05),与pH值、全氮含量和碳氮比显著负相关(P<0.05)。曹瑞等(2016)研究发现土壤微生物生物量碳氮含量与土壤酶活性极显著正相关(P<0.05)。但这些研究仍未能很好地回答调控不同地带性植被群落土壤微生物生物量碳氮含量时空变化的关键影响因子是什么。
云南文山国家级自然保护区地处南亚热带典型季风气候区,森林植被群落以典型的亚热带常绿阔叶林为主,沿海拔自下而上形成了亚热带季风常绿阔叶林、半湿润常绿阔叶林和中山湿性常绿阔叶林地带植被分布景观(杨宇明等, 2008),这3种植被在多样性、生态外貌和土壤类型上差异明显(Zhuetal., 2019),是研究亚热带典型地带性土壤微生物生物量碳氮的理想场地。目前,在该区域已经开展了不同海拔森林植物和动物群落的研究(王革, 2016; 黄乐乐, 2015; 胡彦等, 2004),但尚未开展不同地带性植被土壤碳氮养分循环特征的研究,土壤微生物生物量碳氮沿不同海拔地带性植被的分布特征及其驱动因子有待进一步研究。本研究以保护区内3个典型地带性植被类型为研究对象,探究文山国家级自然保护区土壤微生物生物量碳氮含量沿海拔和土层的分布特征及其关键调控因子,以期为理解亚热带森林土壤碳氮循环过程及其调控机制提供基础数据参考。
1 研究区概况
文山国家级自然保护区(104°41′—104°52′E,23°21′—23°24′N)地处滇东南部低纬度高原,地形以山原地貌和岩溶地貌为主; 气候类型属南亚热带季风湿润型,因受地理位置、地形、海拔等影响,具有明显的立体气候特征,山下部年降雨量1 000~1 400 mm,山上部年降雨量大于1 400 mm。山上部年均气温低至7~8 ℃,山下部则可达20 ℃以上(杨宇明等, 2008); 植被多样性沿海拔上升而减小(杨宇明等, 2008)。
2 研究方法
2.1 样地设置与植被调查
2018年10月,选取文山国家级自然保护区3种不同海拔的地带性植被,即低海拔亚热带季风常绿阔叶林(L)、中海拔半湿润常绿阔叶林(M)和高海拔中山湿性常绿阔叶林(H),参照《植物学实验方法》(章英才等, 2007)在每个植被带内随机布置3块30 m×30 m样地,同一海拔的3块样地间距离均大于10 m,不同海拔植被带样地间距离均大于200 m,样地基本情况见表1。
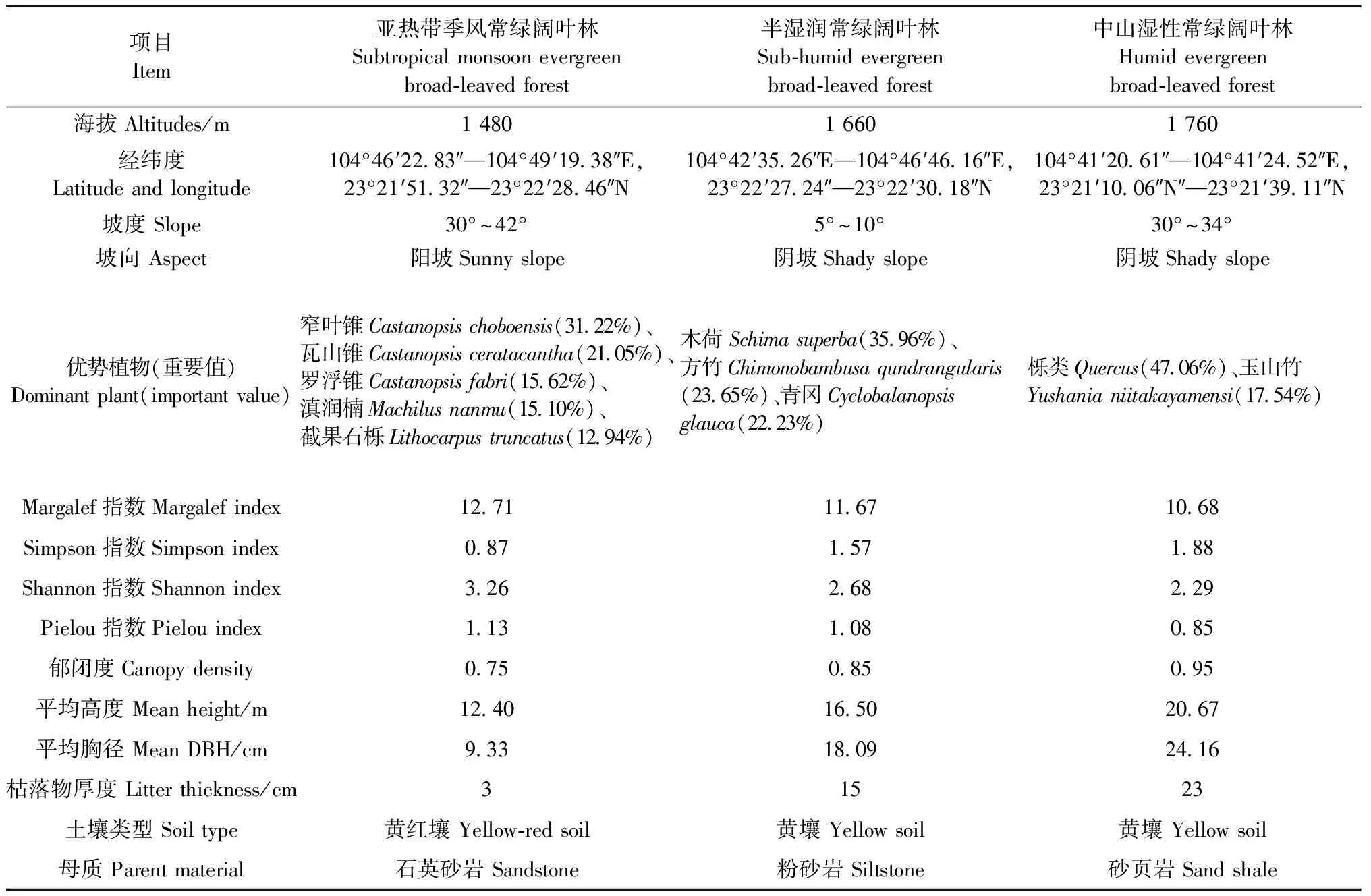
表1 样地基本情况Tab.1 Basic situation of the sample sites
将每块30 m×30 m样地采用“网格法”划分为36个5 m×5 m样方,记录每个样方内乔灌层植物(胸径≥5 cm)的名称、盖度、高度、胸径和优势种,并根据调查结果计算植物群落的多样性指数。
2.2 群落多样性指数计算
植物群落α多样性指数包括:
Margalef指数(R):R=(S-1)/lnN;
Pielou指数(Jsw):Jsw=D/lnS。
式中: S为调查样地物种数;Pi为i物种个体数占总物种个体数的比率,Pi=Ni/N,Ni为某物种个体数,N为所有物种的个体数之和(陈金磊等, 2019)。
2.3 土样采集及土壤理化指标测定
在每块样地中,沿对角线设置3个1 m×1 m采样点,先测量采样点内枯落物厚度,然后去除枯落物挖掘土壤剖面,按0~10 、10~20、20~30、30~40和40~50 cm分5层从上到下采集土样,每土层采集土样2.5 kg,清除土样中的植物根系、枯落物等杂物后,采取四分法将每块样地内3个取样点土样按照上述分层混合成5份,放入无菌土壤袋中带回实验室,共计45个土样。将每个土样分成3份: 一份鲜土(200 g)用于测定土壤自然含水量; 一份鲜土(1 000 g)于室内自然风干、磨细过筛,用于测定土壤理化性质; 一份鲜土(1 000 g)过2 mm筛后冷藏于冰箱中,于一周内测定土壤微生物生物量碳氮含量。
土壤温度采用地温计测定; 土壤含水量采用烘干-恒质量法测定; 土壤密度采用环刀法测定; pH值采用水浸提-玻璃电位法测定; 有机质含量采用重铬酸钾氧化-油浴外源加热法测定; 全氮含量采用半微量-凯氏定氮法测定; 全磷含量采用镍坩锅碱融-钼锑抗比色法测定; 全钾含量采用镍坩锅碱融-火焰光度计法(鲁如坤, 2000)测定。每个样品至少测定3个重复,取其均值。
土壤微生物生物量碳含量采用氯仿熏蒸-K2SO4浸提-重铬酸钾外源加热法获得; 土壤微生物生物量氮含量采用氯仿熏蒸-K2SO4浸提-凯氏定氮法获得(吴金水等, 2006)。每个土样称取6份5 g鲜土,其中3份加入三氯甲烷(别名:氯仿,CHCl3),使用真空泵进行熏蒸处理(CH),另外3份不加氯仿作为对照(CK),分别取熏蒸和对照的有机碳氮和全氮含量均值。根据下列公式计算第i层土壤微生物生物量碳含量(SMC)和微生物生物量氮含量(SMN)(mg·kg-1)(吴金水等, 2006):
SMC=Ei(C)T-Ei(C)CK;
SMN=Ei(N)T-Ei(N)CK。
式中:Ei(C)T和Ei(N)T分别为第i层熏蒸处理的土壤有机碳和土壤全氮含量(mg·kg-1),Ei(C)CK和Ei(N)CK分别为第i层未熏蒸处理的土壤有机碳和土壤全氮含量(mg·kg-1)。
2.4 数据处理
采用SPSS25.0单因素方差分析(One-way Anova)探讨土壤理化指标的差异,差异显著性水平设置为α=0.05,共线性用方差膨胀系数(VIF)表示,若VIF>10,意味着变量之间存在共线性,且值越大共线性越强。Partial Mantel(偏曼特尔)基于曼特尔对变量相关性的评估和检验,计算变量间相异矩阵与样地距离等级矩阵,运用该方法可以排除变量矩阵的多重关联及交互作用,找到最佳关联因子(覃光莲等, 2014); Fourth-Corner方法则用于检验单个因子和单个环境变量的具体关系(王琇瑜等, 2020)。Partial Mantel和Fourth-Corner两种方法具有互补性,常组合起来分析变量子集对环境变化的响应。本研究采用曼特尔偏相关检验土壤理化因子和微生物生物量碳氮含量的相关性,循环1 000次排列,去除共线性,根据相关系数r和显著性P值(r值越大相关性越大,P<0.05表示统计具有显著性),找到关键土壤因子; 再结合植被多样性和枯落物厚度的数据,采用Fourth-Corner方法检验各植被带中环境变量与土壤微生物生物量碳氮含量关系,模拟999次,检测方案为双向检测,选择微生物生物量碳氮矩阵的观测值、标准差及P值反映二者的检验关系,明确土壤微生物生物量碳氮含量在各植被带的主控因子,采用Orign(2021)软件绘图。
3 结果与分析
3.1 不同地带性植被土壤微生物生物量碳氮含量变化
不同地带性植被样地的土壤SMC和SMN均存在显著差异(图1,P<0.05)。由图1可知,SMC和SMN含量均表现为亚热带季风常绿阔叶林(15.75和2.84 mg·kg-1)<半湿润常绿阔叶林(28.05和4.95 mg·kg-1)<中山湿性常绿阔叶林(41.61和7.80 mg·kg-1),均呈现从低海拔亚热带季风常绿阔叶林至高海拔中山湿性常绿阔叶林显著增加的趋势。
方差分析(图2)表明,同一植被带SMC和SMN含量随土层加深而递减的趋势显著(P<0.05)。0~10 cm土层的SMC和SMN含量显著大于40~50 cm土层(P<0.05),且两土层间差异在植被带间表现为: 亚热带季风常绿阔叶林(7.21、3.42倍)>中山湿性常绿阔叶林(2.85、3.28倍)>半湿润常绿阔叶林(2.73、2.95倍)。从0~10 cm土层到40~50 cm土层,SMC含量分别为35.78~68.93、17.91~52.71、12.84~38.64、7.27~30.89 和4.96~22.54 mg·kg-1,SMN含量分别为5.07~12.99、3.32~9.71、2.44~6.06、1.87~5.29和41.48~3.96 mg·kg-1。同一土层的SMC和SMN含量沿海拔升高增加趋势显著(P<0.05)(图2)。
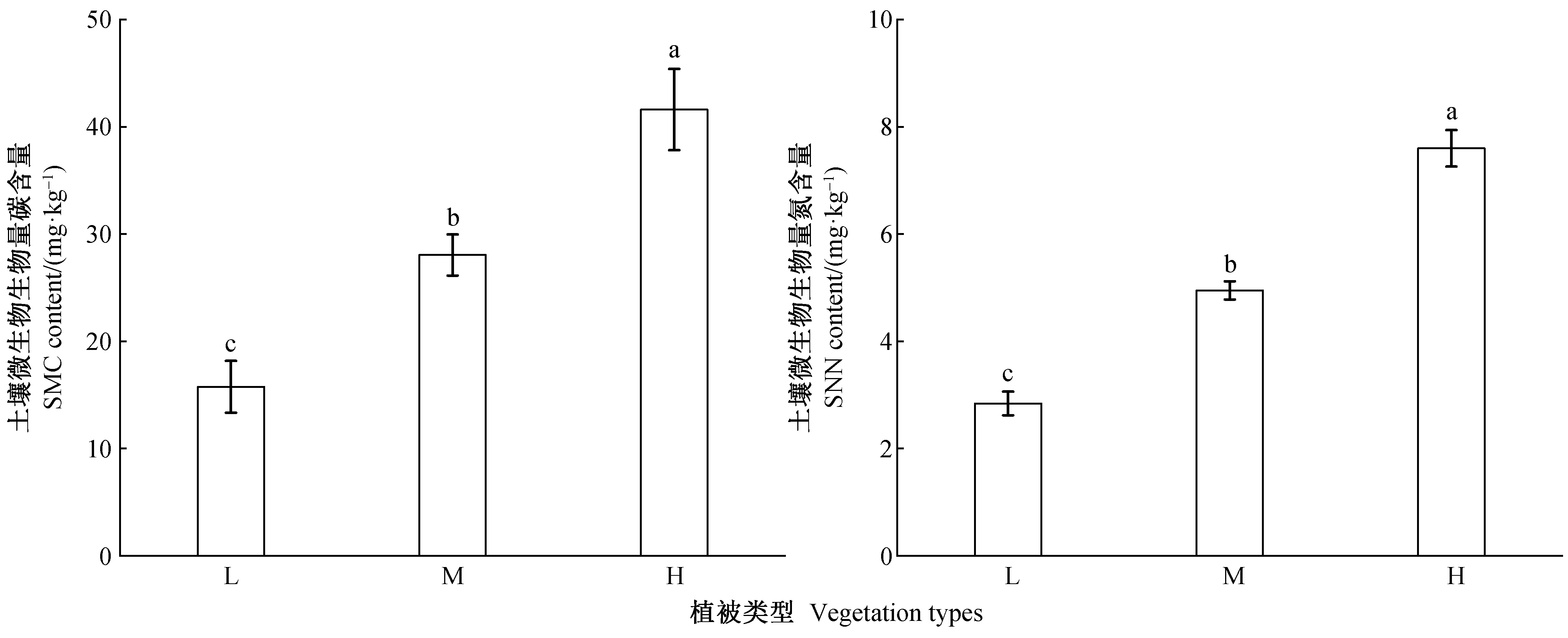
图1 不同植被带土壤微生物生物量碳氮含量Fig. 1 Contents of soil microbial biomass carbon and nitrogen in various vegetation zones植被带间的不同小写字母表示差异显著(P<0.05),最大的平均数标记为a。Different low-case letters indicate a significant differences (P<0.05) among vegetation zones, the maximun mean value mark as a.

图2 不同植被带土壤微生物生物量碳氮土层变化特征Fig. 2 Profile changes of soil microbial biomass carbon and nitrogen in the three vegetation zones不同小写字母代表同一土层各植被带间的微生物生物量碳氮含量在0.05水平上差异显著,不同大写字母表示同一植被带各土层的微生物生物量碳氮含量在0.05水平上差异显著。Different lower-cases indicate significant in microbial biomass C and N content among the three zonal vegetations in the same layer (P<0.05) , the different upper-cases indicate significant in microbial biomass C and N content among the five soil layers in the same zonal vegetation (P<0.05).
3.2 土壤因子随海拔和土层的变化特征
由表2可知,3个地带性植被的土壤环境存在较大的差异。低海拔植被的土壤密度、温度和pH值均显著高于中海拔和高海拔(P<0.05),分别为中海拔的1.23、1.45和1.14倍及高海拔的1.47、2.48和1.23倍; 土壤全磷含量随海拔升高呈先减后增的趋势,高海拔为中海拔的3.32倍,为低海拔的1.66倍; 高海拔的土壤含水量、有机质含量、全氮含量和全钾含量显著高于中、低海拔两个植被带(P<0.05),分别为中海拔的1.22、1.50、1.55和1.24倍及低海拔的的1.53、2.61、2.70和2.26倍。
土壤各环境因子沿土层加深呈明显垂直分异(表2): 土壤含水量、有机质、全氮、全磷、全钾含量呈减小趋势(P<0.05); 土壤密度、温度和pH值则呈增加趋势(P<0.05)。

表2 不同地带性植被各土层土壤因子①Tab.2 Soil environment factors of various soil layers under three zonal vegetations
3.3 土壤理化因子与微生物生物量碳氮含量的偏Mantel检验
为减小各土壤理化因子对微生物生物量碳氮含量的共线性影响(VIF>10,均值为60.82),采用偏Mantel检验方法对不同地带性植被土壤微生物生物量碳氮含量和土壤理化因子之间的关系进行1 000次排列检验,以得到微生物生物量碳氮含量的关键耦合因子(表3)。结果表明,土壤有机质含量、全氮含量、pH值和微生物生物量碳氮的偏Mantel检验的相关系数r≥0.75,且P<0.01,表明上述因子与微生物生物量碳氮含量极显著相关,是与微生物生物量碳氮含量沿植被地带性变化耦合的关键因子(表3); 而含水量、密度、全钾含量与微生物生物量碳氮含量的相关性仅次于土壤有机质、全氮含量和pH(0.5
3.4 不同植被类型微生物生物量碳氮含量对关键环境因子的响应
根据表3分析结果,选取土壤理化关键因子(土壤有机质含量、全氮含量、pH)、植被多样性及枯落物厚度作为关键环境因子,采用第四角度(Fourth-Corner)分析上述因子与微生物生物量碳氮含量矩阵在各海拔植被带的耦合关系。表4显示:亚热带季风常绿阔叶林的土壤微生物生物量碳氮与植被多样性(Shannon指数,Margalef指数,Pielou指数)极显著负相关(P<0.01),而与土壤理化关键因子(表3,土壤有机质含量、全氮含量、pH)的关系不显著; 半湿润常绿阔叶林和中山湿性常绿阔叶林中,土壤微生物生物量碳氮含量与pH值极显著负相关(P<0.01),与枯落物厚度、有机质含量和全氮含量极显著正相关(P<0.01),半湿润常绿阔叶林土壤微生物生物量碳氮含量与植物多样性中的Shannon指数和Margalef指数显著负相关(P<0.01); 中山湿性常绿阔叶林土壤微生物生物量碳氮含量只与植物多样性中的Simpson指数极显著正相关(P<0.01),这表明高海拔植被带土壤微生物生物量碳氮含量对植被多样性的响应越来越小,且主要调控因子为枯落物厚度和土壤理化因子。

表4 不同海拔地带性植被环境因子与微生物生物量碳氮含量的Fourth-Corner分析①Tab.4 Four-Corner analysis for the association of environmental factors with microbial biomass carbon and nitrogen content in zonal vegetation among different altitudes
4 讨论
4.1 森林土壤微生物生物量碳氮的海拔与土层变化
土壤微生物生物量碳氮作为生态系统最具活性的碳和氮组分,能够敏感响应森林土壤环境的海拔与土层变化(李品等, 2019; Ashrafetal., 2020)。本研究中,土壤微生物生物量碳氮含量沿海拔升高表现出显著的增加趋势(P<0.05)。Fourth-Corner分析表明,土壤微生物生物量碳氮含量沿地带性植被带变化是枯落物厚度、植物群落多样性和关键土壤理化因子共同作用的结果。一方面,低海拔亚热带季风常绿阔叶林的群落组成多为矮乔灌,虽然多样性指数较高,但林间空隙较大,植被郁闭度较低,太阳可直射地表,导致土壤温度升高,水分蒸发旺盛,加快了微生物对有机物的分解速率,抑制了微生物生物量碳氮的积累(姚兰等, 2019)。另一方面,随着海拔上升,植被演替从低海拔的矮乔灌群落演变至高海拔的乔木群落,增加了地表枯落物,伴随着土壤水分增加和pH值减小,为土壤微生物生物量碳氮的积累提供了良好的环境(田琴等, 2017)。
各地带性植被土壤微生物生物量碳氮含量随着土层加深呈显著减小的变化(P<0.05),与Ravindran等(2015)和柳杨等(2017)的研究结果相似。枯落物主要富集在表层,为表层微生物提供了充足养分来源(王传杰等, 2018),促进了土壤微生物的生长发育及分解活动。而随着土层加深,土壤水、气、热等条件逐渐恶化,导致微生物向土壤中转移养分的速率下降(段春燕等, 2020),不利于土壤微生物生物量碳氮在深层土壤的积累和储存。研究还发现,不同植被带微生物生物量碳氮含量沿土层的降幅不一,低海拔植被带土壤微生物生物量碳氮的降幅显著高于中高海拔植被带。这与土壤结构有关,低海拔亚热带季风常绿阔叶林土壤密度较高,下层土壤板实,严重削弱了土层水养的运输能力,导致下层土壤微生物活性因缺乏养分而减弱,而近地表层覆盖着枯落物,微生物可获得的有机质最多,因此该海拔植被土壤微生物生物量碳氮含量逐层递减趋势最显著(王宁等, 2015); 半湿润常绿阔叶林处于中海拔地带,土壤水分和养分增多改善了土壤结构,促进了枯落物和养分在该植被带的积累,枯落物庇护和林冠截留使得降雨对土层间养分的淋溶作用较小,且缓坡地形(表1)不易造成底层土壤微生物生物量碳氮的流失,故而该植被带变幅较小; 中山湿性常绿阔叶林系保护区降水较多的林区,且坡度较陡,易造成底层土壤养分子林溶和流失但该地土壤结构较其他2个植被带优,上下层均为质地较松的壤土,土壤胶体的储养能力最强,能有效缓解因地形和降雨造成的土壤微生物生物量碳氮流失过多的问题,故而降幅居于中等。因此,微生物生物量碳氮含量的土层变化与植被带所处的地形、降雨量和土壤结构有关。
4.2 森林地带性植被类型对土壤微生物生物量碳氮的影响
在森林生态系统中,地上植被能够通过凋落物和多样性变化影响土壤碳氮养分循环过程,并对微生物生物量碳氮积累产生影响(Kiranetal., 2018)。先前研究发现,较高的植被多样性可以促进土壤微生物生物量碳氮含量(李鑫等, 2014),但在本研究中,植被多样性与土壤微生物生物量碳氮含量显著负相关(P<0.05),且沿海拔上升呈减弱趋势,这可能是保护区沿海拔植被群落差异造成的。随着海拔升高,“温减水增”的林区微气候变化限制了中高海拔地区一些喜热物种的出现,使植被多样性沿海拔上升呈减小趋势,同时降雨量的增加引起植被群落“由灌到乔”地带性变化,使地表凋落物量沿海拔升高呈递增变化(任玉连等, 2019),不仅增加了高海拔土壤微生物的底物获取量,还削弱了植被多样性减小对微生物生物量碳氮积累的不利影响。
4.3 关键土壤理化因子对微生物生物量碳氮的影响
不同地带性植被的土壤理化性质存在一定的差异(任玉连等, 2020),从而对土壤微生物生物量碳氮含量产生影响。本研究发现,土壤有机质含量、全氮含量和pH值是调节土壤微生物生物量碳氮含量变化的主要理化因子。王云霞等(2018)研究表明,土壤碳氮含量一定程度上反映了土壤环境因子组合的最佳程度,高水平的土壤有机质可以给微生物代谢提供足够能量。偏Mantel检验表明,土壤有机质和全氮含量与微生物生物量碳氮含量极显著正相关(P<0.01),这与全飞等(2019)和贾国梅等(2016)的研究结果类似,表明碳氮的丰缺会引起微生物生物量碳氮的变化。
pH值可调节土壤微生物的活性,进而影响微生物生物量碳氮变化和分布(梁国华, 2015)。本研究发现,土壤pH值与微生物生物量碳氮含量显著负相关,说明较高海拔植被带的土壤pH值较低,有利于土壤微生物生物量碳氮的积累。
本研究还发现,土壤全磷含量未对土壤微生物生物量碳氮含量产生显著影响,这与李艳琼等(2018)全磷含量与微生物生物量碳氮含量极显著正相关的研究结果不一致。这可能与文山保护区土壤发育类型和磷素含量有关。文山保护区地处南亚热带气候区,受降雨淋溶作用,酸性较强,土壤以红壤、黄壤为主,这类土壤富含氧化铝、氧化铁等矿物,易形成难溶性磷,且各海拔植被带全磷含量较低(1.29 g·kg-1),与土壤中其他含量较多的碳氮养分相比,量少难矿化的磷素不是微生物底物利用的第一选择,因此,磷素与微生物生物量碳氮的耦合关系较弱。
4 结论
随着海拔升高,文山自然保护区土壤微生物生物量碳氮含量呈增加趋势; 土壤微生物生物量碳氮含量随土层加深呈减小的趋势。
偏Mantel’s test相关性检验表明,土壤有机质含量、全氮含量和pH值是微生物生物量碳氮含量沿植被地带性变化的关键耦合因子。 第四角度(Fourth-Corner)分析结果表明:低海拔的亚热带季风常绿阔叶林土壤微生物生物量碳氮的主要调控因子为植被多样性(Shannon指数,Margalef指数,Pielou指数),且二者间呈极显著负相关性(P<0.01);而在中海拔的半湿润常绿阔叶林和高海拔的中山湿性常绿阔叶林中,土壤微生物生物量碳氮对植被多样性的响应随海拔升高而减弱,主控因子转变为枯落物厚度、pH值、有机质含量和全氮含量。
