上海市水稻田千金子对3 种乙酰辅酶A 羧化酶抑制剂的抗性现状及酶突变机制
2022-06-14袁国徽田志慧沈国辉
袁国徽, 田志慧, 高 原, 沈国辉
(上海市农业科学院 生态环境保护研究所,上海 201403)
千金子Leptochloa chinensis(L.) Nees 属一年生禾本科杂草,是一种世界性农田恶性杂草,在我国主要分布于华东、华中、华南、西南及陕西省等地[1-2]。由于水稻栽培制度的变化和除草剂长期单一使用,千金子在南方稻区迅速蔓延,严重影响了水稻的产量和质量[3]。
乙酰辅酶A 羧化酶 (acetyl-CoA carboxylase,ACCase) 抑制剂是以ACCase 为作用靶标的一类除草剂,主要包括芳氧苯氧基丙烯酸酯类 (APPs)、环己烯酮类 (CHDs) 及苯基吡唑啉类 (PPZs),多用于苗后一年生禾本科杂草的防除。氰氟草酯、唑酰草胺和精唑禾草灵均属于APPs 除草剂,是目前防除水稻田千金子的主要药剂。近年来,上海市多个地区水稻田千金子使用APPs 除草剂防除失败,表现出较严重的抗药性,已成为该地区稻田杂草防除的一大难题[4]。研究表明,ACCase 抑制剂类除草剂极易导致杂草抗药性的产生[5-6]。截至目前,全球已报道49 种杂草对该类除草剂产生了抗性,2002 年首次报道了千金子对ACCase 抑制剂类除草剂的抗性[7]。靶标酶基因位点突变是杂草对ACCase 抑制剂类除草剂最常见的抗性机制,目前已发现7 个ACCase 氨基酸位点突变能够导致杂草对该类除草剂产生抗性,分别是第1781 位异亮氨酸 (Ile)、第1999 位色氨酸 (Trp)、第2027 位Trp、第2041 位Ile、第2078 位天冬氨酸 (Asp)、第2088 位半胱氨酸 (Cys) 以及第2096 位甘氨酸 (Gly)[8-10]。Yu 等[11]在 2016 年首次报道了ACCase 基因第2027 位Trp 突变为Cys 是千金子对ACCase 抑制剂类除草剂产生抗性的原因。此外,研究发现ACCase 抑制剂类除草剂被快速代谢解毒也是杂草产生抗药性的重要原因之一[12-13]。文马强等[2]研究发现,千金子抗性种群对氰氟草酯产生抗性可能与谷胱甘肽S-转移酶(GSTs)、超氧化物歧化酶 (SOD)、过氧化物酶(POD) 及过氧化氢酶 (CAT) 的活力增强有关。目前关于抗性千金子对除草剂降解和代谢的研究仍局限于酶活性测定的初步探究阶段[8]。
本研究在上海市千金子发生较为严重的地区采集了51 个千金子种群,测定了其对ACCase 抑制剂类除草剂氰氟草酯、唑酰草胺和精唑禾草灵的抗性水平,并对抗性种群进行了靶标酶基因检测,以期进一步明确上海市水稻田抗性千金子种群的发生情况及可能存在的靶标抗性机制,旨在为千金子防除策略制定和延缓其抗性种群发展提供科学依据。
1 材料与方法
1.1 供试材料
1.1.1 千金子种子 2016 年和2017 年9‒10 月采集于上海市千金子发生较为严重的8 个区 (崇明区、奉贤区、嘉定区、金山区、闵行区、浦东新区、青浦区及松江区) 水稻田,共采集51 个千金子种群,所有种子晾干后置于4 ℃冰箱中储存,待用。采集地点等具体信息见表1。
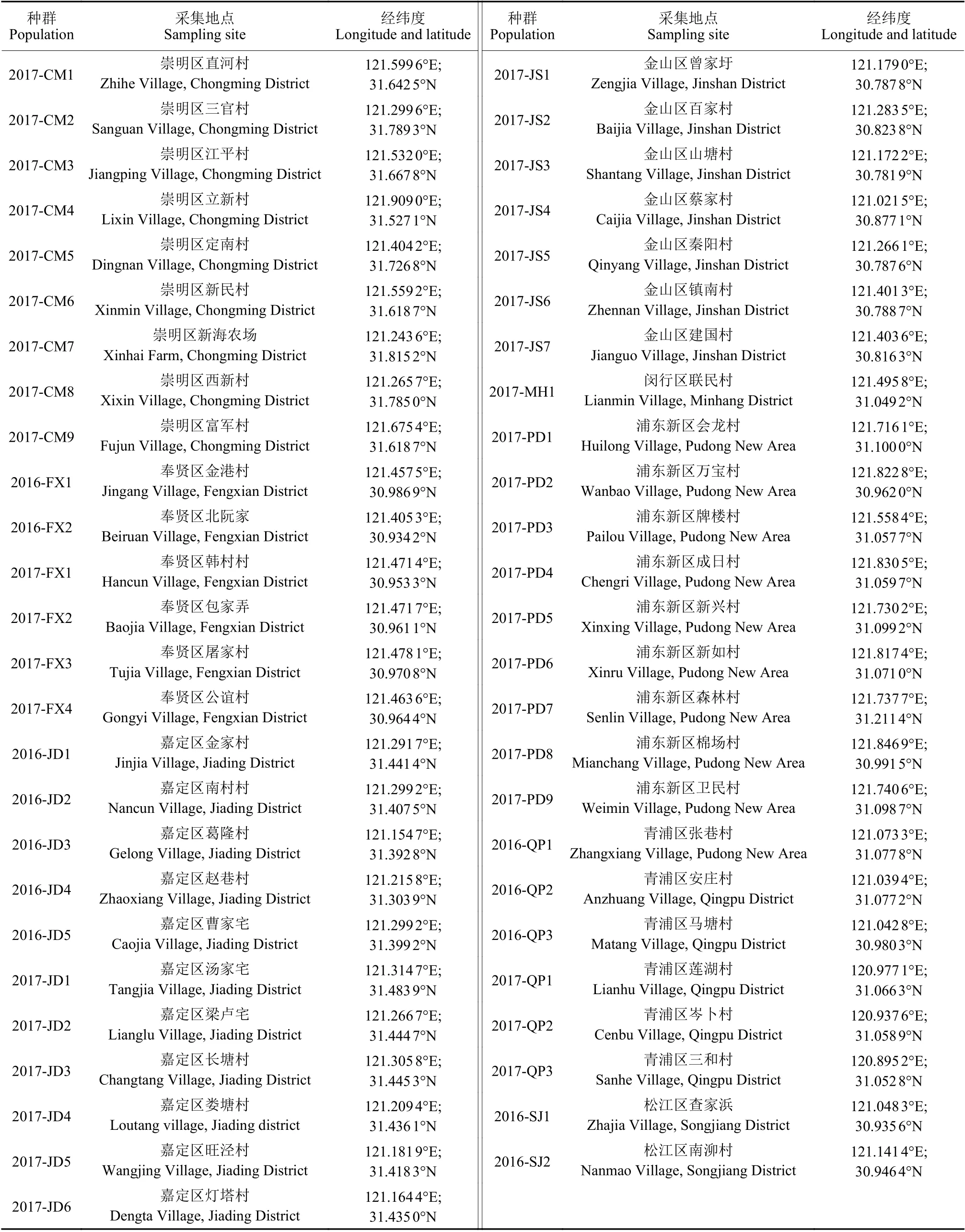
表1 千金子种群的采集地点Table 1 Source locations of the L. chinensis populations examined in this study
1.1.2 除草剂 100 g/L 氰氟草酯 (cyhalofop-butyl)乳油 (EC),美国陶氏益农公司;10%唑酰草胺(metamifop) 乳油,美国富美实公司;69 g/L 精唑禾草灵 (fenoxaprop-P-ethyl) 水乳剂 (EW),拜耳股份公司。
1.1.3 生物试剂和主要仪器 高效植物基因组DNA 提取试剂盒 (Hi-DNAsecure Plant Kit),天根生化科技 (北京) 有限公司;Taq酶 (TransStart®TopTaqDNA Polymerase),北京全式金生物技术(TransGen Biotech) 有限公司。ASS-4 型化学农药实验自动控制喷洒系统,北京盛恒天宝科技有限公司生产;5804R 型高速冷冻离心机,德国 Eppendorf公司;T100 Thermal Cycler PCR 仪,美国Bio-Rad公司;Tannon-1600 全自动数码凝胶图像处理系统,上海天能科技有限公司;DYY-8C 型电泳仪,北京六一生物科技有限公司;GR60DR 高压灭菌器,致微 (厦门) 仪器有限公司;SW-CJ-1FD 洁净工作台,苏州安泰空气技术有限公司;MGC-450HP人工气候箱,上海一恒科学仪器有限公司。
1.2 试验方法
1.2.1 千金子种群室内整株生物测定 采用单剂量抗性甄别法[14]测定各千金子种群对ACCase 抑制剂类除草剂氰氟草酯、唑酰草胺和精唑禾草灵的敏感性。从各千金子种群中分别选取均匀一致的种子播种于填有壤土,长、宽各为10 cm的方形塑料盆钵内,每盆播种20 粒,置于人工气候箱内培养 (光周期L/D:12 h/12 h;白天 30 ℃,晚上25 ℃;相对湿度 70%)。待千金子长至2 叶期时间苗,每盆保留长势一致的幼苗15 株。于3~4 叶期时采用ASS-4 型自动控制喷洒系统进行茎叶喷雾处理,用水量为450 L/hm2,喷雾压力为0.275 MPa。3 种除草剂有效成分施用剂量分别为:氰氟草酯105 g/hm2,唑酰草胺120 g/hm2,精唑禾草灵62.1 g/hm2;以清水处理为空白对照。药后21 d 调查记录每个种群的存活株数,计算存活率。每处理4 次重复,整体试验重复2 次。
1.2.2 千金子种群ACCase 基因突变检测 基于
1.2.1 节的抗性测定结果,从各发展中抗性 (DR)种群和抗性 (R) 种群中分别随机选取60 株千金子幼苗,利用Hi-DNAsecure Plant Kit 分别提取单株的基因组DNA。参考已报道的千金子引物进行ACCase 基因片段扩增[9],所用引物序列分别为QJ-F:5′-ATTCCTATGGAGCGTCCTG-3′ 和QJR:5′- TCCTTCGCAAGAACATCC-3′,所扩增序列包含了目前已知能够赋予抗性的所有突变位点。引物由生工生物工程 (上海) 股份有限公司合成。PCR 扩增反应体系体积为50 μL,包括基因DNA 模板2 μL,正反向引物 (10 μmol/L) 各1 μL,TransStart® TopTapDNA Polymerase (2.5 units/μL)1 μL,10 ×TransStart® TopTapBuffer 5 μL,dNTPs (2.5 mmol/L) 4 μL,ddH2O 36 μL。PCR 反应条件:94 ℃预变性5 min;94 ℃变性30 s,52 ℃退火30 s,72 ℃延伸90 s,34 个循环;72 ℃终延伸10 min。反应结束后,取5 μL PCR 扩增产物于1% 琼脂糖凝胶中进行电泳检测,将含目的条带的PCR 扩增产物送生工生物工程 (上海) 股份有限公司测序。测序所得核苷酸序列于NCBI 库中进行BLAST 比对,以确定所得序列是否为目标基因序列。采用Chromas 2.23 软件分析ACCase 基因的Ile-1781、Trp-1999、Trp-2027、Ile-2041、Asp-2078、Cys-2088、Gly-2096 氨基酸位点处是否发生了碱基置换、移位、缺失和插入等非同义突变情况。
1.3 数据分析
使用Microsoft Excel 2016 软件进行数据统计分析。根据千金子种群存活率判定抗性程度:存活率小于1%为敏感 (S) 种群,介于1%~20%之间为发展中抗性 (DR) 种群,大于20%则为抗性 (R)种群[15-16]。
2 结果与分析
2.1 不同千金子种群对3 种ACCase 抑制剂类除草剂的抗性发生程度
由表2 可看出,在氰氟草酯105 g/hm2有效成分剂量选择压下,2016-JD1、2016-JD2、2017-CM1、2017-CM3、2017-CM9、2017-JD4、2017-JD5 和2017-JS2 种群的存活率均大于23%,标记为抗性种群;2016-JD5、2017-JD2、2017-JD3、2017-JS1、2017-PD1 和2017-PD5 种群的存活率在2%~19%之间,标记为发展中抗性种群;其余千金子种群均为敏感种群,存活率为0。由抗性千金子种群的分布范围看,上海市嘉定区、崇明区、金山区和浦东新区水稻田发现的抗性种群和发展中抗性种群较多。

表2 不同千金子种群在氰氟草酯105 g/hm2 有效成分剂量处理下的存活率和抗性分类Table 2 The survival rate and resistant category of the L. chinensis populations against cyhalofop-butyl at 105 g (a.i.)/hm2
表3 不同千金子种群在唑酰草胺120 g/hm2 有效成分剂量下的存活率和抗性分类Table 3 The survival rate and resistant category of the L. chinensis populations against metamifop at 120 g (a.i.)/hm2

表3 不同千金子种群在唑酰草胺120 g/hm2 有效成分剂量下的存活率和抗性分类Table 3 The survival rate and resistant category of the L. chinensis populations against metamifop at 120 g (a.i.)/hm2
注:a 敏感种群 (S):存活率 <1%;发展中抗性种群 (DR):存活率在1% ~ 20%之间;抗性种群 (R):存活率 >20%。Note: a Susceptible population (S): survival rate <1%; development of resistance population (DR): 1%≤ survival rate ≤20%; resistant population (R):survival rate >20%.
2016-FX1 0 S 2017-JD2 5.0 DR 2016-FX2 0 S 2017-JD3 0 S 2016-JD1 62.5 R 2017-JD4 0 S 2016-JD2 100 R 2017-JD5 6.7 DR 2016-JD3 0 S 2017-JD6 0 S 2016-JD4 0 S 2017-JS1 0 S 2016-JD5 0 S 2017-JS2 0 S 2016-QP1 0 S 2017-JS3 0 S 2016-QP2 0 S 2017-JS4 0 S 2016-QP3 5.8 DR 2017-JS5 0 S 2016-SJ1 0 S 2017-JS6 0 S 2016-SJ2 0 S 2017-JS7 0 S 2017-CM1 28.3 R 2017-MH1 0 S 2017-CM2 0 S 2017-PD1 0 S 2017-CM3 5.0 DR 2017-PD2 0 S 2017-CM4 0 S 2017-PD3 0 S 2017-CM5 0 S 2017-PD4 0 S 2017-CM6 0 S 2017-PD5 3.3 DR 2017-CM7 0 S 2017-PD6 0 S 2017-CM8 0 S 2017-PD7 0 S 2017-CM9 45.0 R 2017-PD8 0 S 2017-FX1 0 S 2017-PD9 0 S 2017-FX2 0 S 2017-QP1 0 S 2017-FX3 0 S 2017-QP2 0 S 2017-FX4 0 S 2017-QP3 0 S 2017-JD1 0 S
表4 不同千金子种群在精唑禾草灵62.1 g/hm2 有效成分剂量下的存活率和抗性分类Table 4 The survival rate and resistant category of the L. chinensis populations against fenoxaprop-P-ethyl at 62.1 g (a.i.)/hm2
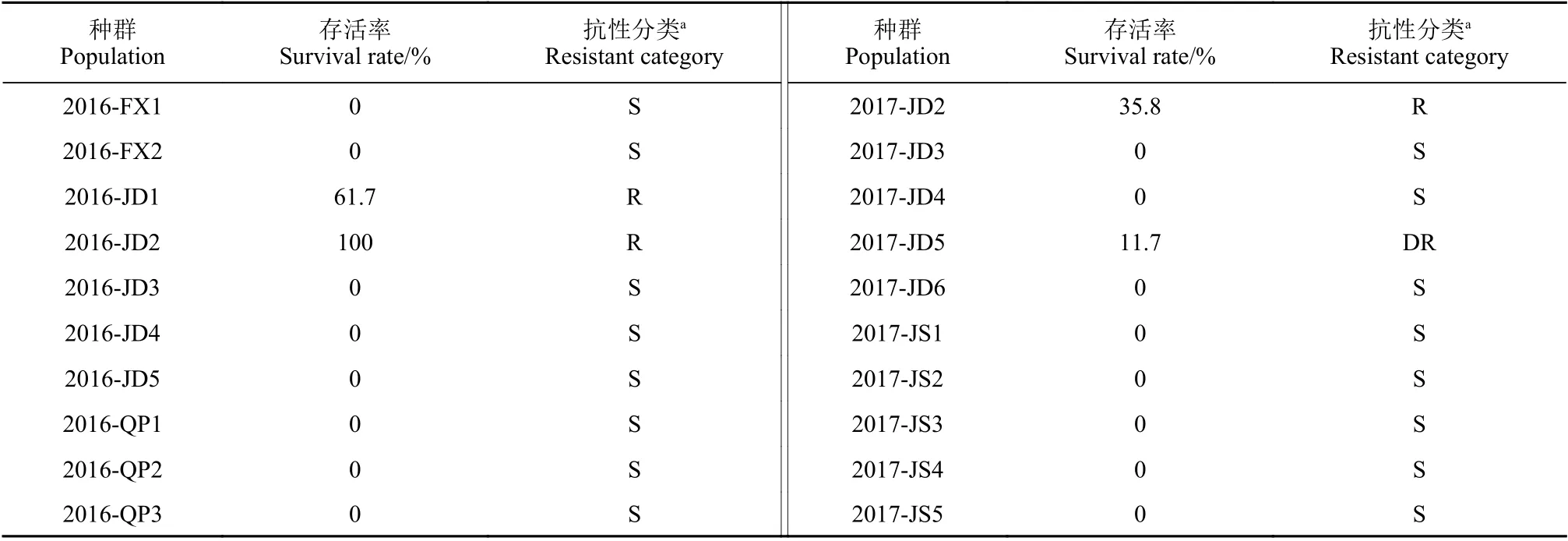
表4 不同千金子种群在精唑禾草灵62.1 g/hm2 有效成分剂量下的存活率和抗性分类Table 4 The survival rate and resistant category of the L. chinensis populations against fenoxaprop-P-ethyl at 62.1 g (a.i.)/hm2
种群Population抗性分类a Resistant category 2016-FX1 0 S 2017-JD2 35.8 R 2016-FX2 0 S 2017-JD3 0 S 2016-JD1 61.7 R 2017-JD4 0 S 2016-JD2 100 R 2017-JD5 11.7 DR 2016-JD3 0 S 2017-JD6 0 S 2016-JD4 0 S 2017-JS1 0 S 2016-JD5 0 S 2017-JS2 0 S 2016-QP1 0 S 2017-JS3 0 S 2016-QP2 0 S 2017-JS4 0 S 2016-QP3 0 S 2017-JS5 0 S存活率Survival rate/%抗性分类a Resistant category种群Population存活率Survival rate/%
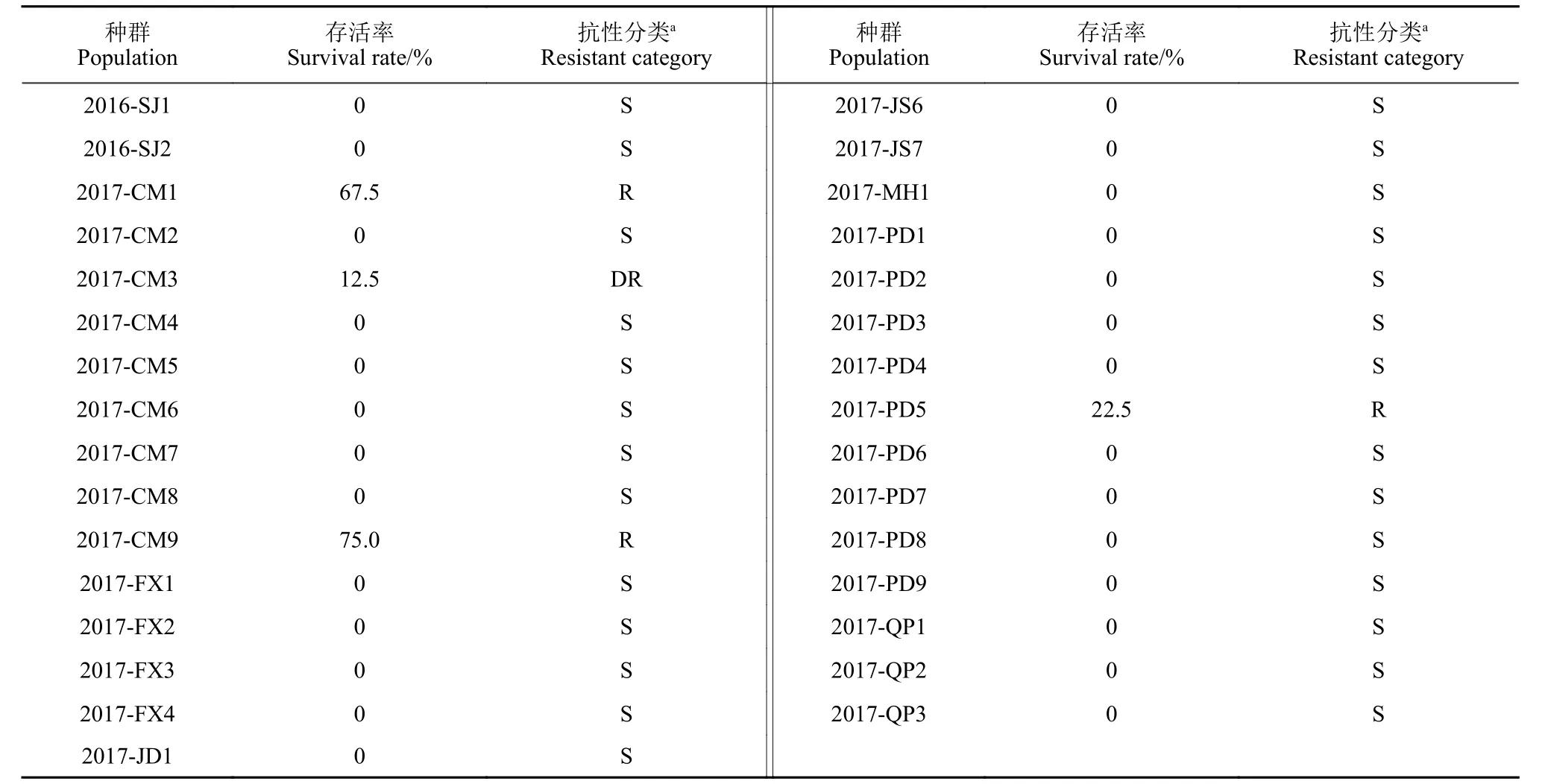
续表4Table 4 (Continued)
2.2 不同千金子种群ACCase 基因抗性突变鉴定
BLAST 比对结果表明,所得核苷酸序列与亲缘关系较近的大穗看麦娘Alopecurus myosuroides对应基因的同源性均大于95%。将翻译后的氨基酸序列与大穗看麦娘进行比对,结果显示:2016-JD1、2016-JD2、2017-CM1、2017-CM3、2017-CM9、2017-JD2、2017-JD5、2017-PD1 和2017-PD5 这9 个种群均发生了靶标基因突变 (表5)。其中,2016-JD1 和2016-JD2 种群ACCase 基因第1999 位氨基酸密码子由TGG 突变为TCG,导致Trp 突变为Ser;2017-JD2、2017-JD5 和2017-PD5 种群ACCase 基因第1999 位氨基酸密码子由TGG 突变为TGT,导致Trp 突变为Cys;2017-CM3、2017-CM9 和2017-PD1 种群ACCase 基因第2027 位氨基酸密码子由TGG 突变为 TCG,导致Trp 突变为Ser;在2017-CM1、2017-CM3 和2017-JD2 种群中发现了ACCase 基因第2027 位氨基酸Trp 被Cys 取代,该突变由密码子TGG 突变为TGT 或TGC 所致。本研究发现,2017-CM3 和2017-JD2 种群同时存在两种ACCase 基因突变类型,而2016-JD5、2016-QP3、2017-JD3、2017-JD4、2017-JS1 和2017-JS2 这6 个种群均未发现已知氨基酸位点的突变。

表5 抗性千金子种群中ACCase 基因突变形式及突变频率Table 5 ACCase gene mutation form and mutation ratio of the resistant L. chinensis populations
通过对各千金子种群单株测序结果进行分析,得到不同种群的ACCase 基因突变频率 (表5)。2016-JD1、2016-JD2、2017-CM1、2017-CM9 和2017-JD5 5 个种群的靶标基因突变频率均大于60%,其中2016-JD2 种群突变频率最高,达96.7%;2017-JD2、2017-PD1 和2017-PD5 种群的突变频率 ≤ 20%, 其中2017-PD1 种群突变频率最低,仅为5%。
3 结论与讨论
氰氟草酯由美国陶氏益农公司开发,对千金子有特效,于2006 年在我国正式登记用于水稻田苗后防除禾本科杂草[17]。唑酰草胺是由韩国东部韩农化学公司开发的水稻田禾本科杂草的苗后除草剂,于2010 年在我国登记[18]。精唑禾草灵由德国拜耳公司开发,加入安全剂解草唑 (fenchlorazole) 后,于1998 年登记用于小麦田禾本科杂草防除[19]。根据中国农药信息网 (http://www.icama.org.cn/) 的查询结果,精唑禾草灵和氰氟草酯的复配制剂于2009 年首次登记用于水稻田防除一年生禾本科杂草。因此,氰氟草酯、唑酰草胺和精唑禾草灵在我国水稻田应用均已有十多年的历史。ACCase 抑制剂类除草剂属于高抗性风险级别[20],据报道,该类除草剂连续重复使用6~10 年就可导致杂草产生抗药性[21]。本研究所测的上海市51 个千金子种群中,8 个种群对氰氟草酯可标记为抗性,6 个种群为发展中抗性;4 个种群对唑酰草胺可标记为抗性,5 个种群为发展中抗性;6 个种群对精唑禾草灵可标记为抗性,2 个种群为发展中抗性。分析所得结果可以看出,抗氰氟草酯、抗唑酰草胺和抗精唑禾草灵的千金子种群之间存在着一定程度的一致性,但又并非完全一致,这可能与采集地点作物种植模式、用药历史背景以及其抗性机制等因素不同有关。
靶标抗性机制和非靶标抗性机制是杂草对ACCase 抑制剂类除草剂产生抗性的两大原因[22],多数情况下,ACCase 基因靶标位点突变是导致杂草抗药性产生的主要原因[23]。到目前为止,已报道7 种ACCase 氨基酸突变方式与千金子的抗药性相关,分别是Ile-1781-Leu、Ile-1781-Val、Trp-1999-Cys、Trp-1999-Ser、 Trp-2027-Cys、Trp-2027-Ser 和Ile-2041-Asn[4,24-26]。本研究对15 个抗性千金子种群的 ACCase 基因部分片段进行了克隆和序列比对,在9 个抗性种群中共发现了4 种氨基酸突变方式:Trp-1999-Ser、Trp-1999-Cys、Trp-2027-Ser 和 Trp-2027-Cys。因此,ACCase 基因中1999 位点及2027 位点的氨基酸突变可能是导致千金子对ACCase 抑制剂类除草剂产生抗性的重要原因之一,这一结论与杨浩娜等[8]关于千金子对氰氟草酯产生的较高抗性与其靶标基因2027 和1999 位点氨基酸突变有关的报道一致。武向文等[4]首次报道了Trp-1999-Ser 突变是千金子对氰氟草酯产生抗性的原因,但其未测定交互抗性问题。本研究发现,ACCase 基因Trp-1999-Ser 发生突变的千金子种群可对氰氟草酯、唑酰草胺和精唑禾草灵产生交互抗性。Powles 等[27]报道,通常ACCase 基因1999、2027、2041 和2096 位点的氨基酸突变会导致杂草对一种或多种APPs 除草剂产生抗性。本研究结果与该报道一致。此外,本研究还发现,抗性千金子种群的ACCase 基因突变频率与其对氰氟草酯、唑酰草胺和精唑禾草灵的抗性程度并不一致。Powles等[27]的研究还表明,杂草对ACCase 抑制剂类除草剂的交互抗性水平与靶标酶基因突变方式、突变基因型以及ACCase 抑制剂类除草剂的种类相关。Deng 等[26]报道千金子具有两个ACCase基因拷贝,存在基因稀释效应。Iwakami 等[28]研究发现,靶标酶基因的稀释效应也会影响抗性突变体对除草剂的抗性水平。
与靶标抗性机制相比,由非靶标抗性机制引起的杂草抗药性更为复杂,更加难以预测[29]。非靶标抗性机制包括杂草对除草剂吸收和转运的减少,对除草剂的隔离屏蔽以及代谢解毒能力增强等[30]。本研究中有6 个抗性千金子种群未发现已报道的氨基酸突变方式,因此推断这些抗性种群中由非靶标抗性机制参与的可能性较高。Deng等[13]的研究表明,千金子对氰氟草酯代谢分解速率的快慢与其抗药性水平密切相关。Chen 等[31]研究发现,细胞色素P450 单加氧酶 (P450s) 抑制剂马拉硫磷 (malathion) 能显著逆转抗性千金子种群对氰氟草酯的抗药性,表明其抗药性的产生与P450s 活性增强有关。本研究仅从靶标抗性方面鉴定了不同抗性种群靶标酶基因的突变,而其抗性中是否存在非靶标抗性则需要进一步研究。
本研究中,上海市部分地区水稻田千金子对ACCase 抑制剂类除草剂的抗性已较为严重,且靶标酶基因突变是导致不同千金子种群产生抗性的重要原因之一,而长期、频繁地使用相同类型的除草剂防除同一种杂草,会加速杂草抗药性的发生。因此,针对水稻田千金子发生区域,尤其是抗性千金子发生区域,不能再单一使用ACCase抑制剂类除草剂,应提倡采用多元化的综合治理措施,如多种作用机制的除草剂交替或混配使用,以及直播稻与移栽稻轮作等方法,从而尽量控制和延缓杂草抗药性的发展。
