复合酶制剂体外预消化仔猪饲粮最佳条件的筛选以及对其品质的影响
2022-06-10武一帆刘思均李步高曹果清郭晓红
杨 阳 王 悦 武一帆 刘思均 赵 燕 李步高 曹果清 郭晓红
(山西农业大学动物科学学院,太谷030801)
我国饲料原料资源紧张,最大化提高饲粮养分的消化率和利用率是缓解饲料资源短缺的有效方式之一。长期以来,玉米-豆粕型配合饲粮作为传统的单胃动物饲粮配方被养殖业广泛应用,但配合饲粮饲料成分复杂,原料中的营养物质被细胞壁包围,这影响了营养物质的利用。此外,饲粮中含有较多的抗营养物质如非淀粉多糖(NSP)等,这阻止了饲粮中营养物质的释放,饲粮利用效果的稳定性不高[1-2]。研究发现,饲粮添加酶制剂可以增加可消化营养物质含量,并降低抗营养因子等,是一种提高饲粮利用率的有效途径[3];但受到催化底物单一性以及饲粮成分复杂性的影响,单一的酶制剂作用效果有限,因此饲粮中添加复合酶制剂得到广泛应用[4]。
仔猪断奶后,饲粮由液态的母乳转变为颗粒状的固态饲料,仔猪由于不适应饮食状态的转变,会出现食欲不振,抗病力降低,肠道有害菌感染的风险增加[5]。而在饲喂过程中,酶制剂主要是随饲粮添加进行饲喂,酶制剂最适的温度、时间和pH没有实现,使得酶没有发挥最佳效果[6]。液态饲料的形状与母乳相似,饲喂液体饲料可以减少对仔猪胃肠道的刺激,提高饲料的转化率,并能降低抗营养因子,抑制有害菌的生长,有效减轻断奶应激压力[7]。目前,在液态饲料中添加复合酶制剂预消化的研究报道还较少,酶解预消化的程序和作用效果还有待进一步建立和评定。
木聚糖是植物细胞壁的重要组成成分,连接细胞壁中的木质素和纤维素。玉米的木聚糖主要是由葡萄糖和阿拉伯木聚糖组成,支链复杂,且由于木聚糖支链结构的存在,木聚糖不能被单一的木聚糖酶所降解[8]。阿拉伯呋喃糖苷酶(arabinofuranosidase,AF)和阿魏酸酯酶(ferulic acid esterase,FAE)可特异性降解木聚糖支链上的阿拉伯呋喃糖苷键和阿魏酸酯键,从而可促进木聚糖酶对木聚糖的水解作用[9]。葡萄糖氧化酶(glucose oxidase,GOD)是一种以β-D-葡萄糖为底物的脱氢酶制剂,水解产物能够抑制肠道内病原微生物的生长,降低肠道pH,提高肠道酸性酶的活性[10-12]。目前,很少有研究评价复合酶制剂中GOD、木聚糖主链酶和支链酶协同作用的添加效果。因此,本研究通过仔猪饲粮中添加复合酶制剂,筛选体外预消化的最佳作用条件,并鉴定其作用效果,以完善饲料体外预消化技术体系,为断奶仔猪无抗饲粮的研发提供参考。
1 材料与方法
1.1 试验材料
饲用酶制剂:酸性蛋白酶、α-淀粉酶和纤维素酶购于湖北某公司,果胶酶、木聚糖酶、β-葡聚糖酶、α-半乳糖甘酶和GOD购于宁夏某集团,FAE和AF购于Megazyme公司,酶学特性见表1。酶解产物标准物:低聚木糖(木一糖、木二糖、木三糖和木四糖)标准品、阿魏酸标准品和葡萄糖酸标准品均购自中国索莱宝公司。
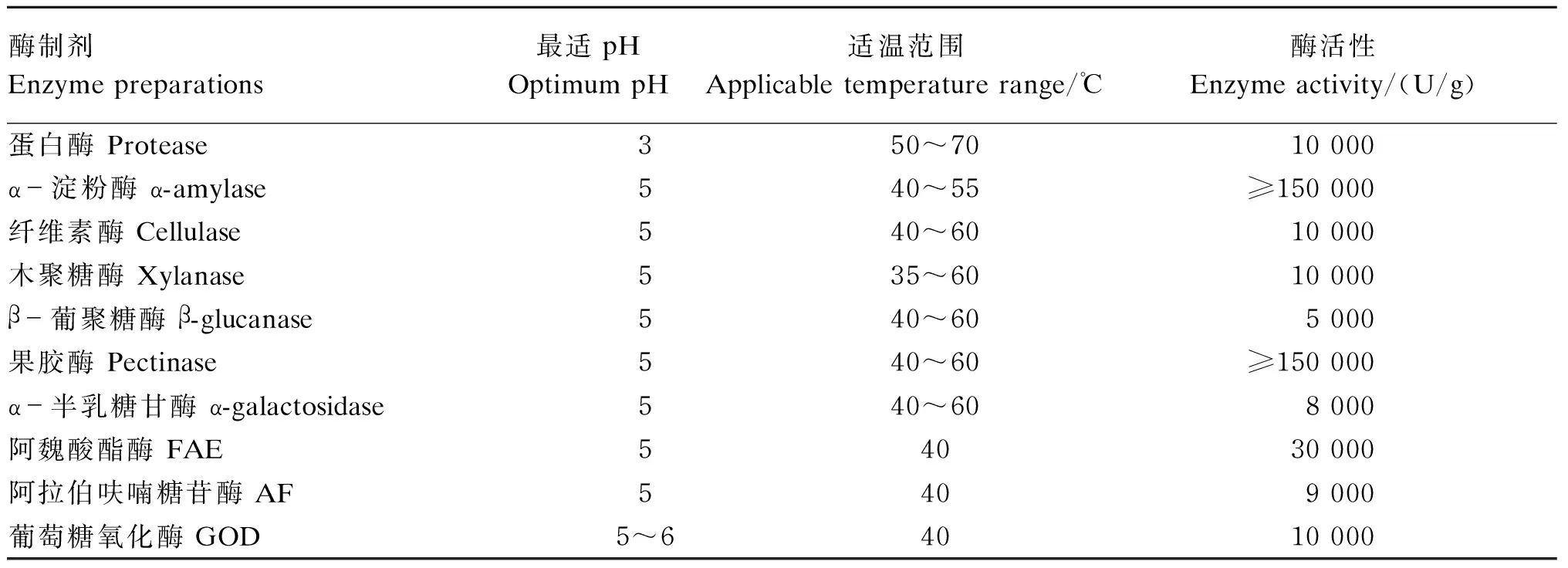
表1 饲用酶制剂酶学特性
1.2 试验用仔猪饲粮
本试验所用基础饲粮为玉米-豆粕型,从山东某公司购买,不添加任何饲料酶制剂及抗生素,其组成及营养水平见表2。
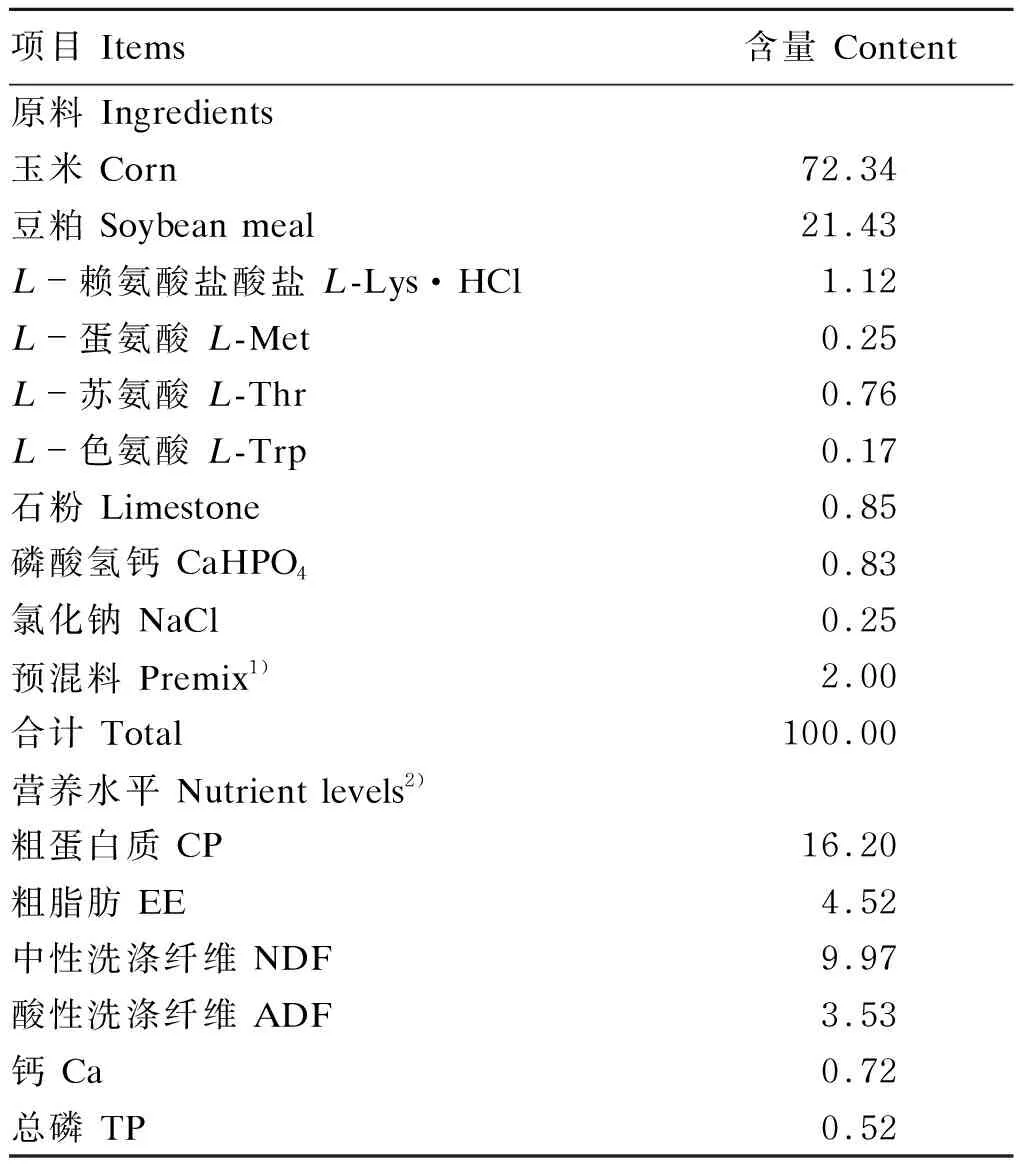
表2 基础饲粮组成及营养水平(风干基础)
1.3 试验设计
称取10 g待测饲粮样品于100 mL三角瓶中,加入蛋白酶100 U/g,按照固液比=1.0∶2.5的比例加入25 mL pH=3的乙酸-乙酸钠缓冲溶液后,在不同温度(50和60 ℃)的恒温摇床中进行不同时间(0.5、1.0、1.5和2.0 h)的预消化处理,检测酶解作用效果。
待确定条件后,将缓冲液调整为pH=5,添加NSP酶群(纤维素酶100 U/g、木聚糖酶100 U/g、葡聚糖酶50 U/g、果胶酶1 500 U/g和半乳糖苷酶 80 U/g)和淀粉酶1 500 U/g,不同温度(40、50和60 ℃)的恒温摇床中进行不同时间(0.5、1.0、1.5和2.0 h)的预消化处理,检测酶解作用效果。
此外,分析木聚糖酶和支链水解酶(AF 36 U/g和FAE 120 U/g)的协同作用效果,以及GOD 100 U/g的酶解作用效果,进一步确定最佳条件。
1.4 指标测定
1.4.1 氨基酸和小肽分子、还原糖和抗营养因子含量测定
酶解反应完成后,沸水浴5 min,灭酶活后,在3 000 r/min条件下离心20 min,吸取上清液用于后续测定。采用总氨基酸试剂盒和十二烷基硫酸钠聚丙烯酰胺凝胶电泳(SDS-PAGE)试剂盒测定总氨基酸和小肽分子的含量;采用二硝基水杨酸(DNS)法测定还原糖的含量,协同因子(degree of synergy,DS)为试验组(加酶组)的还原糖含量/对照组的还原糖含量。大豆球蛋白、β-伴大豆球蛋白、大豆胰蛋白酶抑制因子、蔗糖和棉子糖的含量均采用相对应的酶联免疫吸附试验(ELISA)试剂盒检测。
1.4.2 酶解产物阿魏酸、低聚木糖和葡萄糖酸含量测定
阿魏酸含量采用液相色谱法测定,检测条件如下:利用AltimaTMHPC-18.8色谱柱(4.6 nm×250 nm);流动相为已腈-0.5%乙酸(有机相∶水相=3∶7);柱温:35 ℃;流速:1 mL/min;进样量:20 μL;时间:30 min;检测波长:λ=320.0 nm。阿魏酸含量计算公式为:
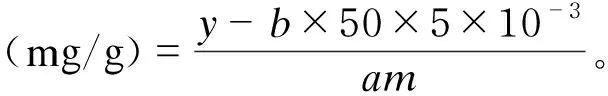
上机检测得阿魏酸标准溶液浓度与峰面积之间的线性关系,标准曲线为y=ax+b,式中:x表示标准溶液浓度(μg/mL);y表示色谱峰面积(mAU·min);m为样品的质量(g)。
低聚木糖含量采用高效阴离子交换色谱-脉冲安培检测(HPAEC-PAD)法测定,色谱柱:Carbo-Pac PA200 column (3 mm×250 mm);流动相:0.1 mol/L氢氧化钠(NaOH)溶液;柱温:35 ℃;流速:0.9 mL/min;进样量:10 μL;计算公式同阿魏酸含量。
葡萄糖酸含量采用HPAEC-PAD法测定,色谱柱:Carbo-Pac PA200 column (3 mm×250 mm);流动相:0.5 mol/L氢氧化钾(KOH)溶液;柱温:30 ℃;流速:1 mL/min;进样量:25 μL;计算公式同阿魏酸含量。
1.4.3 饲粮酶解前后的结构变化
利用扫描电镜和激光共聚焦显微镜观察饲粮酶解前后的结构变化。样品染色利用激光共聚焦观测,采用0.01%的荧光增白剂和罗丹明B染液分别染色10 min后,洗净染色液,用中性树脂胶封片,激光共聚焦观测,利用ImageJ软件定量检测平均荧光强度。
1.4.4 酶解产物对肠道有害菌生长曲线的测定
从-80 ℃冰箱中取出大肠杆菌和沙门氏菌保存菌,设置对照组(单菌落+培养基)和试验组(单菌落+酶解液),将菌活化后涂布于LB固体培养基,置于37 ℃恒温培养箱静置培养24 h,挑取单菌落接种于LB液体培养基中,振荡(180 r/min)培养,每小时取样,在600 nm测定吸光度(OD)值,绘制生长曲线。
1.5 数据统计分析
数据统计分析利用SPSS 20.0统计软件完成,数据间的比较采用t检验;采用单因素方差和LSD法进行分析,差异显著性采用Duncan氏法;抗营养因子含量的比较采用双因素方差分析。P<0.05和P<0.01分别表示差异显著和差异极显著,结果采用“平均值±标准差”表示。
2 结果与分析
2.1 蛋白酶、淀粉酶和NSP酶的最佳作用温度和时间
为了确定复合酶制剂的最佳作用条件,本试验首先在pH=3的体系中,加入蛋白酶反应,结果显示,当温度为50 ℃时,加酶组总氨基酸含量极显著高于对照组(P<0.01)(图1-A);同时,SDS-PAGE结果也发现,加酶组饲粮中的蛋白质由大分子被分解成较小的分子(图1-B);蛋白酶的催化效果在反应0.5 h后已经达到了较好的效果(图1-C)。为了确定淀粉酶和NSP酶群的最佳作用条件,试验随后在pH=5的体系中,加入淀粉酶和NSP酶群,结果如图1-D所示,还原糖含量的高低顺序为:50 ℃>60 ℃>40 ℃,且加酶组还原糖含量均极显著高于相同温度的对照组(P<0.01)。随着酶解时间的增加,还原糖含量先逐渐升高后趋于平缓,并且在酶解1.0 h时还原糖的含量最高(图1-E)。
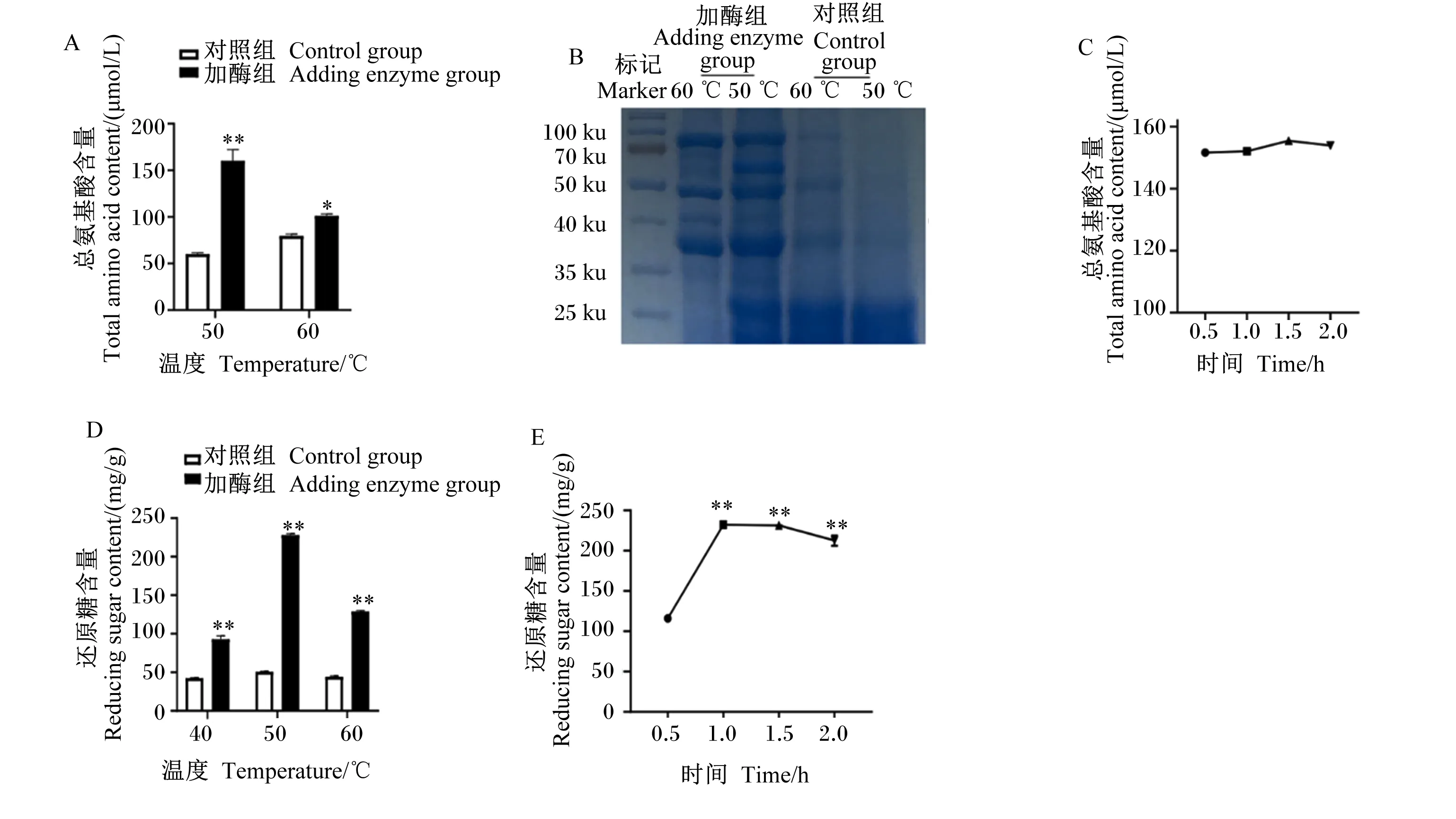
*表示差异显著(P<0.05),**表示差异极显著(P<0.01)。
由表3可知,NSP酶处理极显著影响饲粮中β-伴大豆球蛋白、大豆球蛋白、大豆胰蛋白酶抑制因子、棉子糖和蔗糖含量(P<0.01);其中,各温度条件下,NSP酶添加组β-伴大豆球蛋白、大豆球蛋白和蔗糖含量均显著低于未添加组(P<0.05)。温度和NSP酶的交互效应极显著影响β-伴大豆球蛋白、大豆球蛋白和蔗糖含量(P<0.01)。
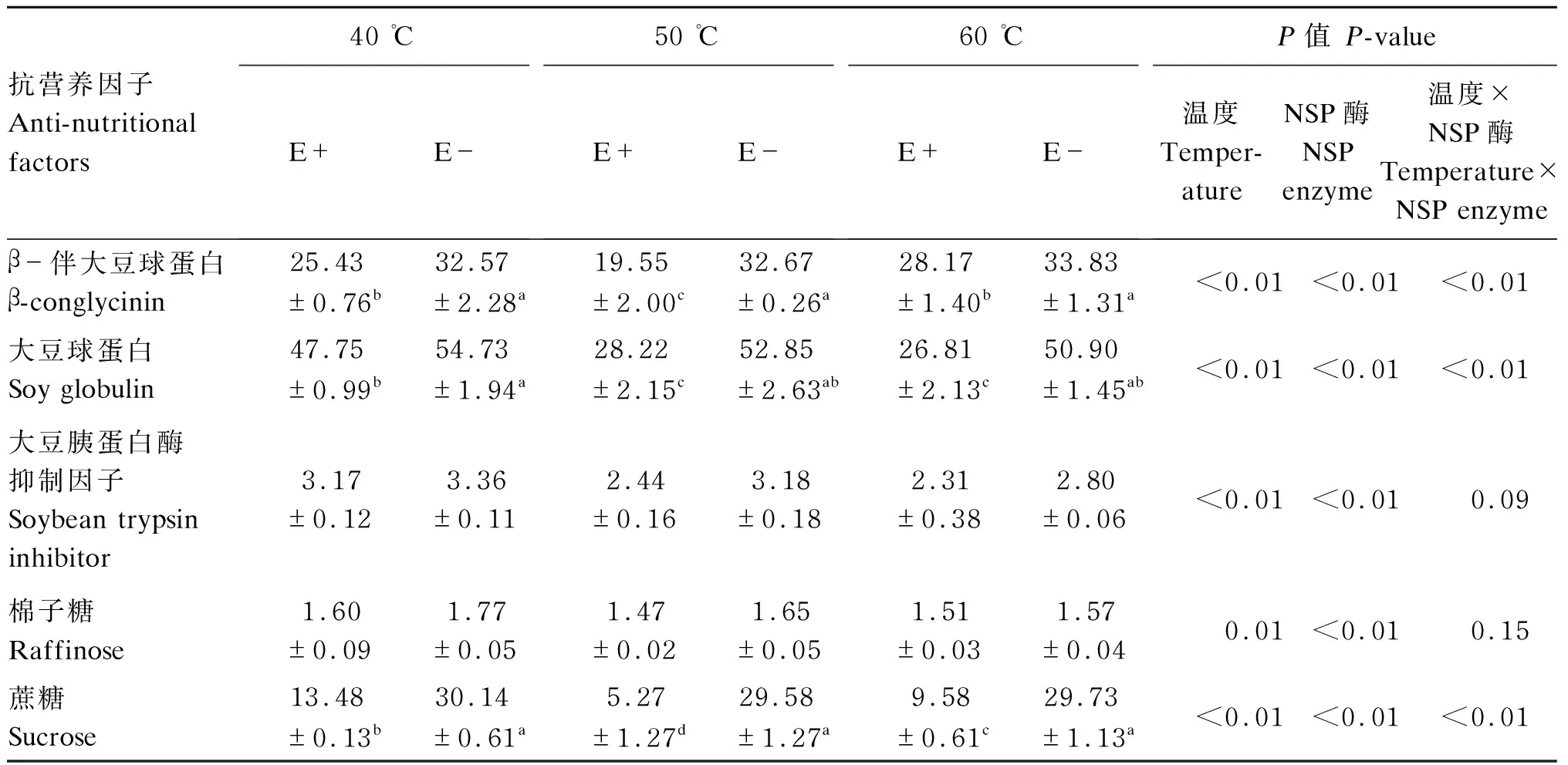
表3 饲粮中抗营养因子含量
2.2 酶解产物阿魏酸、低聚木糖和葡萄糖酸含量
为分析木聚糖主链酶和支链酶的协同作用效果,我们检测了不同添加顺序对木聚糖的分析效果,由表4可知,先加入AF和FAE、再加入木聚糖酶(处理3)或者同时加入木聚糖酶、AF和FAE(处理4),酶解产物中低聚木糖的含量较高,处理3木二糖含量为92.53 μg/mL、木三糖含量为68.73 μg/mL;处理3还原糖释放量(葡萄糖酸含量)最高,极显著高于其他处理(P<0.01),此时协同因子为1.56,与单独添加木聚糖酶(对照)水解饲粮相比,还原糖释放量提高了79.07%。
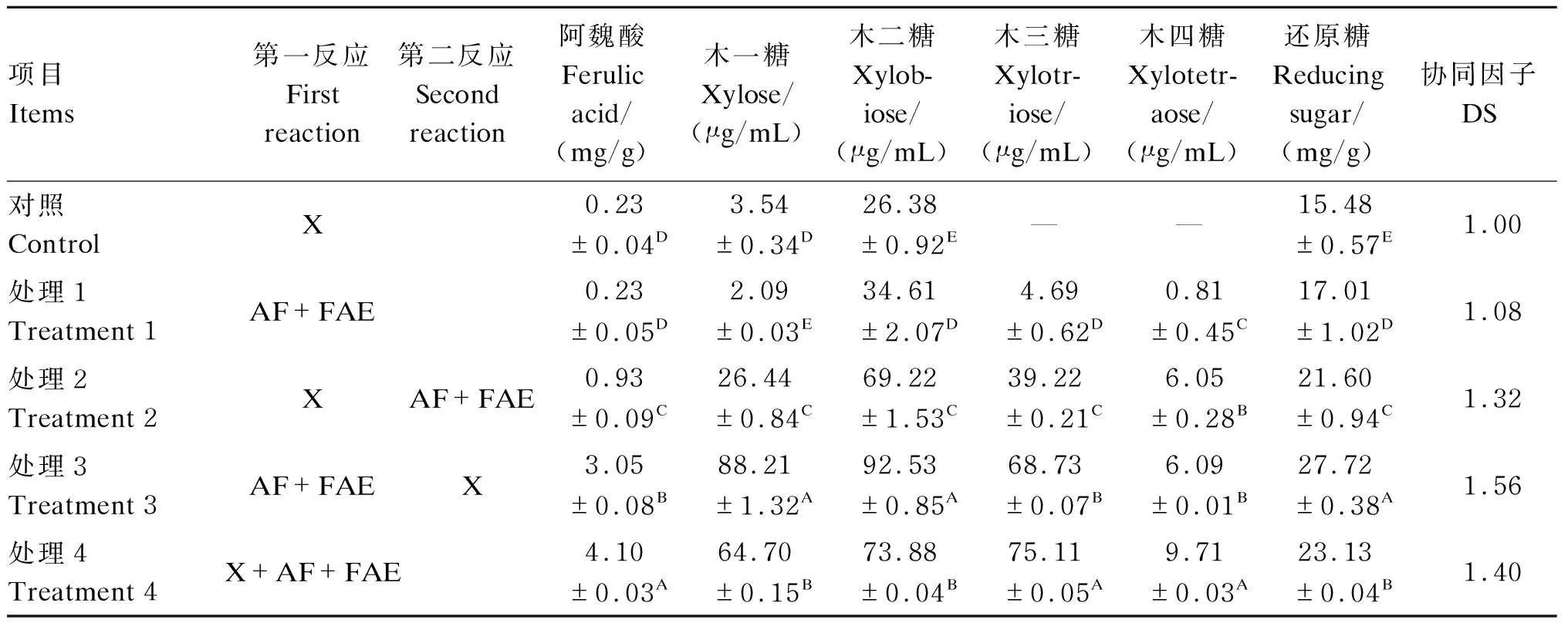
表4 饲粮酶解产物分析
GOD酶解产物和pH的分析见表5,由表可知,对照组中未检出葡萄糖酸,GOD组检出葡萄糖酸,其含量为1.32 μg/mL;并且2组相比,GOD组的pH下降。这说明GOD的酶解产物可以提供酸性环境。

表5 GOD酶解产物和pH
2.3 复合酶制剂对饲粮结构的影响
由前面建立的最佳条件,我们进行了复合酶制剂的体外预消化试验,饲粮结构电镜观察如图2-A,未经酶解的饲粮表面平整,饲粮经复合酶制剂酶解后,细胞表面皱缩,放大倍数观察,细胞壁界面模糊,表明细胞结构受到破坏。罗丹明B染色结果显示(图2-B),对照组饲粮单层细胞壁结构完整,细胞壁界限清晰,但在加酶组中,可清晰地看到细胞壁严重破坏。

A:饲粮在扫描电镜下的结构(500×,1 000×);B:罗丹明B染色后,饲粮在激光共聚焦显微镜下的结构(40×)。
2.4 酶解产物对肠道有害菌生长曲线的影响
为了验证复合酶制剂预消化饲粮后对肠道有害菌的影响,试验检测了复合酶制剂处理饲粮后酶解液对大肠杆菌和沙门氏菌生长情况的影响,由图3可知,对照组大肠杆菌和沙门氏菌的数量逐渐升高,而添加酶解液后(加酶组)细菌几乎不生长。
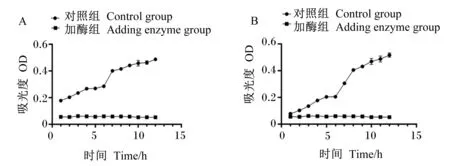
A:大肠杆菌的生长曲线;B:沙门氏菌的生长曲线。
3 讨 论
3.1 复合酶制剂对饲粮抗营养因子含量的影响
饲料酶制剂的使用提高了养分消化率,对断奶仔猪的生长性能有一定的改善作用。但是,以往的研究表明,酶制剂是随饲粮添加进行动物饲喂,并没有使得酶发挥最佳效果[13]。本试验应用复合酶制剂,筛选了充分发挥酶作用效果的条件,先是pH=3、50 ℃下,蛋白酶作用0.5 h;接着上调pH为5,50 ℃淀粉酶和NSP酶群作用1.0 h后,氨基酸和还原糖的含量达到最高,提示复合酶制剂提高了饲粮的可消化利用性。玉米-豆粕型饲粮中存在许多抗营养因子,如大豆中的抗原蛋白会损坏小肠绒毛和隐窝,引起过敏性腹泻[14];胰蛋白酶抑制因子会抑制胰蛋白酶和胰凝乳蛋白酶活性,降低蛋白质消化吸收和造成胰腺肿大[15]。单胃动物体内无降解棉子糖家族的α-半乳糖苷酶,未降解的寡糖进入大肠会被发酵分解,产生二氧化碳、甲烷或氨气,引起肠胃胀气及腹泻。研究发现,添加大豆可溶物(蔗糖和棉子糖等)可使生长猪肠道干物质、有机物及氮的消化能力显著降低[16]。本试验结果发现,加入蛋白酶、NSP酶和淀粉酶后,能够有效降低饲粮中抗营养因子的含量,说明酶解预消化处理能够充分发挥酶的催化作用,提高饲粮品质。
3.2 木聚糖主链酶与其支链酶的协同作用及对酶解产物的影响
由于木聚糖支链结构的存在,木聚糖不能被单一的木聚糖酶所降解。AF和FAE可特异性地降解阿拉伯呋喃糖苷键和阿魏酸酯键,从而促进木聚糖酶对木聚糖的水解作用[17]。很大程度上,协同因子的大小取决于木聚糖的结构和主链支链降解酶的酶解顺序。研究表明,首先添加AF和FAE去除木聚糖的支链结构,再添加木聚糖酶降解主链,更有利于木聚糖的降解[17]。本试验结果也显示,水解木聚糖时,先添加支链酶后添加主链酶,能最大程度地水解木聚糖。木聚糖酶与AF和FAE的协同作用,增加了饲粮中还原糖的生成量,同时也产生了低聚木糖。低聚木糖作为一种益生元,具有抗氧化和免疫调节功能,能够促进有益微生物的繁殖,维护动物肠道健康。研究显示,肉鸡饲粮中添加木聚糖主链酶和支链酶后,能够增加饲粮中低聚木糖含量,进而通过促进乳酸菌等有益菌的增殖,显著提高肉鸡的生长性能及改善肠道健康[18]。研究还发现,断奶仔猪饲粮中添加低聚木糖,能够降低肠道炎症水平,维持肠道屏障,提高乳酸杆菌等微生物的丰度[19]。因此,本试验筛选了AF、FAE和木聚糖酶协同作用方式,经过木聚糖主链酶和支链酶的协同作用,降低了木聚糖的含量,同时增加了还原糖和低聚木糖的生成。
3.3 复合酶制剂对饲粮结构及酶解产物对肠道有害菌的影响
目前,还原糖的生成量是评估酶制剂应用效果的主要指标,但NSP酶并不能完全分解纤维素和半纤维素,其发挥作用的主要方式是破坏植物细胞壁对营养物质的包裹。孟莹莹等[20]利用激光共聚焦技术,观察了玉米、麦麸和菜籽粕等原料在NSP酶解前后的细胞结构,发现复合酶制剂处理过的饲粮细胞壁发生断裂,包裹在其中的蛋白质部分被释放出来[20]。此外,也有研究者利用扫描电镜观察了酶制剂处理后麦麸的细胞结构变化,结果显示木聚糖酶处理后,麦麸表面呈现典型的蜂窝状结构,表明其结构受到了破坏[17]。本试验也利用扫描电镜和激光共聚焦技术,观察到了复合酶制剂处理后的饲粮细胞结构受到破坏,暴露出可被动物消化吸收的营养物质。因此,饲粮细胞结构变化直接观察结合还原糖生成量的测定,将更加全面地反映酶制剂的酶解效果。
GOD是一种以β-D-葡萄糖为底物的脱氢酶制剂,能够催化葡萄糖生成葡萄糖酸和过氧化氢。葡萄糖酸可以降低胃肠道内酸性,有利于抑制病原菌;过氧化氢具有广谱杀菌的功效,能够抑制大肠杆菌、沙门氏菌等致病微生物的增殖[10-12]。GOD在畜禽生产中被广泛使用,可以抑制病原菌及促进厌氧益生菌的增殖,维持肠道健康[21]。研究显示,在断奶仔猪饲粮中添加GOD不仅能降低胃肠道内食糜pH,改善仔猪肠道形态结构,还能促进饲粮中营养物质的消化,从而提高断奶仔猪平均日增重和饲料转化率[22]。此外,FAE分解产生的阿魏酸,也发挥着抑制细菌、真菌和原虫等有害微生物的生长的作用[23]。饲粮中添加FAE能够通过降低抗营养因子含量,抑制有害细菌并促进有益细菌生长,最终提高奶山羊的生产性能[24]。在本试验中也发现了包含FAE和GOD的复合酶制剂处理饲粮后,能够提高阿魏酸和葡萄糖酸的含量,降低饲粮的pH,并能有效抑制大肠杆菌和沙门氏菌的生长。综上所述,复合酶制剂体外预消化仔猪饲粮最佳条件的筛选,将为后续应用酶解预消化饲料,改善断奶仔猪肠道健康和提高生长性能的提供理论支持。
4 结 论
① 本研究通过筛选酶发挥作用的最佳条件,利用复合酶制剂预消化技术,最大效率地降解大分子物质。酶制剂的最佳组合方式为:首先在pH=3、50 ℃下,加入蛋白酶,作用时间为0.5 h;接着上调pH为5,加入AF、FAE和GOD,40 ℃作用1.0 h;最后加入淀粉酶和NSP酶群,50 ℃作用1.0 h。
② 经过酶解的饲粮,细胞壁遭到破坏,细胞中的营养物质可充分得到利用,酶解产物可以抑制有害菌的生长。
