植物单宁的微生物降解及其调控反刍动物瘤胃发酵的研究进展
2022-06-10申家豪郑娟善梁泽毅丁学智
申家豪 郑娟善 梁泽毅 丁学智,2*
(1.中国农业科学院兰州畜牧与兽药研究所,甘肃省牦牛繁育重点实验室,兰州730050;2.中国农业科学院兰州畜牧与兽药研究所,农业农村部兽用药物创制重点实验室,兰州730050)
我国动物源食品安全正受到生物污染和化学物污染的威胁。随着2020年国家饲料端“禁抗”,养殖端“减抗、限抗”兽用抗菌药使用减量化模式的推行,寻找天然绿色、安全的抗生素替代品已成为畜牧业发展的焦点。近年来,单宁作为一种天然绿色的新型饲料添加剂在饲料行业中被广泛关注。植物单宁(plant tannins)又称植物多酚,是植物以碳为基础形成的除木质素以外含量最丰富的复杂酚类次生代谢产物[1],广泛存在于五倍子、栗木、茶、葡萄、高粱和咖啡等植物的根[2]、茎[3]、叶[4]、花[5]等部位及种子[6]中。植物单宁可与家畜的唾液蛋白在口腔内发生沉淀反应,并对消化道内的微生物及消化酶产生拮抗,从而降低家畜的采食量和消化率[7]。同时,植物单宁具有广谱的抑菌性,会降低草食动物对营养物质的吸收率。因此,植物单宁一直以来都被认为是饲料中的抗营养因子。近年来,随着研究的深入,动物营养学家对植物单宁的抗营养作用有了重新认识。植物单宁产生抗营养作用的主要原因为其相对分子质量较大和添加浓度过高,而在饲粮中添加适量的植物单宁不仅可提高畜禽的采食量和平均日增重、抑制寄生虫、改善生产性能,而且在维持动物机体微生态平衡及减少甲烷和氮排放等方面也发挥了重要的作用[8]。然而,目前植物单宁在家畜饲粮中的有效添加剂量尚不确定,其微生物降解途径及调控瘤胃发酵的机理尚不明晰。因此,探究饲粮中植物单宁的适宜添加量与大分子植物单宁的生物降解途径,从而降低或消除植物单宁的抗营养作用,使其作为添加剂更合理的调控瘤胃发酵成为近年来的研究热点。本文基于植物单宁的分类及结构、微生物降解植物单宁的机制及单宁降解酶的相关知识,重点阐述植物单宁对瘤胃发酵的调控及甲烷排放等的影响,旨在为植物单宁作为一种潜在的绿色添加剂在调控瘤胃发酵及改善反刍动物机体健康方面提供新的方法和思路。
1 植物单宁及其微生物降解
1.1 植物单宁的分类及结构
作为天然存在的多酚类物质,植物单宁具有多种官能团,其单体单宁酸化学式为C76H52O46,可与蛋白质、少量碳水化合物和多糖等形成各种复杂化合物,因其结构不同,功能也存在差异。植物单宁通常包含水解单宁(hydrolysable tannins,HT)和缩合单宁(condensed tannins,CT)2种类型[9-10](图1)。水解单宁主要为邻苯三酚类[10],其相对分子质量较小,一般为500~3 000[11],容易被酸、碱和酶水解,进而形成没食子单宁和鞣花单宁。没食子单宁常见于豆荚、漆树叶、栓皮栎中[12],可进一步水解产生没食子酸和葡萄糖[13];鞣花单宁主要存在于合金欢、板栗、松树皮、橡木、黑莓、石榴中[12,14],水解可产生葡萄糖和六羟基二苯甲酸[15],其中六羟基二苯甲酸可形成鞣花酸等[16]。缩合单宁主要为邻苯二酚类,最常见的缩合单宁为原花青素(儿茶素和表儿茶素),其相对分子质量较大,一般为1 900~28 000[11],主要存在于豆科植物、乔木和灌木中[14]。缩合单宁亚基结合成低聚物和聚合物的方式主要是通过1个黄烷-3-醇单体C-环的C-4位共价连接到其他亚基C-环的C-6和C-8位,最终聚合成较稳定的C6·C3·C6结构,在胃肠道中不易被降解,酸性条件下会进一步缩合成花青素等物质[10-11,17]。过量的单宁可与蛋白质形成不易被消化的复合物,还可与消化酶结合改变消化酶的活性和功能,进而影响家畜的采食量和消化率[18]。近年来,研究发现反刍动物饲粮中添加适量的植物单宁会促进机体对进入瘤胃的营养物质的消化、吸收与利用[19]。
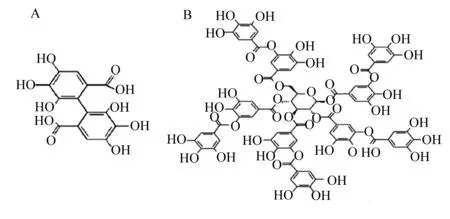
A:水解单宁 hydrolysable tannins;B:缩合单宁 condensed tannins。
1.2 可降解植物单宁的微生物
植物单宁对大多数微生物存在广谱抑制性,使植物单宁的微生物降解存在一定的困难,但自然界内仍有部分细菌、霉菌以及酵母菌等(表1)对植物单宁存在一定的耐受性,能够降解不同种类的植物单宁。此外,在反刍动物瘤胃内也发现了部分可降解植物单宁的微生物[20]。

表1 可降解水解单宁与缩合单宁的部分微生物
2 植物单宁的微生物降解机制
2.1 水解单宁
通常条件下,细菌对水解单宁的降解主要是通过其分泌的酶将水解单宁转化为没食子酸,没食子酸通过没食子酸脱羧酶或没食子酸羧化酶转化为邻苯三酚,然后在氧化酶的作用下氧化断裂为脂肪酸[27]。然而,由于反刍动物瘤胃内的严格厌氧环境,使得没食子酸降解途径发生改变,即邻苯三酚经异构酶转化为间苯三酚,再经还原酶和水解酶转化为3-羟基-5-氧己酸并在一系列酶的作用下先转化为乙酰辅酶A(acetyl-CoA),继而分解生成乙酸酯和丁酸酯[28]。此外,Krumholz等[29]在对水解单宁细菌降解的相关研究中发现,瘤胃中3-羟基-5-氧己酸酯(HOHN)由HOHN-辅酶A转移酶的酶促作用产生,瘤胃细菌通过β-羟基丁酰辅酶A脱氢酶、丁酰辅酶A脱氢酶、乙酰辅酶A转移酶、烯酰辅酶A水合酶、磷酸乙酰转移酶和乙酸激酶的顺序酶促作用转化为乙酸盐和丁酸盐。同时,Nelson等[30]发现一种新的瘤胃细菌(西塞利镰刀菌)可分解单宁酸和酚类单体,如没食子酸、连苯三酚、阿魏酸和对香豆酸,经验证此类化合物的降解途径与早期研究结果相似,即该类化合物单体首先经乙酰辅酶A还原,最后水解成六碳酮酸。此外,在反刍动物体内,水解单宁经瘤胃和肠道代谢可形成易吸收的小分子代谢物(邻苯三酚、间苯二酚和间苯三酚)和其他酚类代谢物(儿茶酚、尿石素和尿石素衍生物)[31-32]。然而,通过水解单宁的不完全生物转化产生的水解单宁代谢物(主要是邻苯三酚)对瘤胃微生物群落和宿主有不利影响[32]。
可降解水解单宁的真菌大多来源于青霉属和曲霉属。不同种属的霉菌对水解单宁的降解机制大体相同(在单宁酶的作用下,霉菌先将没食子单宁转化为没食子酸和葡萄糖),却又不完全一致,如烟曲霉可直接将没食子单宁转化为没食子酸;在黑曲霉中,没食子酸是可水解单宁的酚类单体,被加氧酶氧化裂解形成不稳定的三羧酸中间体(顺乌头酸、α-酮戊二酸和柠檬酸等),然后经氧化脱羧产物甲酰乌头酸脱羧,最终进入柠檬酸循环[33];而在黄曲霉中,没食子酸经三羧酸中间体降解为草酰乙酸,最后转化为丙酮酸[12]。
2.2 缩合单宁
早期研究发现,缩合单宁单体(黄酮类化合物)可被瘤胃液降解[34]。1999年McSweeney等[35]首次发现了以缩合单宁为单一碳源的瘤胃细菌,为之后缩合单宁的细菌降解提供了思路。2009年,Brown等[36]研究发现缩合单宁在厌氧环境中被消化道菌群转化为芳香中间体,之后生成甲烷。胡文斌等[37]指出瘤胃内黄酮类化合物的降解途径为:瘤胃微生物将栎精(黄酮醇)降解为间苯三酚和苯乙酸盐的衍生物,瘤胃内的厌氧条件促使间苯三酚快速降解,其产物进入三羧酸循环最终生成甲烷和二氧化碳。陈度宇等[38]利用梯度驯化法从牛、羊粪中分离出1株以缩合单宁为唯一碳源的革兰氏阴性菌,并通过正丁醇-盐酸法和液相色谱法检测缩合单宁降解率,72 h后缩合单宁降解率可达90%以上。该研究首次表明革兰氏阴性菌可以降解缩合单宁为儿茶素,为缩合单宁的细菌降解提供了新思路。
缩合单宁结构复杂,分子中的芳香环以C—C键相连,是黄烷-3-醇类单宁的聚合物,自身缺少易水解的没食子酰基结构,加上空间位阻效应的影响,在反刍动物胃肠道内很难被微生物降解[39]。但研究发现仍有部分真菌可以断裂缩合单宁分子中的C—C键,使其成为黄烷醇单体,然后进一步转化为酚类和酸类等小分子化合物[40-42]。
3 单宁降解酶
微生物降解植物单宁的实质就是微生物分泌的酶催化植物单宁降解的酶促反应过程[37]。研究发现,微生物能够分泌使植物单宁发生降解的单宁降解酶,如单宁酶、脱羧酶、多酚氧化酶等[31]。实际上,无论水解单宁还是缩合单宁,本身均含有大量的羧基和羟基等官能团,能够与蛋白质形成复合物,进而抑制微生物生长,阻碍微生物的代谢过程。单宁-蛋白质化合物的降解包含2个步骤:第1步是单宁-蛋白质化合物分子间的化学键受酶的作用率先断裂,形成游离的单宁;第2步是游离单宁分子中的酯键或缩酚键在单宁酶的作用下发生断裂,形成分子质量较小的缩酚酸和多元醇,缩酚酸与多元醇继续被多酚酶与脱羧酶等酶降解为间苯三酚和间苯二酚,最终转化为3-羟基-5-氧-己酸、5-氧-6-己酸甲酯等小分子物质[37,43]。
3.1 单宁酶
单宁酶也称单宁酰基水解酶(TAH),该酶能够催化没食子单宁等底物中酯键和缩酚键的水解,释放没食子酸和葡萄糖[44]。自然界内,动物、植物、细菌和真菌均可分泌单宁酶。其中,用于工业化生产的单宁酶大多源自于微生物[45]。早期研究发现,真菌、酵母菌等微生物均能分泌单宁酶,已被研究发现的可分泌单宁酶的真菌有根瘤菌(Rhizobium)、木霉菌(Trichodermaspp.)等[46];细菌有芽孢杆菌(Bacillus)、纤维单胞菌(Cellulomonas)、葡萄球菌(Staphylococcus)和肠杆菌(Enterobacteriaceae)等[47-48]。此外,山羊等反刍动物消化道内也分离到单宁降解菌[49]。研究表明,单宁酶的催化作用主要取决于酶-底物复合物(enzyme-substrate compound,ES)的形成[50]。同时,微生物分泌的单宁酶在水解单宁转化为没食子酸方面有巨大潜力,但是在反刍动物肠道内的具体作用机制尚不清楚[51]。
3.2 脱羧酶
微生物在降解单宁类化合物的过程中会产生脱羧酶,它可将没食子酸在脱羧作用下生成焦棓酸,因此也被称为没食子酸脱羧酶。研究发现,由反刍动物肠道内筛选的大肠杆菌、链球菌等具有较强的脱羧酶活性,可使棓酸脱羧成为焦棓酸,甚至可以直接脱去单宁分子酚环上的羧基形成焦棓酸[52]。比如,水解单宁在单宁酶作用下生成中间产物1,2,3,4,6-黄酰单宁和2,3,4,6-四酰单宁,然后单宁酶催化其分子内酯键断裂产生没食子酸和葡萄糖,葡萄糖进入三羧酸循环[53];没食子酸再通过脱羧酶的作用形成焦棓酸,进而被各类氧化酶还原为间苯三酚、间苯二酚,最终形成3-羟基-5-氧-己酸、5-氧-6-己酸甲酯[54](图2)。
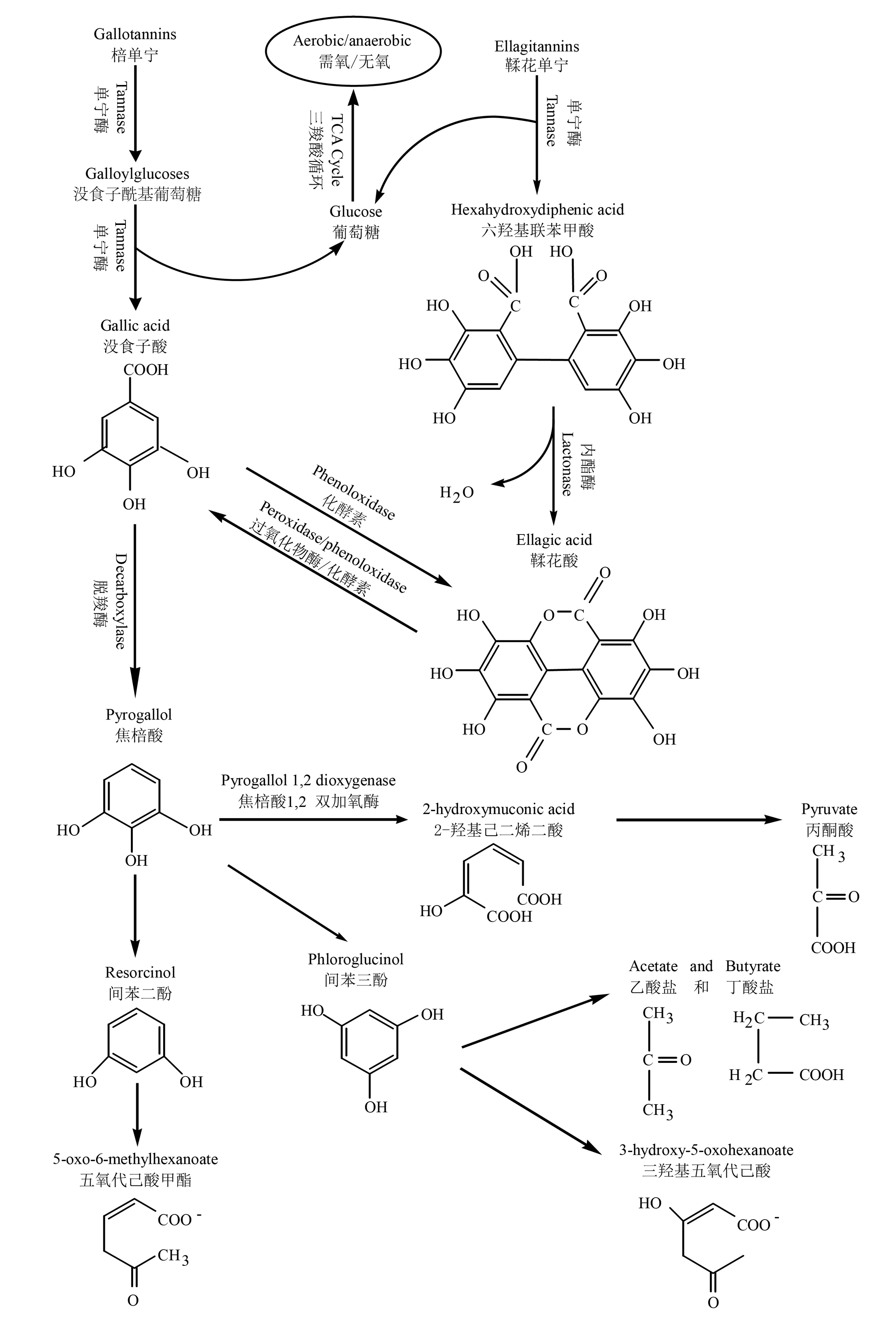
图2 没食子单宁、鞣花单宁的酶促降解过程
3.3 多酚氧化酶
多酚氧化酶是一类广泛存在于动物、植物、微生物中能够高效催化多酚类物质氧化的含铜质体的金属酶类[56]。多酚氧化酶又称儿茶酚氧化酶、邻苯二酚氧化还原酶,广义上可分为2类:1)单酚氧化酶(EC 1.14.18.1),即酪氨酸酶;2)双酚氧化酶(EC 1.10.3),包括儿茶酚氧化酶(EC 1.10.3.1)和漆酶(EC 1.10.3.2)[57]。植物中的多酚氧化酶一般指儿茶酚氧化酶[58]。有氧条件下,多酚氧化酶可通过2步将酚类物质转化为邻醌,其一是现有的羟基发生邻位羟基化;其二是邻二羟基苯氧化为邻苯醌[59-60]。
4 植物单宁对瘤胃发酵的调控
4.1 调节瘤胃微生物并提高饲料转化率
大量研究表明,植物单宁等植物多酚作为潜在的“替抗”饲料添加剂,可影响瘤胃微生物的生长、代谢和繁殖,进而在调控瘤胃发酵模式,减少甲烷排放,提高反刍动物对饲粮及营养物质消化率等方面表现出巨大潜力[61]。这可能与植物单宁可以选择性地影响特定的瘤胃细菌,从而改变瘤胃上皮中乙酸、丙酸等短链脂肪酸的吸收和代谢过程有关[62-63]。Díaz Carrasco等[64]指出饲粮中添加单宁降低了瘤胃微生物总丰富度,但没有显著影响细菌、古菌等类群相对丰度的平衡。Shabat等[65]发现饲粮中添加单宁会降低瘤胃微生物相对丰度,代谢物输出种类减少,而特定代谢物数量增多,进而提高了饲料转化效率(图3)。此外,饲粮中合适比例的单宁能够沉淀蛋白质,绕过瘤胃消化,增加过瘤胃蛋白的数量,提高小肠对于蛋白质的吸收率并降低瘤胃液中可溶性氮和氨态氮(NH3-N)浓度,增加宿主对于氮的利用率[64,66],进而减少能量损失。
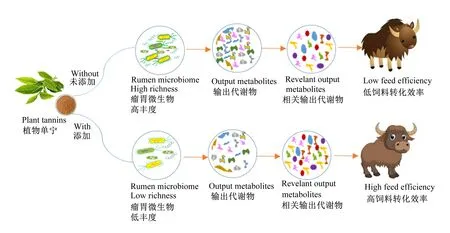
图3 植物单宁对瘤胃微生物丰度、代谢物及饲料转化效率的影响(参考Shabat等[65],略有调整)
4.2 调控瘤胃液pH、挥发性脂肪酸(VFA)和NH3-N浓度
瘤胃液pH是衡量瘤胃发酵的一个特征性指标,一般瘤胃液呈中性或弱酸性,瘤胃发酵的最适pH是6.0~7.0,pH低于或超出此范围均不利于瘤胃微生物的生长和瘤胃发酵等活动的进行[67],瘤胃液pH降低会抑制产甲烷菌的生长,降低纤维消化率,增加酸中毒的风险[68]。肖敏敏等[69]研究发现,饲粮添加2%的高粱单宁提取物对瘤胃液总挥发性脂肪酸(TVFA)浓度无显著影响。同样,Abdalla等[70]通过体外试验发现,在绵羊饲粮中添加3%的缩合单宁能够使瘤胃液中丙酸、异丁酸和异戊酸的比例显著降低,同时能够使乙酸和丙酸的比例显著提高,但是TVFA浓度无显著变化。杨凯[71]也发现,在肉牛饲粮中添加1.3%和2.6%的单宁酸均能够显著降低其瘤胃液内异戊酸和戊酸比例,同时显著提高丙酸比例,但是对瘤胃液pH和TVFA浓度无显著影响。但Bueno等[72]通过体外发酵试验指出,添加5%的缩合单宁会显著降低培养液TVFA浓度,但乙酸、丙酸和异戊酸等各组分比例无显著变化。因此,我们认为植物单宁添加量约为3%时不会对瘤胃液pH及TVFA浓度产生显著影响,但能够调整乙酸、丙酸、异丁酸和异戊酸等各组分的比例。王敬尧等[73]通过体外培养试验发现,随单宁酸浓度的升高,瘤胃液pH呈现下降的趋势,推测其原因可能是单宁酸抑制了瘤胃微生物的发酵,使VFA浓度降低,进而降低了pH。同时,王敬尧等[73]研究发现,添加单宁酸的试验组发酵6和24 h的NH3-N浓度显著低于对照组,且单宁酸添加量越高,NH3-N浓度下降越明显。由此,我们推测NH3-N浓度的下降可能与植物单宁抑制了瘤胃微生物的活性有关。潘发明等[74]指出,饲粮中单宁酸的添加量低于1.5%DM时能够显著提高氮的留存率,反之则会降低。总之,饲粮中添加适量的植物单宁可以抑制瘤胃微生物降解,增加过瘤胃蛋白的数量,提高机体对蛋白质的利用效率;减少可溶性氮和NH3-N浓度,提高反刍动物对于饲粮中必需氨基酸和氮的利用效率[65,75]。饲粮中植物单宁添加量的不同对瘤胃发酵的影响也不同(表2),呈现出剂量效应。因此,探索饲粮中植物单宁的适宜添加量,明晰植物单宁调控瘤胃发酵的作用机制十分关键。
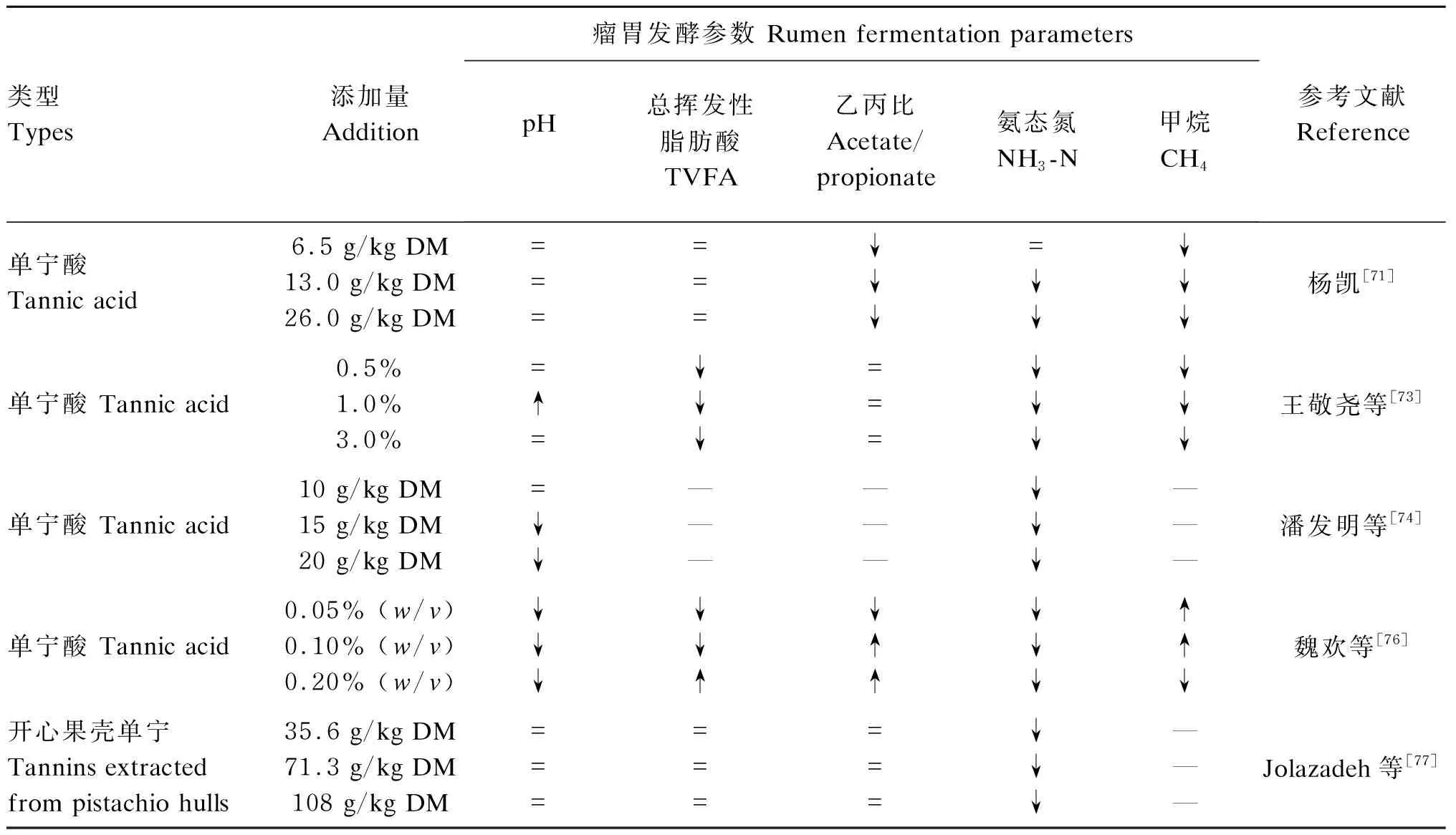
表2 植物单宁对瘤胃发酵参数的影响
4.3 对采食量及营养物质消化的剂量效应
与单胃动物不同,反刍动物特有的瘤胃是其消化饲粮中营养物质的主要场所。杨凯[71]研究发现,在饲粮中添加低浓度(0.65%、1.50%)的水解单宁对肉牛中性洗涤纤维(NDF)、酸性洗涤纤维(ADF)、粗蛋白质(CP)的表观消化率无显著影响,但随着水解单宁浓度的不断升高,肉牛对CP的表观消化率逐渐下降;当水解单宁浓度达到2.60%时,肉牛对干物质(DM)的表观消化率显著下降。Kamel等[78]研究发现,饲粮中添加0.3%的白坚木缩合单宁使得绵羊对NDF、ADF和CP的表观消化率显著降低。Kronberg等[79]通过试验也得到类似结论,随着饲粮中缩合单宁添加量的增加(0.7%、1.5%、2.9%),绵羊对DM、NDF、ADF、CP的表观消化率呈现下降的趋势。此外,缩合单宁会影响瘤胃蛋白质的消化并呈现出剂量效应,当饲粮中蛋白质含量较高而碳水化合物等其他营养物质含量较低时,缩合单宁会表现出有利的影响[80];而当饲粮中蛋白质含量不足时,缩合单宁的存在会降低反刍动物对蛋白质的消化率并对反刍动物的健康构成威胁[81]。潘发明等[74]指出,饲粮中单宁酸浓度低于1.5%时,能够提高对营养物质的消化率;而饲粮中单宁酸浓度高于1.5%时,绵羊采食量降低,对营养物质的消化率也会降低。因此,植物单宁对饲粮中营养物质吸收的促进或抑制作用与其浓度密切相关,植物单宁浓度过高导致其与蛋白质结合形成单宁-蛋白质复合物,从而降低反刍动物对饲粮中营养物质的吸收率。不同类型植物单宁及其添加量对干物质采食量及营养物质消化率的影响也不尽相同(表3)。
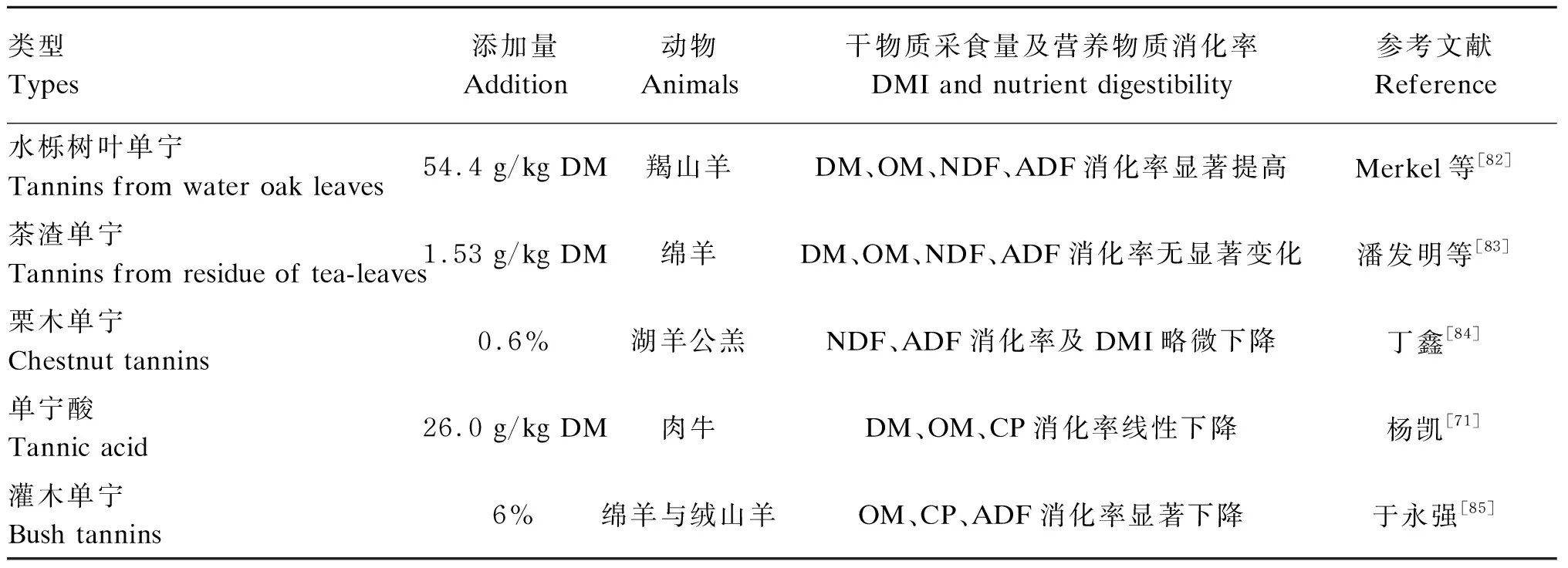
表3 植物单宁对干物质采食量及营养物质消化率的影响
4.4 抑制瘤胃甲烷排放
甲烷作为一种温室气体,其温室效应潜力约为二氧化碳的28倍,甲烷产量的增加对全球环境构成严重威胁。据统计,反刍动物每年的甲烷排放量约为8 000万t,可达全球甲烷排放总量的25%[86],占饲粮能量总损失的5%~9%[87]。大量研究表明,单宁等植物次生代谢物可以调控瘤胃发酵,提高动物生产力,减少甲烷排放等[88]。例如,丽丽等[89]通过研究不同添加量的柠条单宁对绵羊产甲烷菌多样性的影响指出,不同添加量的柠条单宁均显著降低了瘤胃产甲烷古菌群落的丰富度与多样性,这可能是由于单宁率先与产甲烷菌表面的蛋白质相结合,形成了单宁-蛋白质复合物,从而抑制产甲烷菌的生物活性,使得甲烷产量降低[90]。Grainger等[91]发现,在放牧奶牛中添加高水平缩合单宁(453 g/d)时,甲烷产量下降了14%,在放牧奶牛中添加较低水平缩合单宁(244 g/d)时,甲烷产量下降了29%。同样,Animut等[92]也指出缩合单宁可抑制甲烷排放,且抑制程度与缩合单宁浓度有很大的关系。此外,Saminathan等[93]通过体外试验指出,瘤胃甲烷产量会随着银合欢缩合单宁分子质量的增加而显著降低,但对干物质采食量的影响不显著。同时,Tapio等[94]报道称甲烷产量与原虫数量之间存在正相关关系,因此原虫数量的减少会抑制甲烷的生成。而Jolazadeh等[77]在荷斯坦公牛饲粮中添加7.13%的缩合单宁后发现,瘤胃中原虫数量减少。同样,丁学智[95]也发现,随着单宁浓度的增加,瘤胃中原虫数量呈现降低的趋势。出现以上结果的原因可能是植物单宁降低了瘤胃细菌的数量,间接影响了原虫生长所需的氮的供给,从而使得原虫的数量下降[96]。因此,植物单宁抑制瘤胃甲烷排放的另一个原因可能与植物单宁减少了瘤胃原虫数量有关。诸多学者对植物单宁影响瘤胃甲烷排放进行的大量的试验结果存在差异,可能与试验中使用的单宁种类、结构、相对分子质量及添加量的不同有关。总体而言,植物单宁能够在一定程度上抑制反刍动物瘤胃甲烷排放,是一种非常有潜力的甲烷抑制剂,但是其抑制瘤胃甲烷生成的作用机理与途径还亟待进一步深入挖掘。
5 小 结
综上所述,微生物降解植物单宁具有毒副作用小、残留量低等优点。随着合成生物学、基因工程、酶的定向突变技术等新型生物技术的兴起,未来利用微生物完成大分子植物单宁的生物降解,将成为植物单宁降解的主流途径。同时,随着饲料端“禁抗”的提出以及对植物单宁研究的不断深入,植物单宁作为一种潜在的新型“替抗”饲料添加剂在畜牧生产中的应用前景将十分广阔。但是,植物单宁调控瘤胃微生物区系与瘤胃发酵等瘤胃活动的具体作用机制尚不明确,在许多不同的试验研究中,由于植物单宁的结构、纯度、种类和添加量的不同,往往会呈现出不同的效果,当然,这也可能与试验动物的种类和饲养方式等有关。因此,今后应深入挖掘植物单宁作为瘤胃调控剂、甲烷抑制剂和饲料添加剂等的作用机制,使其更大程度的促进动物生产。
