基于微卫星分析的长丰鲢种质资源遗传监测
2022-06-09罗相忠覃维敏梁宏伟邹桂伟
罗相忠 覃维敏 梁宏伟 沙 航 邹桂伟
(中国水产科学研究院长江水产研究所, 武汉 430223)
鲢(Hypophthalmichthys molitrix)是我国重要的特有经济鱼类, 具食物链短、易饲养、成本低、调节水质等特性和功效。其以水体中浮游植物为主要饵料, 属滤食性鱼类, 为典型的碳汇渔业养殖对象[1]。鲢的营养价值较高, 必需氨基酸含量均略高于鲤(Cyprinus carpio); 二十碳五烯酸(EPA)和二十二碳六烯酸(DHA)的含量高于鳙(Aristichthys nobilis); 一直深受广大养殖户和消费者所青睐。鲢在大部分池塘、湖泊和水库中均有养殖或增殖, 是淡水养殖的当家品种之一。我国鲢养殖产量仅次于草鱼(Ctenopharyngodon idellus), 位居淡水养殖种类的第二位[2]。长丰鲢(Changfeng silver carp)(CF)是中国水产科学研究院长江水产研究所选育而成的鲢新品种(证书: GS-01-001-2010), 也是我国“四大家鱼”的第一个人工培育新品种, 具有生长快、遗传纯合度高、出肉率高和较耐低氧等优良特点, 其推广应用提高了我国鲢的良种覆盖率[3]。
长丰鲢自通过审定, 已连续繁育了3代, 即长丰鲢子一代(CF1)、长丰鲢子二代(CF2)和长丰鲢子三代(CF3)。其大多数亲本均由长丰鲢良种场和繁育基地连续繁育, 就此系统开展长丰鲢种质资源遗传监测和评估, 对其优良性状的保持具有重要意义。微卫星(SSR)因其稳定性高、多态性丰富、共显性遗传、位点分布广及分析成本低等特点被广泛应用于水产种质资源的遗传监测。唐首杰等[4]通过微卫星对团头鲂(Megalobrama amblycephala)3个选育群体进行的遗传纯度检测, 为纯化育种群体和监测种质质量提供了理论依据。李耀国等[5]采用微卫星对三亚海域卵形鲳鲹(Trachinotus ovatus)的养殖群体与野生群体进行了遗传多态性分析, 及基因型与性状的关联分析, 证实野生群体遗传多样性较养殖群体高; 微卫星位点中存在全长、体宽、体高和体质量的优势基因型。王丰等[6]用12个微卫星标记对长江4个野生青鱼(Mylopharyngodon piceus)群体和1个养殖群体进行了遗传多样性和结构分析, 5个群体均具有较高遗传多样性, 其中4个野生群体之间遗传距离较近, 且均与养殖群体之间遗传距离较远。此外, 还有对大口黑鲈(Micropterus salmoides)[7]、唇䱻(Hemibarbus labeo)[8]、大泷六线鱼(Hexagrammos otakii)[9]、团头鲂[10]、鲤[11]和大菱鲆(Scophthalmusmaximus)[12]等选育群体遗传监测的研究报道, 为评估相关选育群体的育种潜力和培育新品种(系)提供了支撑。本研究旨在利用微卫星标记分析长丰鲢连续不同世代群体的遗传多样性与遗传结构, 为其优良性状延续保持技术的建立与品种的持续升级换代提供理论依据。
1 材料与方法
1.1 实验材料
CF1亲本群体采集于河南省南阳市宛城区白河桥渔场长丰鲢良种繁育基地, 共采集21尾; 而CF2、CF3和L群体采集于湖北省浠水县长丰鲢良种场, 分别采集30尾。采集样本置于95%的乙醇中固定,-20℃冰箱中保存备用。样本基本信息见表 1。
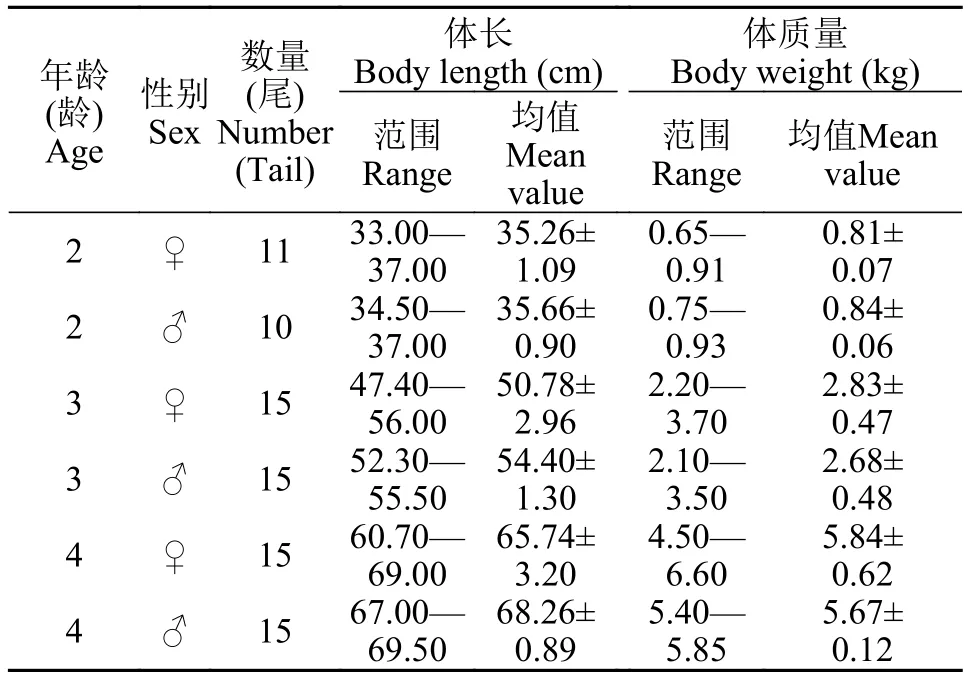
表1 长丰鲢各龄体长及体质量组成Tab. 1 Body length and body mass in each age group of Changfeng silver carp
1.2 DNA提取
采用高盐法进行样本基因组DNA提取, 经1%琼脂糖凝胶电泳检测后, 用无菌双蒸水调整浓度为50 ng/μL, -20℃保存备用。
1.3 微卫星引物
采用本团队自主开发的18对鲢微卫星引物进行分析, 荧光引物由武汉天一辉远生物公司合成,引物序列及退火温度如表 2所示。

表2 鲢卫星分子标记及其引物序列Tab. 2 Microsatellite markers and their primers sequences in silver carp
1.4 PCR扩增
PCR反应体系为20 μL, 其中模板DNA 1 μL (50 ng),上下游引物各0.5 μL, PCR mix 18 μL; PCR扩增程序为: 94℃预变性3min; 94℃变性30s, 退火(44—60℃)30s, 72℃延伸30s, 30个循环; 72℃延伸5min;4℃保存。
1.5 数据处理
采用荧光引物对4个群体样本进行扩增, 其产物经1.5%琼脂糖凝胶电泳检测后进行毛细管电泳;采用Popgene1.3.1软件分析等位基因数(Na)、有效等位基因数(Ne)、期望杂合度(He)和观测杂合度(Ho)及子代间的遗传相似性系数、遗传距离等遗传参数; 利用Cervus 3.0软件计算多态信息含量(Polymorphism information content,PIC); 近交系数(Fis)和群体间的遗传分化指数(Fst)及哈代-温伯格平衡由Arlequin 3.1.1分析软件统计获得; 群体间聚类分析图采用MEGA 5.0软件中UPGMA方法构建。
2 结果
2.1 鲢与长丰鲢各子代的遗传多样性分析
18个鲢微卫星位点在L、CF1、CF2和CF3群体中检测到等位基因数分别为130、103、93和91个(表 3)。L、CF1、CF2和CF3的平均等位基因数(Na)分别为7.2222、5.7222、5.1667和5.0556, 平均有效等位基因数(Ne)分别为4.3122、3.2551、3.2274和3.1461(表 3); 鲢平均等位基因数和平均有效等位基因数均比长丰鲢后代高, 其在长丰鲢后代中呈逐代下降趋势。各微卫星位点Na和Ne见表 3, 在鲢L群体中Na和Ne最高的是位点BL5; 在长丰鲢子一代(CF1)群体中Na和Ne最高的分别是位点BL109和位点BL5; 在长丰鲢子二代(CF2)群体中Na最高的是位点BL5和位点BL109, 而Ne最高的是位点BL5; 在长丰鲢子三代(CF3)群体中Na最高的是位点BL5、BL55和BL109,Ne最高的是位点BL5。
2.2 群体的杂合度
鲢观测杂合度为0.2667—1.0000, 平均观测杂合度为0.7190, 期望杂合度为0.2350—0.8994, 平均期望杂合度为0.7260; 长丰鲢子一代(CF1)平均观测杂合度为0.6975, 平均期望杂合度为0.6422; 长丰鲢子二代(CF2)平均观测杂合度为0.6111, 平均期望杂合度为0.6353; 长丰鲢子三代(CF3)平均观测杂合度为0.5407, 平均期望杂合度为0.6235(表 3)。4个群体中平均观测杂合度和期望杂合度: 鲢>长丰鲢子一代>长丰鲢子二代>长丰鲢子三代; 鲢杂合度较长丰鲢高, 不同群体在同一位点杂合度也有所不同, 同一位点在同一种群中观测杂合度和期望杂合度相差较大, 如位点S92和位点H121; 可能是该位点上的无效等位基因所致; 若无效等位基因不被识别, PCR扩增时会导致纯合子过剩或杂合子缺失,从而导致Ho和He出现偏差。
2.3 多态信息含量(PIC)
4个群体多态信息含量(PIC)为0.0320—0.8730(表 3), 鲢的平均多态信息含量(PIC)高于长丰鲢三个子代, 而长丰鲢各子代间平均PIC呈现逐代下降趋势。4个群体多态信息含量由高到低依次为: 鲢(0.6748)>长丰鲢子一代(0.5784)>长丰鲢子二代(0.5730)>长丰鲢子三代(0.5609); 鲢群体中PIC最高位点是BL109, 长丰鲢各子代群体中PIC最高位点都是BL5。
2.4 Hardy-Weinberg平衡分析
如表 4所示, 位点H121和BL55的d值在4个群体中均为负值, 表明4个群体在这2个基因位点上可能存在杂合子缺失。在鲢群体中有8个位点的d值为负数, 而在长丰鲢子一代、子二代和子三代中d值为负数的位点数量分别为5、11和13个。4个群体平均d值分别为0.0044、0.0681、-0.0345、-0.1178,其中鲢和长丰鲢子一代的d值为正数, 长丰鲢子二代与子三代2个群体的d值为负数。这说明后2个群体都可能存在杂合子缺失情况, 且长丰鲢子三代杂合子缺失最为严重。如表 3所示, 位点H121、H129和S78在4个群体中都偏离了Hardy-Weinberg平衡状态(P<0.05)。4个群体平均Hardy-Weinberg平衡P值分别为0.3096、0.4230、0.2844和0.1635,P值均大于0.05, 4个群体都处于Hardy-Weinberg平衡状态。

表3 18个位点在鲢和长丰鲢4个群体中的遗传多样性Tab. 3 Genetic diversity of four populations of silver carp and Changfeng silver carp based on 18 polymorphic microsatellite loci
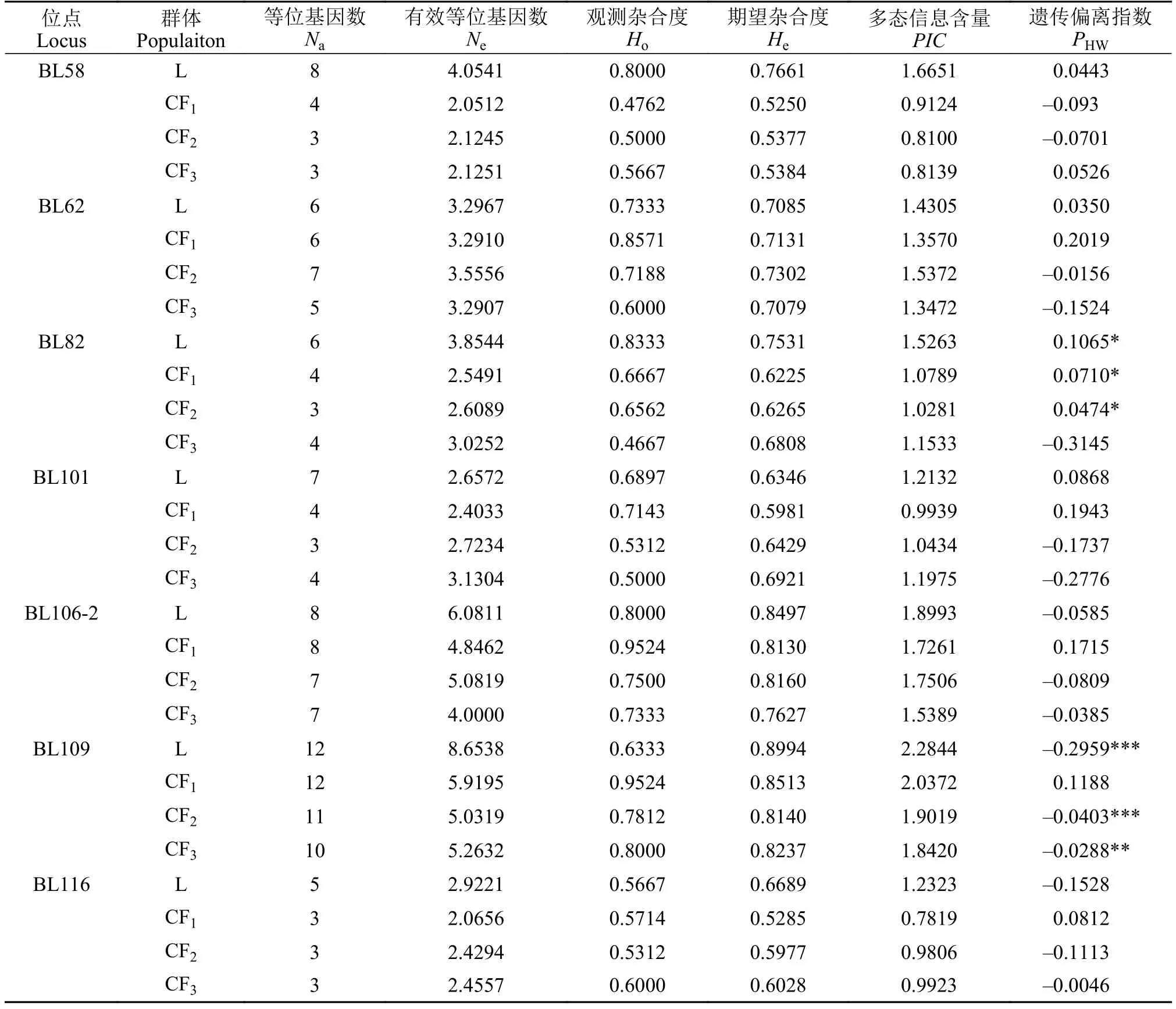
续表3
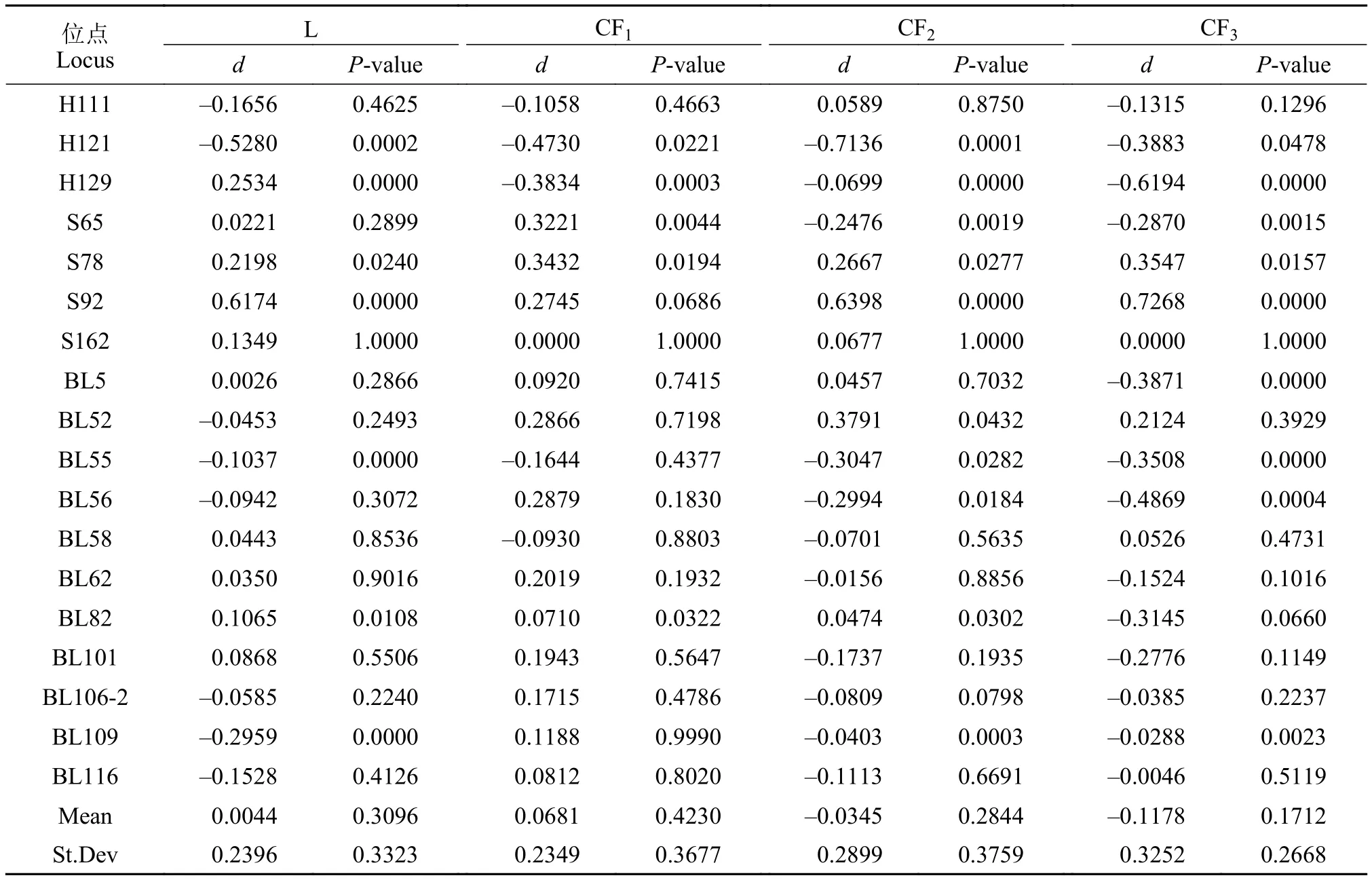
表4 Hardy-Weinberg平衡检测d值及P值Tab. 4 The d and P value of each locus by Hardy-Weinberg test
2.5 固定指数
鲢与长丰鲢子一代、子二代和子三代平均近交系数Fis分别为-0.057、-0.0017和0.0372(表 5);鲢与长丰鲢后代间遗传分化系数Fst分别为0.0319、0.0301和0.0352(表 6); 4个群体间Fst最大的是鲢与长丰鲢子三代, 为3.52%(表 6)。在长丰鲢后代中,CF1与CF2间遗传分化指数值0.0164, CF1与CF3间遗传分化指数值0.0286, CF2与CF3间遗传分化指数值0.0176; 其中CF1与CF2间遗传分化指数最小, 其遗传变异主要是来自个体之间; 鲢与长丰鲢后代之间遗传分化指数值(Fst)较高, 而长丰鲢各后代之间遗传分化很低。
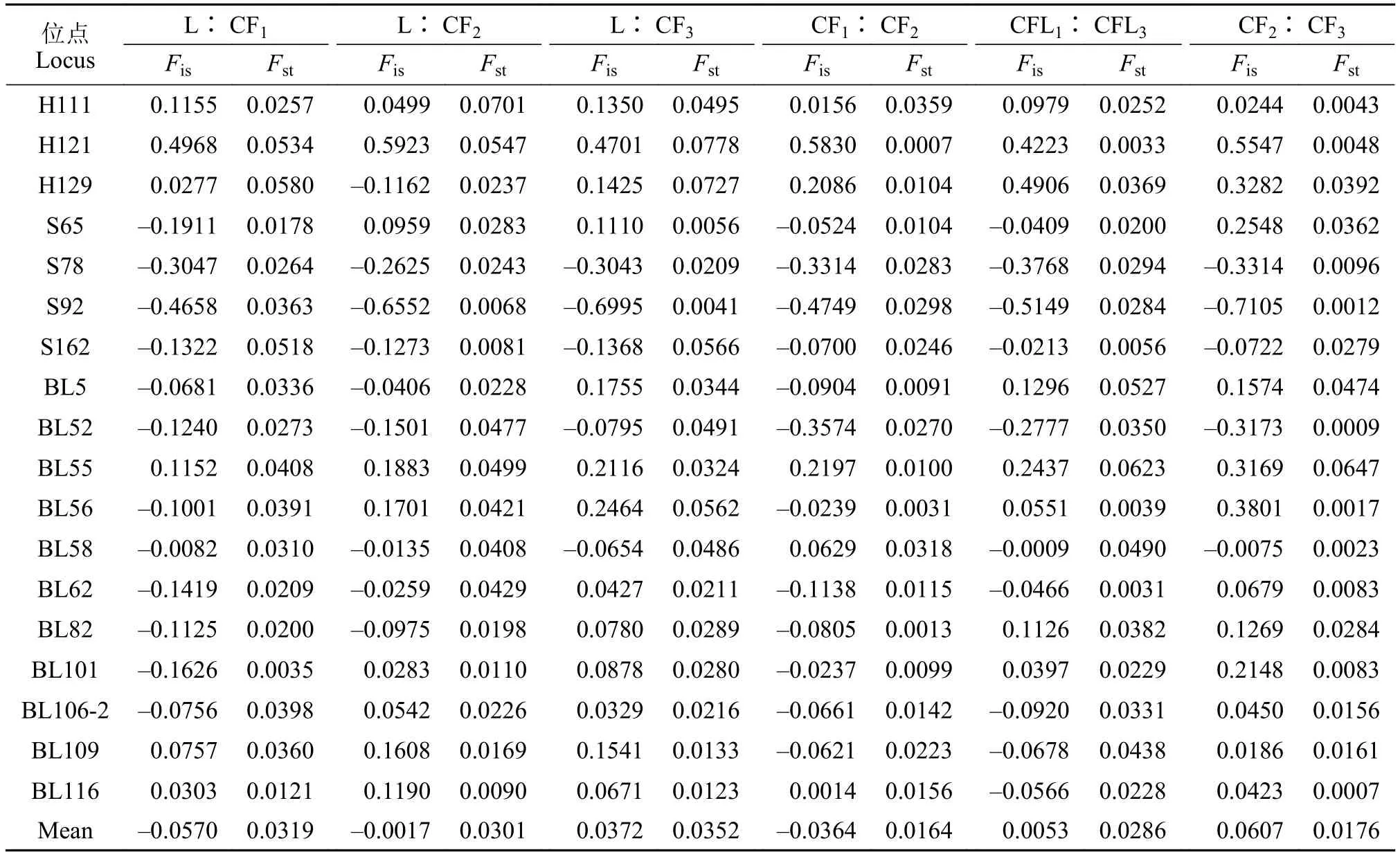
表5 鲢、长丰鲢各世代间Fis和Fst值比较Tab. 5 Comparison of Fis and Fst values among different generations of silver carp and Changfeng silver carp
长丰鲢子代群体有一定程度观测杂合度过剩现象, 子一代、二代和三代群体杂合子过剩位点数分别为13、7和6个; 鲢群体中近交系数Fis最高位点是H121(0.5200; 表 7), 该位点在群体内遗传变异程度低, 遗传多样性也低; 4个群体微卫星位点中BL55遗传分化(Fst)值最大(0.0642), BL116值最小(0.0178); 平均值为0.0406; 遗传分化程度较弱的位点有13个, 中等的位点 5个; 而在长丰鲢后代中遗传分化(Fst)平均值为0.0282, 仅BL55遗传分化程度中等, 其他位点都程度较弱, H121和BL56分化程度最低为0.0039(表 7); 长丰鲢后代在检测的18个多态位点上都表现出遗传分化不明显, 处于低等水平。
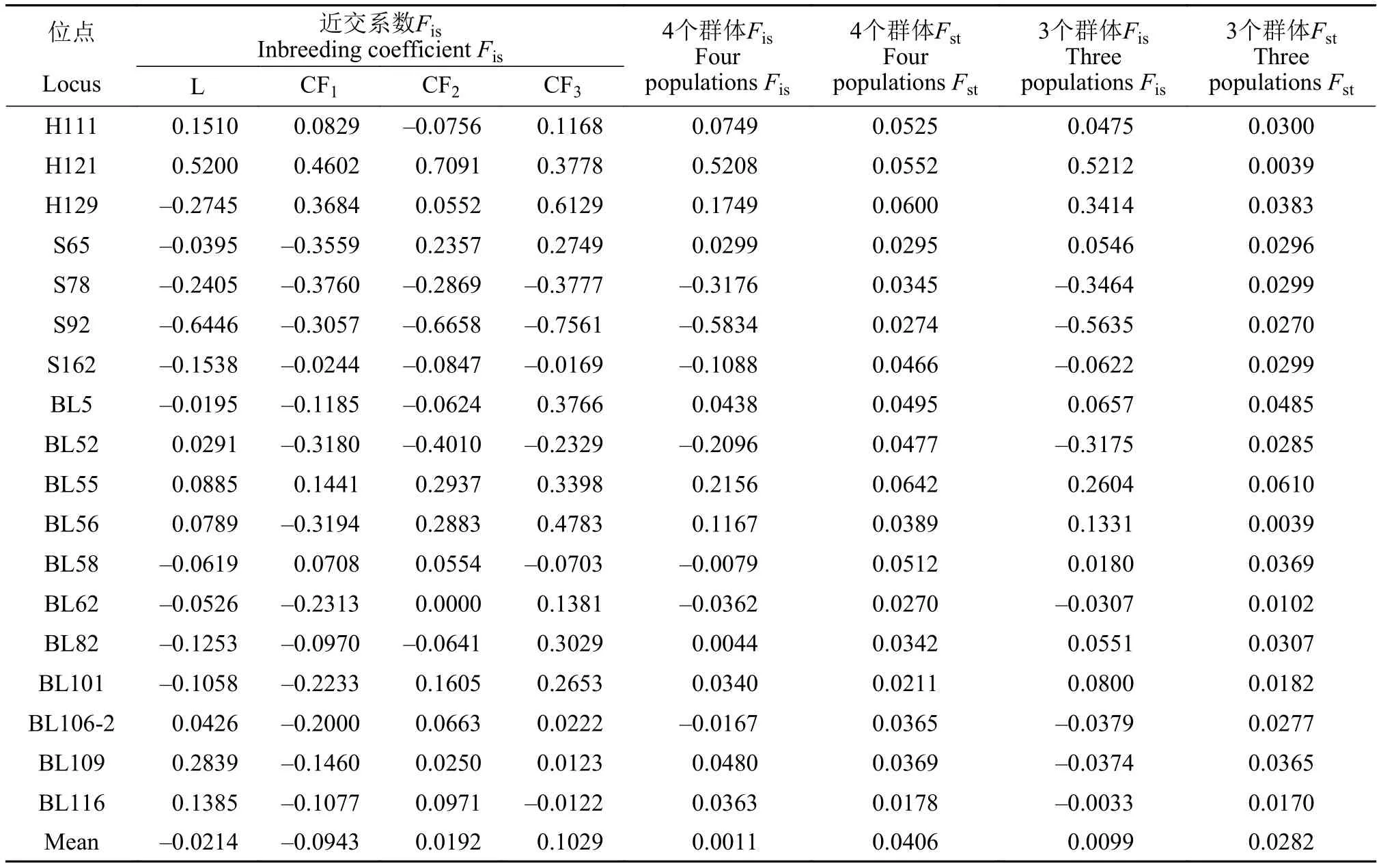
表7 鲢、长丰鲢各世代18对微卫星位点的F-检验Tab. 7 F-statistics for different generations of silver carp and Changfeng silver carp at 18 microsatellite loci
2.6 遗传距离及遗传相似性系数
4个群体间遗传距离为0.0299—0.0934(表 8),鲢与长丰鲢三个世代遗传距离最远(0.1174), 与CF2的遗传距离为0.1115, 与CF1的遗传距离为0.1114, 鲢与长丰鲢子一代、子二代和子三代之间遗传距离逐渐上升; 而在长丰鲢子代之间, 子一代与子二代之间遗传距离为0.0392, 与子三代之间遗传距离为0.0951; 子二代和子三代遗传距离为0.0544, 其后代遗传距离有上升趋势; 鲢与长丰鲢各子代之间遗传相似性系数为0.8892—0.8946, 群体间遗传相似度较高, 但长丰鲢后代群体间遗传相似性系数则更高, 其中CF1与CF2之间遗传相似度最高(0.9616), 而CF1与CF3相对较低(0.9093)。依据4个群体间遗传距离值, 采用UPGMA法构建聚类图(图 1), 长丰鲢CF1群体最先与CF2汇成一支, 接着与CF3汇成一簇, 最后与鲢聚在一起。
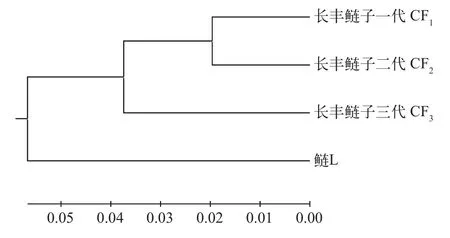
图1 鲢、长丰鲢共4个群体基于Nei氏无偏遗传距离的UPGMA进化树Fig. 1 An unweighted pair group method with arithmetic mean(UPGMA) dendrogram for 4 populations of silver carp and Changfeng silver carp based on Nei’s unbiased genetic distance
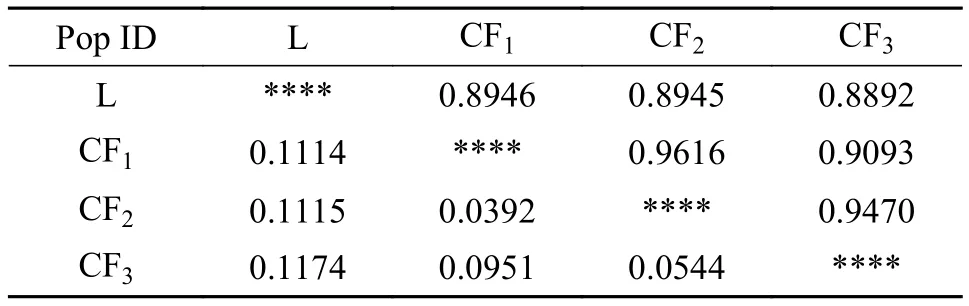
表8 鲢、长丰鲢共4个群体的Nei氏遗传距离(下三角)及遗传相似性系数(上三角)Tab. 8 Nei’s standard genetic distance (below) and genetic identity (above) between 4 populations of silver carp and Changfeng silver carp
3 讨论
3.1 群体遗传多样性
生物遗传多样性能反映一个种群的进化、演化历程, 是物种长期适应环境和生存的基础。遗传多样性的高低是衡量生物对环境适应能力的重要指标。遗传多样性越高, 生物对环境的适应性就越强; 生物越易存活下来, 适应新环境并繁衍后代[13]。遗传多样性也是衡量物种遗传潜力的重要指标。遗传潜力与生物遗传多样性多呈正相关; 潜力越大, 遗传多样性也越丰富。在本研究中鲢群体等位基因数和有效等位基因数均高于长丰鲢群体, 主要是因为长丰鲢由长江水系的原始亲本雌性个体经雌核发育等技术选育而来。在雌核发育的草鱼和团头鲂后代中也发现其平均等位基因数低于普通群体[14,15]。在长丰鲢后代中, 不同子代间等位基因数(Na)和有效等位基因数(Ne)都呈逐渐下降的趋势。随着人工繁育世代的增加, 群体间近交程度加重, 群体中会存在等位基因丢失现象。与此同时,也会剔除一些非目标性状基因, 稳定目标性状基因,达到相关基因进一步纯合, 遗传多样性和育种潜力也降低。在长丰鲢后代中平均等位基因和有效等位基因均比叶香尘等[16]检测的长丰鲢等位基因数要多, 可能是由于检测样本或检测方法不同造成的。在通常情况下, 检测等位基因数和研究样本量的大小有关[17]。与传统聚丙烯酰胺凝胶电泳相比,基因分型技术有更高的分辨率, 可分辨出2—4 bp的等位基因[18], 极大地提高检测的准确性和检测效率。
杂合度是评估遗传多样性的重要指标之一。期望杂合度是评价群体生物多样性的最适参数, 能直接反映群体遗传多样性。期望杂合度He高低与生物适应性呈正相关。一个物种的期望杂合度越高, 适应环境能力越强, 生存机会亦越大[19]。在本研究中4个群体的观测杂合度和期望杂合度均为鲢最高, 长丰鲢子三代最低。在长丰鲢3个群体中, 期望杂合度区间为0.6235—0.7260, 均高于叶香尘等[16]检测的广西养殖长丰鲢(0.4133), 且低于检测的长江三峡库区4个群体鲢(0.7500—0.7943)[20]。总体上, 在长丰鲢3个子代中, 期望杂合度呈逐渐下降趋势, 这与大口黑鲈世代选育群体逐代下降变化趋势一致[7]。在长丰鲢3个子代间, 期望杂合度相差不大, 从子一代到子三代减少了3.00%。观测杂合度子三代较子一代减少了29.00%。与张天时等[21]在中国对虾(Fenneropenaeus chinensis)发现的观测杂合度下降3.5%相比, 降比更大。长丰鲢随着繁育子代的增加, 群体纯合度增加。与此同时, 长丰鲢3个子代群体期望杂合度均较高, 群体尚未出现明显的近交及瓶颈效应, 群体遗传多样性保持较好, 其具备持续选育潜力。
根据Botstein等[22]对微卫星位点多态性的定义,当PIC<0.25时, 该位点多态性低;PIC值在0.25—0.50, 表明其具有中度多态性;PIC>0.50时,该位点多态性高。鲢及长丰鲢子一代、二代和三代4个群体平均PIC值分别为0.6748、0.5784、0.5730和0.5609, 表明4个群体遗传多样性都较高, 但长丰鲢群体间呈逐代下降趋势。基于Hardy-Weinberg平衡检验结果, 4个群体平均d值分别为0.0044、0.0681、-0.0345和-0.1178。d值为负数, 表明长丰鲢子二代和子三代群体杂合子缺失严重, 在中国对虾、南方拟䱗(Pseudohemiculter dispar)、斑点叉尾鮰(Ictalurus punctatus)、团头鲂和德国镜鲤(Cyprinus carpioL.)等群体的研究中也发现类似现象[21,23—26]。杂合子缺失可能是无效等位基因存在或在聚合酶链式反应中出现丢失现象所引起, 但也可能与选育群体被高强度选择有关[27]。或者是在人工繁殖时群体雌雄比例、配组繁殖不合理导致近亲繁殖, 也可能因检测样本数太少引起。在长丰鲢后代亲本培育时受人为干扰严重, 亦会导致杂合子缺失。平衡偏离会造成一些不利影响。如群体内等位基因丢失, 导致纯合子比例上升, 种群遗传多样性随之降低,近交衰退几率也会上升, 最终可能会导致整个种群衰退。在今后应持续监测关注群体遗传多样性。
3.2 群体遗传变异分析
生物对环境适应性改变是其生存和繁衍的基础, 群体遗传变异程度是衡量一个群体存在选育潜力和种质资源可持续利用的前提。18个鲢微卫星位点中有10个位点近交系数Fis值为正值, 8个位点为负值, 平均Fis值0.0119。3个长丰鲢子代间存在轻度的近交现象, 3个群体Fis值分别为-0.0943、0.0229和0.1029, 世代间近交程度逐渐加重。今后在人工繁殖中应尽可能扩大繁育亲本群体, 制定科学合理的繁育计划。遗传分化指数(Fst)是衡量群体间遗传分化程度的重要参数。McConnell S等[28]认为,Fst值在0.00—0.05, 群体间遗传分化较小; 在0.05—0.15, 存在中等程度遗传分化; 在0.15—0.25,群体间遗传分化较大; 在0.25以上, 群体间遗传分化明显。长丰鲢后代群体中各位点Fst值在0.0039—0.0610, 群体平均遗传分化指数为0.0282, 群体遗传分化很小。此研究结果与李镕等[7]研究大口黑鲈世代群体总遗传分化指数为0.013相近。这表明长丰鲢后代遗传结构基本一致, 虽出现一定的遗传分化,但分化程度较低。长丰鲢子一代与子二代Fst值最小为0.0164, 子二代与子三代Fst为0.0176, 子一代与子三代的遗传分化指数Fst最大(0.0286)。随着选育进程的不断开展, 世代间遗传分化系数在相邻世代之间相继减小, 亦可能会进一步增大[29]。长丰鲢不同世代间遗传变异大部分来自群体内, 群体间遗传分化程度很低。长丰鲢世代间遗传距离均较小,说明其遗传相似性变高。就目前而言, 虽然长丰鲢群体间遗传多样性逐代降低, 但遗传多样性水平还维持在较高水平, 长丰鲢不同世代间遗传结构趋于稳定。
