基于稳定同位素技术的保安湖食物网结构特征研究
2022-06-09陈俊伊郭钰伦祝孔豪巴爱荣赵康顺
陈俊伊 王 康 郭钰伦 祝孔豪 巴爱荣 刘 云* 赵康顺 *
(1. 北京农学院生物与资源环境学院, 农业农村部华北都市农业重点实验室, 北京 102206; 2. 中国环境科学研究院, 湖泊水污染治理与生态修复技术国家工程实验室, 国家环境保护湖泊污染控制重点实验室, 北京 100012;3. 中国科学院水生生物研究所, 武汉 430072; 4. 中国科学院大学, 北京 100049)
食物网是生态系统中多种生物及其营养关系的网络, 它描述了生物群落内不同生物体之间复杂的营养相互作用, 并揭示整个生态系统水平上的物质循环和能量流动过程[1,2], 是生态系统中物质和能量流动的重要形式[1,3]。食物网研究可以指示群落的组成、结构及物种之间复杂的摄食关系, 对探究生态系统中能量流动和物质循环具有重要意义[4,5]。近年来, 生态系统水平的研究强调将食物网作为理解水域生态系统结构和功能的核心问题[6,7]。湖泊是内陆水体供水的主要部分, 其作为地球表面陆地水圈的重要组成部分及许多大河流域沿岸地区最重要的湿地类型, 在改善生态环境和维持生态系统健康等方面具有重要作用[8,9]。因此, 研究湖泊食物网对于维持生物多样性、保护水生态环境有重要作用, 并能为基于生态系统的生态保护和修复提供决策依据。
水域生态系统中物质的循环和能量的流动过程一直是生态学研究中的热点问题。早期对水域生态中食物网结构和生物营养级的研究多采用胃含物分析法, 该方法优点是较直观, 但存在许多缺陷, 如胃含物中主要反映的是动物短期摄食的结果,通常残留的是难以被消化的食物, 同时在研究小型动物方面存在困难, 存在较大的不确定性[6,10—12]。近年来, 稳定同位素技术作为生态学中兴起的一项新型研究手段, 它从新的角度为研究物种间营养关系提供了量化指标。测定生物体内天然存在的碳、氮稳定同位素比值(δ13C和δ15N)的变化可反映动物长期摄食吸收的食物来源、营养级和食物网结构[6,13,14]。应用碳氮稳定性同位素技术对水域生态系统食物网结构进行分析的研究在国外已广泛开展, 国内的研究也已起步, 但关于水域生态系统食物网的研究尤其是湖泊食物网结构特征的研究还很少。
保安湖是长江中游江汉湖群的一个典型浅水草型湖泊[15], 也是典型的江湖阻隔湖泊。随着湖区沿岸人口的增加以及渔业利用强度的加大, 可能对湖泊生态系统产生一定的干扰, 生态系统结构和功能产生不同程度的改变。针对保安湖面临的水生态与资源衰退等问题, 本研究应用碳、氮稳定同位素技术结合理论鱼类食性数据分析与同位素模型预测的方法, 同步构建保安湖的食物网结构, 量化物质流动和能量传递主要途径, 以期为长江中下游江湖阻隔湖泊生态系统修复、鱼类群落结构重建与渔业科学管理提供基础理论支持。
1 材料与方法
1.1 研究区域概况
保安湖(30°12′—30°18′N, 114°39′—114°46′E)位于湖北省大冶市西北部, 地处长江中游江汉平原东部边缘, 原与三山湖连成一片, 现以北练山、南练山为界, 山以东称为三山湖, 以西称为保安湖, 属梁子湖水系, 为长江中游南岸的一个中富营养型湖泊。保安湖湖水表面积约39 km2, 平均水深1.5—2.5 m, 最大水深3.7 m, 常年水位在17.5 m。湖面由扁担塘、肖四海、桥墩湖和主体湖组成, 其中肖四海被围堤拦断, 与主体湖水不再相通, 桥墩湖与主体湖间也筑起了透水坝, 但仍有水道与主体湖保持水联通[15,16]。
1.2 样品采集与测定
本研究于2019年6月在保安湖主湖区、扁担塘和肖四海开展了采样工作。鱼类样品主要是通过在沿岸带和敞水区设置网具获得, 其中扁担塘设置3个样点, 肖四海3个和主湖区6个, 共计12个鱼类采集点位, 每个样点设置多网目复合刺网一组(沉网和浮网各1条), 迷魂阵1个。鱼类样品取其背部肌肉, 去除骨刺, 然后放入离心管密封冷冻保存, 带回实验室后60℃烘干至恒重, 利用珠磨式组织研磨器(MiniBeadbeater-16)研磨粉碎, 干燥保存。用彼德森采泥器采集沉积物样品和底栖动物样品, 底栖动物种类鉴定后被放置于蒸馏水中过夜, 让其排空消化道内残留物。为了获得足够的样品分析质量, 同一物种的底栖动物被混合在一起, 烘干磨碎后干燥保存。底栖动物蚌和螺去壳取肌肉, 60℃烘干至恒重, 研磨粉碎后干燥保存待测。用于δ15N分析的沉积物样品使用1 mol/L 盐酸酸化去除碳酸盐的影响[17]。植物样品采集后洗净烘干, 研碎待测。在沿岸带采集上中下层水混合后用预烧的GF/C滤膜过滤水样获得有机颗粒物Particulate organic matter(POM)样品, 使用1 mol/L盐酸酸化去除碳酸盐的影响, 后用蒸馏水冲洗烘干, 充分磨碎。使用13#和25#浮游生物网收集浮游生物, 并将浮游生物样品过滤至预烧的GF/C滤膜, 使用1 mol/L 盐酸酸化去除碳酸盐的影响, 用蒸馏水冲洗后烘干磨碎。丝状附着藻经过蒸馏水清洗, 非丝状藻分离泥沙后过滤至预烧的GF/C滤膜, 烘干磨碎。
所有样品利用稳定同位素比质谱仪(CE公司的CarloErba NC 2500元素分析仪与Thermo fisher公司的Delta Plus质谱仪联用)测得其碳、氮稳定同位素比值δ13C和δ15N。分析碳(δ13C)、氮(δ15N)同位素的参照物质分别是VPDB (Pee Dee Belemnite)和空气中N2, 标准物质分别是国际上通用的IAEA-USGS24和IAEA-USGS26。
1.3 数据分析
根据Zhang等[18—22]总结分析的长江中下游鱼类食性数据, 基于社会网络分析(Social Network Analysis, SNA)原理[23—26]构建全湖捕食者的食性矩阵与理论食物网。为了能够更真实地反应保安湖的食物网结构与特征, 本文还采用Kadoya等[27]开发的贝叶斯同位素混合模型(Bayesian Isotope Mixing Model, BIMM)结合测量的实际样品同位素值进行全湖捕食者的食性预测分析[27]与食物网构建, 将基于理论食性数据构建的食物网与基于同位素混合模型预测的食物网进行比对分析。其中, 贝叶斯同位素混合模型结构如下:
一般情况下, 一个目标食物网的消费者i利用Mi资源,J元素的稳定同位素比率的N测量值取自每个消费者和每个食物来源:

式中,αi1,···,αiMi是Dirichlet先验参数, Λj和为食物网中各营养链中元素j营养富集因子先验分布的均值和方差, 营养富集因子在营养链和食物网中有很大差异, 但近似地遵循正态分布[3,27]。模型的准确性则通过比对初始食物网样品的同位素值与模型预测值之间的差异来判断。BIMM模型拟合使用R语言中的R2WinBUGS包[27,28]。最后, 使用cultevo包[29]中的Mantel检验分析理论食性数据构建的食性矩阵与基于BIMM模型预测的食性矩阵之间的相关性。所有数据处理、分析与图形制作均在R语言中进行。
2 结果
2.1 碳、氮稳定同位素特征
如图 1所示, 各食物源的δ13C值范围跨度较大,数值范围从-36.8‰到-8.5‰, 其中附着藻的δ13C值最高为-8.5‰, 浮游植物的δ13C值最低是-36.8‰。相对于δ13C值而言, 食物源的δ15N值变化范围较小,从-2.3‰到8.2‰, 以底栖动物最高是8.2‰, 沉积物最低是-2.3‰。此次共采集到16种鱼类, 鱼类的碳、氮稳定同位素测定结果如图 1所示。不同鱼类间的δ13C和δ15N值差异较大,δ13C值范围为-27.6‰到-14.7‰, 草鱼(Ctenopharyngodon idellus)的δ13C值较大;δ15N值变化范围为5.1‰到15.1‰, 湖鲚(Coilia ectenes taihuensis)、翘嘴鲌(Culter alburnus)和鳜(Siniperca chuatsi)的δ15N值较大, 草鱼(Ctenopharyngodon idellus)的δ15N值最低。
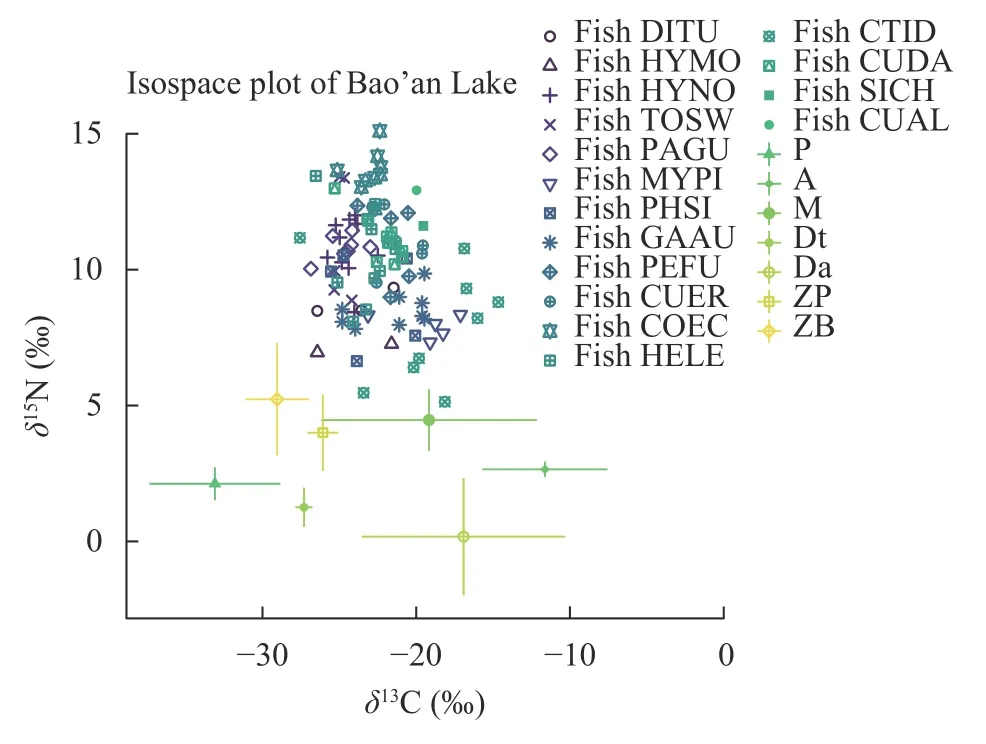
图1 保安湖水生态系统碳、氮稳定同位素混合空间Fig. 1 δ13C and δ15N of Bao’an Lake Ecosystem
2.2 理论食性网络
依据Zhang等[18—22]总结分析的长江中下游湖泊鱼类食性数据, 我们构建了保安湖食物网理论食性矩阵与依赖性矩阵(图 2)。从食性矩阵可以看出,POM、浮游植物、浮游动物与底栖动物是保安湖食物网中鱼类的主要食物来源。从食物依赖性矩阵中也可以看出保安湖食物网中捕食者对于POM、浮游植物、浮游动物与底栖动物的依赖程度较高,且对POM和浮游植物的依赖性最高, 底栖动物和浮游动物的被依赖性相对较低。
在理论食性矩阵的基础上, 我们构建了基于食性矩阵的保安湖食物网(图 3)。在构建的食物网络关系中, 我们重点关注主要的三个中心度指标: 点度中心度(Degree centrality), 为每个行动者所直接连接的其他行动者的数量; 中间中心度(Betweenness centrality), 衡量的是一个行动者在他相连的行动者之间所起到的影响和控制(桥梁)作用; 接近中心度(Closeness centrality)是计算某个行动者与他所有相连的行动者之间最短路径的总和[30—32]。从图 3的三个中心度指标和一个权重指标可以看出, 虽然鳜与翘嘴鲌的点度中心度较高, 但是权重较低, 说明这两种鱼捕食的鱼类种类较多, 但在整个网络中的重要性不高。而POM、浮游植物、浮游动物与底栖动物在整个食物网中的重要性与影响力相对较大。圆尾鲴(Distoechodon tumirostris)与底栖动物的中间中心度较高, 表明在食物网中路径间的媒介能力较强。因此, 结合图 3的食物网结构图可以发现, 基于理论食性分析数据构建的保安湖食物网由两条营养传递途径组成, 即由POM、浮游植物、浮游动物到杂食性鱼类的浮游牧食链与水生植物、沉积物和底栖动物到杂食性鱼类的底栖食物链, 浮游牧食链相对较强。
2.3 基于BIMM模型的食性网络预测
将原始的样品同位素实测值带入到贝叶斯同位素混合模型(BIMM)中进行食物网内各组分的食性预测分析, 并构建食性矩阵与食物网。首先, 通过图 4可以看出, 模型的碳、氮同位素预测值与实际样品的测量值差异很小, 数据重合度高, 说明模型的预测效果较好。我们将理论食性矩阵与基于贝叶斯同位素混合模型预测的食性矩阵进行矩阵间相关性分析, 两个食性矩阵具有显著相关性(P<0.001; 图 5), 表明贝叶斯同位素混合模型预测的食物网中各消费者的食性与理论食性具有很高的一致性。其次, BIMM模型的预测食性矩阵表明POM、浮游植物、浮游动物与底栖动物是保安湖水域食物网中鱼类的主要食物来源, 其次是沉积物中的碎屑和水生植物等, 这与理论食性矩阵一致(图 2和图 6A)。最后, 从BIMM模型预测的食物网结构可以看出, 从POM、浮游植物、浮游动物到杂食性鱼类的浮游牧食链在整个食物网中具有主导性, 而从水生植物、沉积物和底栖动物到杂食性鱼类的底栖食物链相对重要性较低(图 6B), 表明基于食性数据的理论食物网与BIMM模型预测的食物网总体吻合度很高(图 3和图 6B)。

图2 食物网食性矩阵(A)与依赖性矩阵(B)Fig. 2 Food web diet matrix (A) and dependency matrix (B)
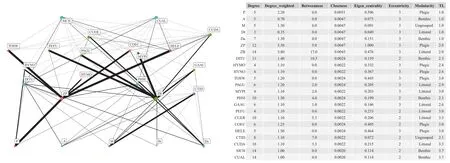
图3 基于理论食性矩阵的食物网Fig. 3 Food web based on theoretical diet matrix

图4 贝叶斯同位素混合模型碳、氮同位素预测值与测量值分布Fig. 4 δ13C and δ15N predicted value of Bayesian Isotope Mixing Model and measured value distribution
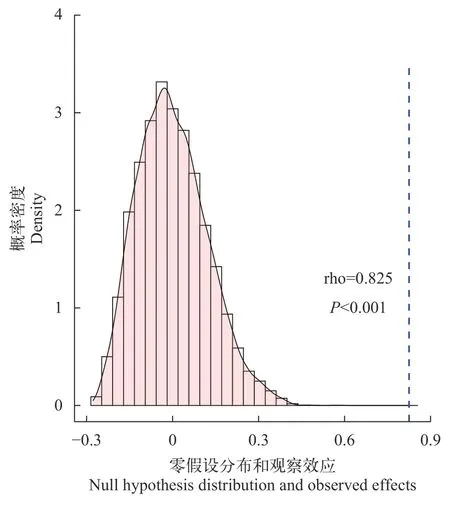
图5 理论食性数据矩阵与BIMM模型预测食性矩阵相关性分析Fig. 5 Correlation analysis between traditional diet matrix and BIMM model predictive diet matrix
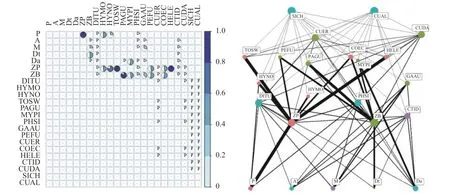
图6 基于贝叶斯同位素混合模型预测的食性矩阵(A)和食物网(B)Fig. 6 Diet matrix (A) and food web (B) based on Bayesian Isotope Mixing Model
3 讨论
3.1 不同类型食物网对比分析
目前, 国内利用碳、氮稳定同位素技术研究水域食物网大多通过δ13C和δ15N值的特征、空间分布特征、营养级及传统食源贡献率来分析, 而利用贝叶斯同位素混合模型(BIMM)结合δ13C和δ15N值进行食物网中各类消费者的食性预测, 并构建食物网的研究还很少。通过将BIMM模型预测的食物网与基于理论食性数据构建的食物网进行对比分析后发现, 两者具有很强的相关性, 且食性矩阵与食物网结构特征契合度很高。这表明基于BIMM模型的食物网中各类消费者的食性预测方法合理可靠, 在应用同位素技术研究水域食物网的过程中,具有一定的借鉴作用。
3.2 保安湖食物网结构分析
两个食性矩阵结果表明, POM、浮游植物、浮游动物与底栖动物是保安湖水域食物网中鱼类的主要食物来源, 其次是沉积物中的碎屑和水生植物等, 陆源有机物对整个食物网的贡献不大。两个食物网的结构特征则表明, 从POM、浮游植物、浮游动物到杂食性鱼类的浮游牧食链在整个湖泊食物网中具有占据主导地位, 而由水生植物、沉积物和底栖动物到杂食性鱼类的底栖食物链相对于浮游牧食链较弱。综合食性矩阵与食物网结构特征结果, 表明POM与浮游植物已成为保安湖食物网中最重要的基础食物源。这可能与湖泊的营养程度有关, 保安湖历史上与长江相通, 20世纪90年代初总氮和总磷可达地表水Ⅱ类—Ⅲ类水标准(GB3838-2002)[33], 属于中营养型湖泊, 也是良好渔业水体[34]。但近几十年来由于湖岸周边人口数量增加与渔业利用强度的加大, 加上周边农业活动的影响, 保安湖的营养盐浓度不断升高, 已发展成为中富营养型湖泊。Carpenter等[35]研究发现当湖中营养盐浓度升高时, 湖内部初级生产力对于消费者的贡献增加,而外源输入对消费者的贡献降低。因此, 这可能是由于营养盐浓度的提升促进了浮游植物的生长, 从而使得浮游植物的贡献升高。
3.3 保安湖食物网调控建议
保安湖中鱼类消费者的主要营养级范围为2.1—3.3。在调查到的16种鱼类中, 顶级肉食性鱼类种类很少, 仅有翘嘴鲌和鳜两种, 而杂食性鱼类的种类最多。保安湖大多数杂食性鱼类属于次级消费者, 以浮游动植物和底栖动物为主要食源(图 3—6)。此外, 在浅水富营养湖泊, 藻类是食物网中消费者的主要碳源, 而藻类是浮游动物的必需食物,也可以为杂食性鱼类提供稳定食物来源[36]。因此,水体营养水平升高导致浮游藻类成为最主要基础食物源可能是保安湖杂食性鱼类种类较多的原因之一。虽然杂食性鱼类的多样化会降低外界干扰对食物网结构与功能的影响[37], 但以杂食性为主的小型鱼类成为优势种后会导致浮游动物被过度摄食, 不能有效抑制藻类的过度生长。此外, 在保安湖生态系统中, 高营养级的凶猛鱼类种类偏少, 且数量偏低, 无法有效缓解牧食浮游动物的压力。因此, 对于保安湖的降低水华风险的食物网调控建议为: (1)增加滤食性鱼类数量, 利用其主动滤食方式直接降低浮游植物数量; (2)增加凶猛性鱼数量, 降低水体中浮游动物食性鱼类, 提升浮游动物数量,从而加大浮游动物对浮游植物的摄食强度, 降低浮游植物的数量; (3)增加大型滤食性底栖动物数量,利用其被动滤食方式, 滤取并降低浮游植物数量。
