广东南雄和新会农田鼠情监测结果分析
2022-06-07林思亮黄立胜蓝子胜赖荣丰姚丹丹姜洪雪冯志勇
林思亮,黄立胜,蓝子胜,赖荣丰,姚丹丹,姜洪雪,冯志勇
(1.广东省农业科学院植物保护研究所/广东省植物保护新技术重点实验室,广东 广州 510640;2.广东省农业有害生物预警防控中心,广东 广州 510500;3.江门市农作物病虫测报中心站,广东 江门 529000;4.南雄市植保植检管理站,广东 南雄 512400)
【研究意义】鼠类是地球上种类最多、分布最广、数量最大的哺乳动物,其中一些种类对农业生产、人类健康及公共安全可造成巨大危害[1-3]。广东省是我国农业鼠害的高发地区之一,由于气候及农业产业结构等原因,农业害鼠的繁殖快且强度大,害鼠长年处于中等偏重、局部重发生的趋势,对各种农作物的生产均有严重威胁[4]。开展农田鼠情监测工作,及时了解鼠种种类、鼠种组成及种群数量变化规律,是农田鼠害预测预报及防治工作的重要基础,可为各级主管部门开展鼠害防控工作决策提供科学依据。【前人研究进展】近年来,农村城镇化进程加快,农业产业结构持续变化,鼠类栖息环境发生较大变化,鼠类群落结构亦随之产生变化[5]。国内外对鼠类群落结构变化的研究已有许多报道[6-10]。在广东省内,冯志勇等[11]研究了珠江三角洲稻区鼠类的群落结构及演替规律,发现鼠类群落持续变动,农田作物布局改变是主要驱动因子。高志祥等[4]探讨种植区环境对鼠类群落结构的影响,发现广东省内5 个地市种植区的作物结构变化、农村城镇化引起了鼠类群落结构的变化。姚丹丹等[12]发现农村城镇化导致农田鼠类群落发生一定程度的改变,家栖鼠类数量大幅增加,群落生物量加大,野栖鼠类优势度明显下降。综上,作物结构、农村城镇化是农田鼠类群落结构变化的重要驱动因子,是鼠害灾变过程中需要关注的问题之一。【本研究切入点】珠三角地区与粤北山区的城镇化率、作物结构及种植技术水平等发展甚为不均,鼠类栖息环境具有较大差异。但针对粤北山区与珠三角的农田鼠类群落结构差异分析却鲜有报道。因此,本研究对粤北山区的韶关南雄市、珠江三角洲的江门市新会区2 个监测点2018—2020 年连续3 年的鼠情监测数据进行分析,旨在掌握监测区农田鼠种组成情况和种群数量的变化趋势,并探讨不同栖息环境下鼠类群落结构特征的差异,以此为广东省农田鼠害预测预报及防治工作提供科学依据。【拟解决的关键问题】鼠类种群动态监测作为农田鼠害防控的重要基础工作之一,城镇化率、作物结构各异的农田鼠类种群结构特征差异如何,科学客观地掌握鼠类种群的动态变化趋势,对于因地因时调整鼠害防控策略及方法,保障广东省农业生产安全、人民生命健康具有重要意义。
1 材料与方法
1.1 监测地点及时间
监测地点为广东省粤北山区的韶关南雄市、珠江三角洲的江门市新会区2 地的典型农田区域(包括稻田和旱地耕作区)。
监测时间为2018—2020 年,连续监测3 年,每个监测点分别监测36 月次。
1.2 调查方法
每个监测点每年调查12 个月,调查方法为夹夜法,调查工具采用大号铁板夹,主要在监测点的农田(包括稻田和旱地耕作区)开展调查。采用直线或曲线排列,以5 m 的间隔进行鼠夹布防,花生仁作诱饵,晚放晨收。对捕获的鼠类标本进行编号、鉴定鼠种及性别、外部形态指标测量和解剖观察其繁殖状况。
1.3 统计方法
根据调查结果分别统计捕获率和鼠种组成,以捕获率(鼠密度)表示其种群数量。并根据有效夹数所捕获的鼠类体重之和表示生物量。

2 结果与分析
2.1 种类及组成
2018—2020 年共置夹13 131 个,捕获各类鼠类样本836 只,总捕获率为6.37%。经鉴定,捕获鼠类隶属于3 属5 种,分别为鼠属的黄毛鼠(Rattus losea)、褐家鼠(Rattus norvegicus)和黄胸鼠(Rattus tanezumi),小家鼠属的卡氏小鼠(Mus caroli)和板齿鼠属的板齿鼠(Bandicota indica)。在捕获样本中,黄毛鼠占捕获总数的70.45%,捕获率4.49%,为优势密度鼠种;板齿鼠占捕获总数的11.84%,捕获率0.75%,为次优势密度鼠种;卡氏小鼠占捕获总数的11.24%,捕获率0.72%,也为次优势密度鼠种;虽然褐家鼠和黄胸鼠占捕获总数的比例不高,但也有一定的捕获数量(表1)。综上,监测区农田鼠类的基本群落结构可以概括为“黄毛鼠+板齿鼠+卡氏小鼠+褐家鼠+黄胸鼠”,其中黄毛鼠为农田的优势密度鼠种。

表1 2018—2020 年监测点农田鼠种Table 1 Rodent species in farmlands of monitoring sites from 2018 to 2020
2.2 种群数量
2.2.1 年度变化趋势 新会监测点在2018—2019年的捕获率维持在5%以上,到2020 年下降至2.31%;南雄监测点3 年的捕获率维持在8%上下波动,其中2020 年捕获率最高,达到9.45%,是新会区监测点当年的4.1 倍(图1)。

图1 2018—2020 年监测点鼠类种群数量年度变化Fig.1 Annual change of rodent population at monitoring sites from 2018 to 2020
2.2.2 月度变化趋势 监测区不同月份农田鼠类种群数量变化呈双峰型。1 月为第1 个高峰,平均捕获率为7.92%;之后逐渐下降,至4 月达到全年最低值,平均捕获率为4.03%;之后略有回升,至6 月达到较高水平,平均捕获率为7.49%;随后下降到7 月的低谷期,平均捕获率为5.01%;随后逐月上升,到11 月出现第2 个高峰,为全年最高值,平均捕获率为8.57%(图2)。

图2 2018—2020 年监测点鼠类种群数量月度变化Fig.2 Monthly change of rodent population at monitoring sites from 2018 to 2020
2.2.3 季度变化趋势 监测区冬季的农田鼠类种群数量最高,平均捕获率为7.17(±2.49)%;春季最低,平均捕获率为4.86(±2.21)%,最高季节平均捕获率为最低季节的1.48 倍。平均捕获率冬季>秋季>夏季>春季(图3)。

图3 2018—2020 年监测点鼠类种群数量季度变化Fig.3 Quarterly change of rodent population at monitoring sites from 2018 to 2020
2.3 生物量结构
监测区内害鼠种类多为植食性物种,害鼠生物量可作为其对农作物危害程度最直观的指示指标[4,11]。监测期间农田鼠类群落中,黄毛鼠平均体重为71.87 g,其生物量最大,总百夹夜生物量为322.34 g,占捕获总生物量的54.75%。其次是板齿鼠,总百夹夜生物量为234.22 g,占捕获总生物量的29.99%,但捕获板齿鼠平均体重仅为234.22 g,说明捕获的个体多为幼体和亚成体。之后是褐家鼠,总百夹夜生物量为71.13 g,占总捕获生物量的12.08%(表2)。由此可认定,黄毛鼠和板齿鼠是对监测区农田为害最大的害鼠。
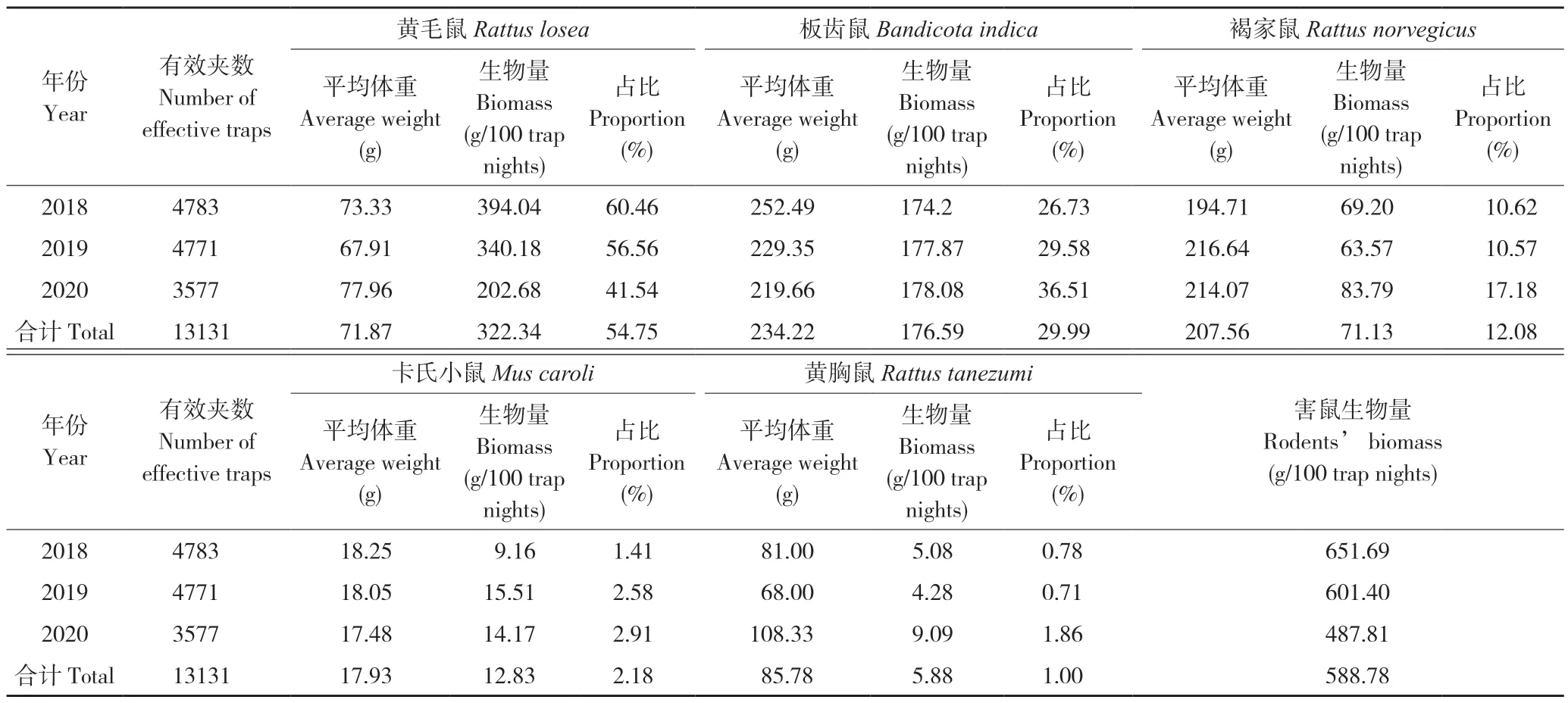
表2 2018—2020 年监测点害鼠群落生物量结构Table 2 Biomass structure of rodent community at monitoring sites from 2018 to 2020
2.4 监测点间差异
2.4.1 鼠种组成差异 2 个监测点皆捕获5 个鼠种,而经χ2检验在鼠种组成上存在极显著差异(χ2=62.589,P<0.001***)。黄毛鼠在2 个监测点均为优势鼠种,占比均在55%以上(表3)。
2.4.2 种群数量差异 南雄监测点的平均捕获率为8.38(±5.05)%,显著高于新会监测点的4.88(±4.28)%(P=0.003**),新会监测点捕获率变异系数达到87.7%,高于南雄监测点的60.3%,说明新会监测点鼠类种群数量的月间变化程度较高(表3)。

表3 不同监测点农田鼠密度及各种害鼠数量构成比率Table 3 Rodent density and species composition ratio of various monitoring sites
2.4.3 生物量结构差异 害鼠群落生物量构成在2 个监测点存在极显著差异(χ2=119.18,P<0.001***),南雄监测点害鼠总百夹夜生物量(727.92 g)显著高于新会监测点(472.52 g)。新会监测点农田鼠类群落中,板齿鼠生物量最大,总百夹夜生物量为213.10 g,占捕获总生物量的45.10%;其次为黄毛鼠,总百夹夜生物量为197.78 g,占捕获总生物量的41.86%。南雄监测点农田鼠类群落中,黄毛鼠生物量最大,总百夹夜生物量为471.44 g,占捕获总生物量的64.76%;其次为板齿鼠,总百夹夜生物量为132.89 g,占捕获总生物量的18.26%(表4)。

表4 不同监测点害鼠群落生物量结构Table 4 Biomass structure of rodent community at various monitoring sites
3 讨论
从组成来看,黄毛鼠、板齿鼠的捕获数占比分别为70.45%、11.84%;从生物量来看,黄毛鼠、板齿鼠的生物量占比分别为54.75%、29.99%,这2 种鼠类是监测区农田的优势种和为害最大的害鼠;同时由于城镇化,家栖鼠类(褐家鼠、黄胸鼠)逐渐扩散至农田,占据一定的比例。该结果与东莞、中山、湛江、梅州和广州[4,11-13]等地的害鼠群落结构一致。此外,板齿鼠捕获数占比为11.84%,低于同期黄立胜等[14]使用物联网智能监测系统在南雄的监测结果,我们捕获板齿鼠个体多为幼体和亚成体,说明夹夜法未能准确反映板齿鼠的种群密度,板齿鼠应具有更高的密度。综上,黄毛鼠和板齿鼠应为广东省农田鼠害防治主要的监测和防治对象,同时应密切关注家栖鼠类迁入农田的趋势。
鼠类种群数量动态呈双峰型,每年有2 个高峰期(1 月和11 月,捕获率分别达到7.92%和8.57%)和2 个低谷期(4 月和7 月,捕获率分别为4.03%和5.01%),该结果与中山农田害鼠种群数量变化规律一致,皆呈现双峰型的规律[15]。环境因子、土地利用方式及人为活动直接或间接地影响鼠类种群数量消长[16]。农田鼠类种群数量的季节动态应与灭鼠次数有关,监测区的农区在每年会全面灭鼠2 次:1 次在春播春种前,1 次在夏种结束后,这应是鼠类种群数量动态呈现双峰型的主要原因之一[15]。
2 个监测点在鼠种组成、种群数量及群落生物量结构上皆存在显著差异。究其原因,应是2个监测点的生境差异所造成的,由于新会监测点种植作物结构调整,许多稻田转变为柑橘种植园,减少鼠类食物来源和适宜栖息地,限制种群增长[16-20];而在南雄监测点,仍存在较大面积的稻田为鼠类提供食物来源,并有部分丢荒杂草丛生的土地,形成鼠类适宜的栖息地,利于种群增长[16-20]。由于本研究仅对鼠类种群及生物量开展了调查监测,而关于鼠类种群动态与农田的农作物种植结构动态的关系还有待继续深入研究。
鼠害治理工作是一项复杂的社会工程,科学地开展鼠害持续控制,对于保障广东省农业生产安全、人民生命健康具有重要意义[21]。本研究结果表明,农田鼠种的种群数量在不同年度、不同月份、不同季节存在明显的差异,且在不同地区,鼠种组成、种群数量及群落生物量结构亦存在显著的差异。因此,一方面需要提高鼠类监测数据质量,有效科学地监测农田鼠类种群动态及规律的同时,还应对农作物种植结构动态进行监测,以预测鼠类种群未来的潜在动态,进一步提高农田害鼠早期预警的准确度[15,22-25]。另一方面,既要做好全省鼠害的统防统治,亦要因地制宜开展精准防控,根据监测获得的鼠情发生规律,特别要注意因种植结构调整、栖息地状况发生重大改变的区域,需根据监测结果及时调整防控策略和方法[23-26]。
4 结论
本研究结果表明,黄毛鼠和板齿鼠是广东农田鼠害防治主要的监测和防治对象。监测点农田鼠类种群数量在不同月份、不同季节存在明显差异,1 月和11 月是鼠类种群数量的高峰,4 月和7 月是种群数量的低谷,种群数量呈双峰型的变化趋势。此外,冬季是鼠类种群的高峰季节,捕获率可达到7.17%,是最低季节的1.48 倍。2 个监测点由于生境差异造成其在鼠种组成、种群数量和群落生物量结构上皆存在显著差异。提高鼠类监测质量,根据监测结果动态调整防控策略和方法,是实现新形势下鼠害可持续治理目标的必由之路。
