不同环境因素对苘麻种子萌发及出苗的影响
2022-06-07耿志同韩玉军张永倩陶波马红
耿志同 韩玉军 张永倩 陶波 马红


摘要 苘麻Abutilon theophrasti Medikus具有生长旺盛、结籽量大和适应性强等特点,已成为多种农田中的恶性杂草,研究环境条件对苘麻萌发的影响对进一步探究其生物学特性与综合防治技术具有重要意义。通过室内试验研究温度、光照、酸碱度、水分胁迫、盐胁迫与苘麻种子萌发及埋土深度与其出苗的关系。结果表明,在10~35℃下苘麻种子均可萌发,20~30℃为其最适萌发温度范围;种子萌发对光照不敏感;适应pH范围广,在pH 4~9条件下种子发芽率均在70%以上;水势在-0.2~0 MPa范围内种子发芽率最高,当水势小于-0.7 MPa时,种子萌发受到完全抑制;有较强的耐盐性,NaCl浓度为160 mmol/L时发芽率超过40%;种子对播种深度适应性较强,覆盖不超过8 cm的土层均可出苗。
关键词 苘麻;萌发;出苗;环境条件
中图分类号: S451
文献标识码: A
DOI: 10.16688/j.zwbh.2021208
Abstract Abutilon theophrasti Medikus has become a malignant weed in many fields because of its strong growth, large seed bearing capacity and strong adaptability. It is of great significance to study the effects of environmental factors on the germination of A.theophrasti for further research on its biological characteristics and comprehensive control technology. The relationship between temperature, light, pH value, water, salt stress and seed germination, burial depth and seedling emergence of A.theophrasti was studied by laboratory experiments. The results showed that the seeds of A.theophrasti could germinate at the temperature of 10-35℃, and the optimum temperature range was 20-30℃. Seed germination was not sensitive to light. It was suitable for a wide range of pH, and the germination rate of seeds was above 70% at pH4-9. The germination rate of seeds was the highest when the water potential was in range of -0.2 to 0 MPa, and the germination was completely inhibited when the water potential was less than -0.7 MPa. It had a strong salt tolerance, and the germination rate was over 40% when the NaCl concentration was 160 mmol/L. The seeds had strong adaptability to burial depth, and seedlings could emerge from the soil layer of a depth of no more than 8 cm.
Key words Abutilon theophrasti;germination;emergence;environmental factors
苘麻Abutilon theophrasti Medikus又名青麻、麻果,隸属于锦葵科苘麻属的一年生草本植物,是我国多种农作物田间的主要恶性杂草之一[1]。苘麻生长旺盛,枝叶繁茂,在与作物共生时会截获大部分光照,从而抑制作物对水分和养料的转化,造成显著的产量损失[2]。有研究报道,苘麻在与大豆和棉花的竞争中占有显著的优势,密度超过2株/m2(或2株/m 行长)的苘麻会严重影响大豆和棉花的产量及品质[3-4],高密度的苘麻(12株/m2以上)会使玉米严重减产甚至接近绝产[5]。除此之外,苘麻具有结实量大,种子活力高的特点,这使其易形成难以彻底清除的土壤种子库从而产生持续性的危害[6]。
杂草种子是杂草防除的重要靶标,系统地了解杂草种子的萌发特性可对杂草进一步发生和传播的方向进行科学合理的预测,为杂草的综合防除提供理论指导[7]。例如:过低的水势会严重抑制菵草Beckmannia syzigachne[8]、Malva parviflora[9]的萌发,因此这类喜湿性杂草多在水田或湿润的稻茬麦田发生,在旱田几乎不能出苗;碎米知风草Eragrostis japonica为光敏感型种子,无光条件下不能萌发[10];超过2 cm的土层可以完全遏制马齿苋Portulaca oleracea种子出苗[11],因此可以通过耕翻等农业措施对其进行防除。
目前国内外学者对苘麻种子的休眠及萌发进行了一些研究。低温处理及60℃温水浸种与赤霉素浸种结合处理均可以有效地打破苘麻种子的休眠[12-13]。王金淑的研究发现,苘麻种子为需光型种子,适宜其萌发的温度为15~30℃,pH 4~8[14]。邢维芹等的研究结果显示,光照对苘麻种子的发芽率并无显著影响[15]。张秀玲的研究发现,苘麻种子的发芽率随NaCl或Na2CO3溶液浓度的增加而下降[16]。Sadeghloo等[17]的研究结果显示,伊朗地区的苘麻在5℃下即可萌发,最适萌发温度为35℃,其萌发较耐酸碱和盐分而不耐干旱。然而国内有关影响苘麻萌发出苗因素的详细报道较少。因此,本研究选择温度、光照、pH、盐浓度、水势及埋土深度等指标,明确其对苘麻种子萌发与出苗的影响,以期为苘麻发生区域预测及综合防控提供理论依据和数据支撑。BF751EA3-644D-4795-BEC9-C8EEE8102281
1 材料与方法
1.1 试验材料
供试种子:苘麻种子于2020年采自黑龙江省哈尔滨市东北农业大学大豆试验田,将种子自然风干后选取其中成熟饱满的种子用60℃温水浸泡15 min解除休眠(经测定浸种结束后种子未吸胀),发芽率达90%以上。
试剂及仪器:所用试剂均为国产分析纯。BCD-641WASN 冰箱,青岛海尔股份有限公司;LRH-250光照培养箱,上海一恒科学仪器有限公司;FE-28 pH计,梅特勒-托利多仪器(上海)有限公司。
1.2 试验方法
1.2.1 苘麻种子萌发基础条件
选取大小一致的苘麻种子,用蒸馏水反复冲洗3次,均匀置入铺有双层滤纸的直径9 cm玻璃培养皿中,每皿30粒,加入15 mL蒸馏水或相应溶液,用封口膜封口后置入光∥暗温度为25℃∥20℃的光照培养箱中,L∥D=12 h∥12 h,光照强度为8 000 lx。以胚根露出种皮为发芽标准,每隔24 h观察记录种子萌发数并移除已发芽的种子,调查持续至连续3 d无种子发芽为止。每试验重复3次。
1.2.2 温度对苘麻种子萌发的影响
设置恒温和变温两种条件。恒温条件下分别设置5、10、15、20、25、30℃和35℃,变温条件的光∥暗温度分别设置为15℃∥5℃、20℃∥10℃、25℃∥15℃、30℃∥20℃、35℃∥25℃,其余培养条件同1.2.1。
1.2.3 光周期对苘麻种子萌发的影响
将培养箱光∥暗时间分别设置为0 h∥24 h、6 h∥18 h、12 h∥12 h、18 h∥6 h、24 h∥0 h, 其余培养条件同1.2.1。
1.2.4 pH对苘麻种子萌发的影响
设定缓冲溶液pH为4、5、6、7、8和9, 其中pH为4的缓沖溶液用2 mmol/L的邻苯二甲酸氢钾与1 mol/L的HCl溶液配制;pH为5和6的缓冲溶液用2-吗啉乙磺酸与1 mol/L的NaOH溶液配制;pH为7和8的缓冲溶液用2 mmol/L的4-羟乙基哌嗪乙磺酸与1 mol/L的NaOH溶液配制;pH为9的缓冲溶液用N-三(羟甲基)甲基甘氨酸与1 mol/L的NaOH溶液配制。使用pH计校对溶液pH,以蒸馏水为对照,其余培养条件同1.2.1。
1.2.5 盐胁迫对苘麻种子萌发的影响
分别配制浓度为0、10、20、40、80、120、160、240 mmol/L 和320 mmol/L的NaCl溶液进行盐胁迫试验, 其余培养条件同1.2.1。
1.2.6 水分胁迫对苘麻种子萌发的影响
配制不同浓度的PEG 6000溶液以模拟水势胁迫,分别溶解0、75.4、105.3、133.6、160.3、185.5、209.0、230.9、251.3、270.0 g和287.2 g PEG 6000于1 L蒸馏水中,使各处理溶液水势分别为-0.1、-0.2,-0.3、-0.4、-0.5、-0.6、-0.7、-0.8、-0.9 MPa 和-1.0 MPa[18],其余培养条件同1.2.1。
1.2.7 埋土深度对苘麻种子出苗的影响
埋土深度分别设置为1、2、3、4、5、6、7、8、9、10、11、12、13、14 cm和15 cm,每处理4次重复,在高约20 cm、直径约25 cm的底部打孔塑料盆中装入厚度约5 cm的试验用土,每盆内均匀播撒20粒苘麻种子,将盆钵置于托盘中,试验期间向托盘中加水使盆钵从底部吸水,培养箱设置同1.2.1。播种后15 d记录出苗数。
1.3 数据统计与分析
试验所得数据采用SPSS 14.0软件采用LSD法进行显著性检验,利用SigmaPlot 14.0软件对不同NaCl浓度和不同水势下苘麻种子的发芽率及不同埋土深度下苘麻种子的出苗率进行曲线拟合,回归模型如下:
G=Gmax/[1+(x/x50)Srate],G为发芽率或出苗率,Gmax为最大发芽率或最大出苗率,x为溶液浓度、水势或埋土深度,x50为G=Gmax/2时对应的x值,Srate为曲线斜率。
2 结果与分析
2.1 温度对苘麻种子萌发的影响
温度对苘麻种子的萌发影响显著,苘麻种子的发芽率随温度上升呈先上升后下降的趋势(图1)。恒温条件下,温度为5℃时苘麻种子不能萌发,在10~35℃的条件下均能萌发(图1a)。其中20~30℃下发芽率最高,均在90%以上;低于20℃或高于30℃,发芽率显著下降,10℃、15℃和35℃的发芽率分别为26.67%、82.22%和74.44%。变温条件下,光∥暗温度为15℃/5℃时苘麻种子不能萌发,20℃∥10℃与35℃∥25℃下苘麻种子的发芽率均显著低于25℃∥15℃与30℃∥20℃(图1b)。说明适宜苘麻种子萌发的温度范围较广,但温度过低或过高均会抑制苘麻种子的萌发。
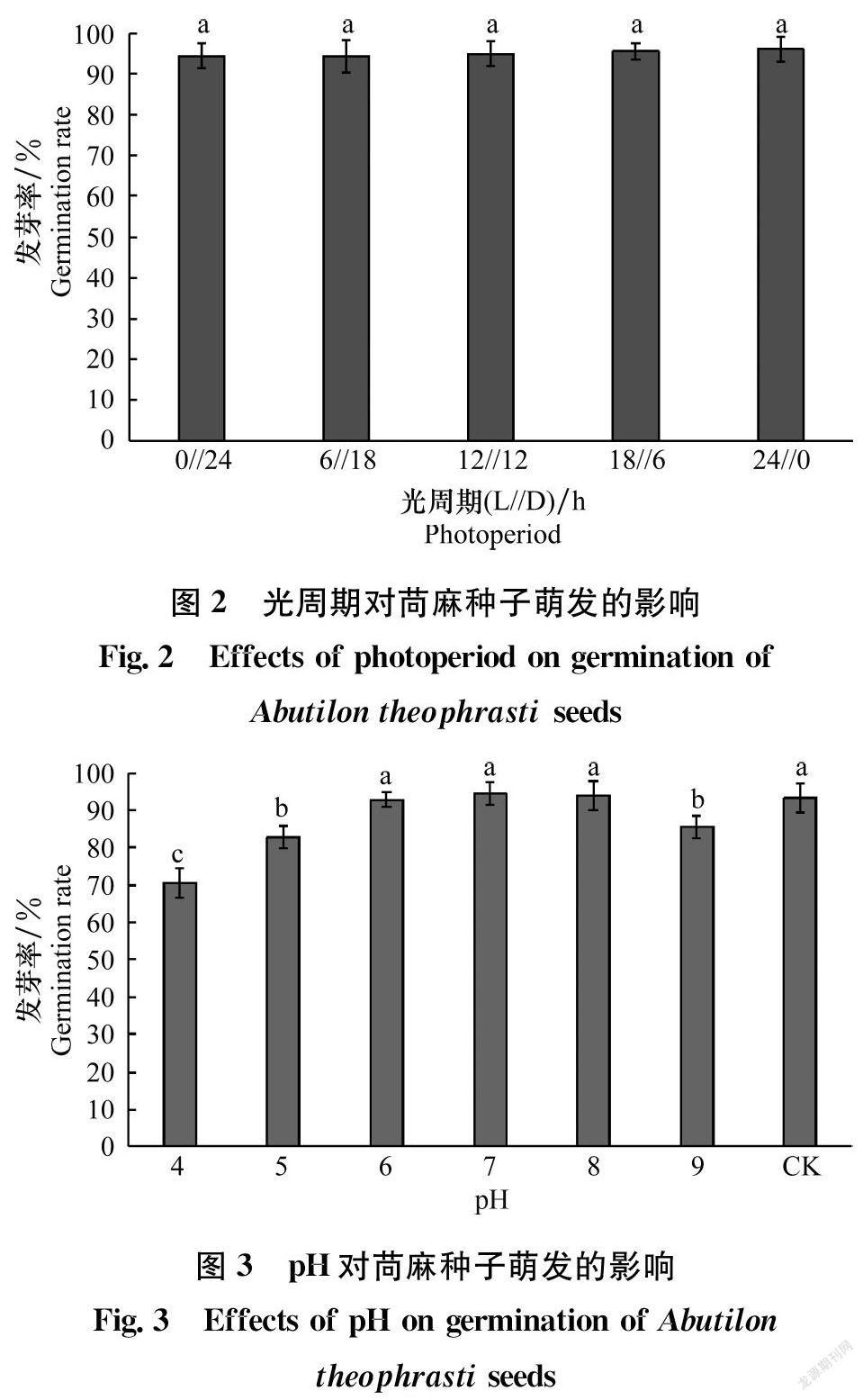
2.2 光周期对苘麻种子萌发的影响
在不同光周期处理下,苘麻种子发芽率均在90%以上,且处理间差异不显著(图2)。说明苘麻种子的萌发对光周期不敏感。
2.3 pH对苘麻种子萌发的影响
苘麻种子在pH 4~9的范围内均可萌发,且发芽率均在70%以上(图3)。pH为6、7、8处理下的发芽率与对照(蒸馏水)相比差异不显著,且均在90%以上。pH为5和9处理下苘麻种子的发芽率显著低于对照,分别为82.78%和85.56%。pH为4的处理苘麻种子发芽率显著低于其他处理,但仍高达70.56%,说明苘麻种子对pH有广泛的适应性,土壤酸碱度并不是限制苘麻种子萌发的主要因素。
2.4 盐胁迫对苘麻种子萌发的影响
苘麻种子的发芽率与NaCl溶液浓度的关系见图4。在NaCl浓度不超过40 mmol/L范围内,苘麻种子的发芽率均高于90%,且各处理间差异不显著;随着NaCl浓度由40 mmol/L上升至240 mmol/L,苘麻种子发芽率随着NaCl浓度的上升显著下降;当NaCl浓度上升至320 mmol/L时,仅有5.09%的苘麻种子萌发。对不同浓度NaCl溶液下苘麻种子的发芽率进行曲线拟合,得到方程G=91.79/[1+(x/146.85)3.64](R2=0.99),模拟得出抑制其发芽率为最高发芽率50%时的NaCl浓度为146.85 mmol/L。BF751EA3-644D-4795-BEC9-C8EEE8102281
2.5 水势胁迫对苘麻种子萌发的影响
水势变化对苘麻种子萌发有显著影响(图5)。在-0.2~0 MPa的水势范围内,苘麻种子的发芽率均在90%以上,且处理间无显著差异,当水势低于-0.2 MPa时,苘麻种子的发芽率随水势降低呈显著下降趋势,当水势低至-0.7 MPa时,苘麻种子的萌发受到完全抑制。对不同水势下苘麻种子的发芽率进行曲线拟合得到方程G=92.12/{1+[x/(-0.39)]5.99}(R2=0.99),模拟得到抑制苘麻种子发芽率为最高发芽率50%时的水势为-0.39 MPa。
2.6 埋土深度对苘麻种子出苗的影响
在埋土深度1~8 cm的范围内,苘麻种子均能出苗(图6)。在埋土深度1~3 cm内,苘麻种子的出苗率均能达到80%以上,且处理间差异不显著。埋土深度超过3 cm,苘麻种子的出苗率随着埋土深度的增加而下降,在埋土深度达到8 cm时,仍有少部分苘麻种子可以出苗。对不同埋土深度下苘麻种子的出苗率进行曲线拟合得到方程G=88.10/[1+(x/5.47)6.45](R2=0.99),模拟得到当苘麻种子出苗率达最高出苗率50%时的埋土深度为5.47 cm。
3 结论与讨论
种子的萌发与外界环境条件紧密相关,如温度、光照、土壤的水分、酸碱度和盐浓度以及种子所处深度等[19-20]。杂草种群的建立和传播与其对外界环境条件的适应能力密不可分。本研究结果显示,苘麻种子适宜萌发的温度范围广但不耐低温,这一结果与Sadeghloo等[17]的研究结果存在差异,这种差异可能由于不同地区的苘麻对温度的适应性有所不同;苘麻的萌发需要较高的水势支持,虽然苘麻属于典型的旱生杂草,但也需要在较为湿润的环境下才能萌发,这可能与其种皮透水性差有关,这种特性也与同属旱生杂草的猪殃殃Galium aparine var. tenerum[21]较为一致。我国大部分地区处于温带、亚热带和热带,在晚春和夏季降水充足,温度适宜,因此可以预测此时期为苘麻的出苗高峰期,在苘麻与作物形成竞争之前应对其予以集中防除。光照对其萌发的影响较小,这一结果与王金淑的研究略有差异[14],与邢维芹的研究结果较为一致[15]。这种差异可能来源于供试种子的产地或储存方式、时间的不同。与其他杂草相比[10-11, 21-23],苘麻对酸碱胁迫与盐胁迫具有一定的耐受性,因此我国大部分地区的盐碱土壤不会限制苘麻的发生与危害。除此之外,苘麻种子的拱土能力较强,生产中可以通过适度的深耕等农业措施在播种前将土壤表层的苘麻种子深翻至地下以减小其发生密度。
目前对苘麻的防除措施主要为化学防除,但长期重复使用除草剂不仅会使苘麻产生抗药性,还会造成环境污染等危害。通过对以上特性的明确可以对苘麻的适生区域及发生时期进行科学的预测,减少对化学除草剂的依赖,更好地为苘麻的综合防除提供理论指导,也能为室内试验种植苘麻的方法提供参考。
参考文献
[1] 李扬汉.中国杂草志[M].北京: 中国农业出版社, 1988: 694-695.
[2] ZISKA L. Observed changes in soyabean growth and seed yield from Abutilon theophrasti competition as a function of carbon dioxide concentration [J]. Weed Research, 2013, 53(2): 140-145.
[3] 王义明. 龙葵和苘麻发生数量与大豆产量的关系[J].杂草科学, 1996(3): 10-11.
[4] CORTS J A, MENDIOLA M A, CASTEJN M. Competition of velvetleaf (Abutilon theophrasti M.) weed with cotton (Gossypium hirsutum L.). Economic damage threshold [J]. Spanish Journal of Agricultural Research, 2010, 8(2): 391-399.
[5] MCDONALD A J, RIHA S J, MOHLER C L. Mining the record: historical evidence for climatic influences on maize-Abutilon theophrasti competition [J]. Weed Research, 2004, 44(6): 439-445.
[6] LUESCHEN W E, ANDERSEN R N. Longevity of velvetleaf (Abutilon theophrasti) seeds in soil under agricultural practices [J]. Weed Science, 1980, 28(3): 341-346.
[7] 田志慧, 沈國辉. 杂草种子休眠与萌发调控的研究进展[J]. 上海农业学报, 2015, 31(2): 137-141.
[8] RAO Na, DONG Liyao, LI Jun, et al. Influence of enviromental factors on seed germination and seedling emergence of American sloughgrass (Beckmannia syzigachne) [J]. Weed Science, 2008, 56(4): 529-533.
[9] CHAUHAN B S, GILL G, PRESTON C. Factors affecting seed germination of little mallow (Malva parviflora) in southern Australia [J]. Weed Science, 2006, 54(6): 1045-1050.BF751EA3-644D-4795-BEC9-C8EEE8102281
[10]张培培. 碎米知风草(Eragrostis japonica)生物学生态学特性及化学防除技术研究[D].南京: 南京农业大学, 2014.
[11]杨彩宏, 冯莉, 岳茂峰. 恶性杂草马齿苋(Portulaca oleracea)种子萌发特性的研究[J]. 植物保护, 2009, 35(1): 62-65.
[12]邓天福, 王霞. 不同植物生长调节剂和温度处理对苘麻种子萌发的影响[J]. 河南科技学院学报(自然科学版), 2015, 43(1): 20-24.
[13]常青山, 张利霞, 刘晶, 等. 苘麻种子休眠机理及破眠方法[J]. 植物生理学报, 2016, 52(6): 967-974.
[14]王金淑. 光照和温度等因素对苘麻种子萌发特性的影响[J]. 北方园艺, 2012(1): 50-51.
[15]邢维芹, 刘辉, 曾冰, 等. 光照和浸种对5种具有修复重金属污染土壤潜力的植物种子萌发的影响[J]. 种子, 2017, 36(9): 72-75.
[16]张秀玲. 不同盐胁迫对苘麻种子萌发的影响[J]. 江苏农业科学, 2008(1): 35-37.
[17]SADEGHLOO A, ASGHARI J, GHADERI-FAR F. Seed germination and seedling emergence of velvetleaf (Abutilon theophrasti) and barnyardgrass (Echinochloa crus-galli) [J]. Planta Daninha, 2013, 31(2): 259-266.
[18]MICHEL B E, KAUFMANN M R. The osmotic potential of polyethylene glycol 6000 [J]. Plant Physiology, 1973, 51(5): 914-916.
[19]CHACHALIS D, REDDY K N. Factors affecting Campsis radicans seed germination seed germination and seedling emergence [J]. Weed Science, 2000, 48(2): 212-216.
[20]ZHAO Ning, LI Qi, GUO Wenlei, et al. Effect of environmental factors on germination and emergence of shortawn foxtail (Alopecurus aequalis) [J]. Weed Science, 2018, 66(1): 47-56.
[21]張田田, 马冲, 吴翠霞, 等. 不同环境因素对猪殃殃种子萌发及出苗的影响[J]. 植物保护, 2017, 43(2): 102-106.
[22]李涛, 袁国徽, 钱振官, 等. 野燕麦种子萌发特性及化学防除药剂筛选[J]. 植物保护, 2018, 44(3): 111-116.
[23]王晓阳, 于海燕, 杨娟, 等. 不同环境因素对节节麦萌发的影响[J]. 植物保护, 2019, 45(3): 196-200.
(责任编辑:杨明丽)BF751EA3-644D-4795-BEC9-C8EEE8102281
