不同光周期下草地贪夜蛾两性生命表的比较
2022-06-07孟令贺江幸福李平夏吉星张同强程云霞张蕾
孟令贺 江幸福 李平 夏吉星 张同强 程云霞 张蕾
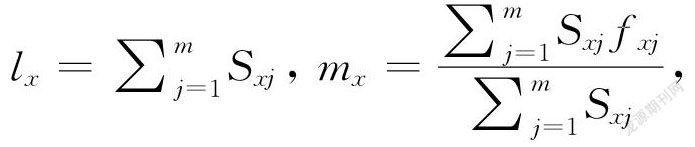




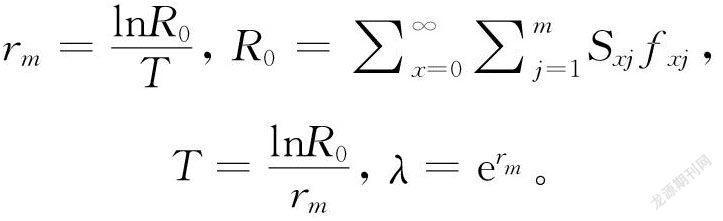



摘要 为了阐明光周期对草地贪夜蛾生长发育和生殖能力的影响,在25℃条件下,设置5个光周期(L∥D=16 h∥8 h、L∥D=14 h∥10 h、L∥D=12 h∥12 h、L∥D=10 h∥14 h和L∥D=8 h∥16 h)测定不同光周期条件下草地贪夜蛾的生长发育参数和生殖参数并建立两性生命表。结果表明,L∥D=12 h∥12 h时草地贪夜蛾的幼虫历期(17.53 d)和产卵前期(5.73 d)最长,L∥D=16 h∥8 h时幼虫历期(15.12 d)和产卵前期(4.18 d)最短。各发育阶段存活率在L∥D=16 h∥8 h 时最高,在L∥D=12 h∥12 h和L∥D=10 h∥14 h时最低。产卵整齐度(PFO)在L∥D=10 h∥14 h时最大(2.82 d),在L∥D=14 h∥10 h最小(1.42 d)。L∥D=14 h∥10 h时成虫的产卵量(1 950.64粒)、产卵历期(7.48 d)以及交配次数(3.55次)和交配率(95.25%)均高于其余光周期。L∥D=16 h∥8 h和L∥D=14 h∥10 h时净增殖率、内禀增长率和周限增长率均高于其余光周期。本文的结果表明,长光照条件下草地贪夜蛾的生殖能力和发育速度均高于短光照。
关键词 草地贪夜蛾;光周期;两性生命表
中图分类号: S435.132
文献标识码: A
DOI: 10.16688/j.zwbh.2021088
Abstract In order to elucidate the effect of photoperiod on the growth and reproductive capacity of the fall armyworm, under the conditions of 25℃, five photoperiods (L∥D=16 h∥8 h, L∥D=14 h∥10 h, L∥D=12 h∥12 h, L∥D=10 h∥14 h and L∥D=8 h∥16 h) were set to determine the growth parameters and reproductive parameters under different photoperiods, and the bisexual life table was established. The results showed that L∥D=12 h∥12 h had the longest larval duration (17.53 d) and the pre-oviposition duration (5.73 d), while L∥D=16 h∥8 h had the shortest larval duration (15.12 d) and the pre-oviposition duration (4.18 d). The survival rate was the highest at L∥D=16 h∥8 h, and the lowest at L∥D=12 h∥12 h and L∥D=10 h∥14 h. PFO (period of first oviposition) was the highest at L∥D=10 h∥14 h (2.82 d) and the lowest at L∥D=14 h∥10 h (1.42 d). The number of oviposition (1 950 eggs), duration of oviposition (7.48 d), mating frequency (3.55 times) and mating rate (95.25%) of the adults at L∥D=14 h∥10 h were higher than those at the other photoperiods. L∥D=16 h∥8 h and L∥D=14 h∥10 h resulted in higher increase rates of net, intrinsic and finite growth than other photoperiods. The results of this study suggested that the reproductive capacity and developmental rate of the fall armyworm under long photoperiods were higher than those under short photoperiods.
Key words Spodoptera frugiperda;photoperiod;bisexual life table
草地貪夜蛾 Spodoptera frugiperda (J. E. Smith),别名秋黏虫,英文名fall armyworm,属于鳞翅目Lepidoptera 夜蛾科Noctuidae,源于美洲的热带和亚热带地区,是一种多食性农业害虫[1-3]。草地贪夜蛾具有飞行能力强、繁殖数量大、暴食为害重、寄主范围广等特点。其幼虫能够为害玉米、水稻、高粱、棉花等主要农作物以及多种蔬菜、果树和观赏植物[4-7]。随着经济全球化以及国际贸易的进行,该虫于2016年首次入侵非洲[8-9],并迅速在非洲多数国家蔓延为害[10-11],对当地的粮食产量造成严重影响[12]。2018年草地贪夜蛾从印度半岛入侵亚洲其他地区[13],并于2019年1月从云南入侵我国,进而向其他地区蔓延为害[14-15]。
在自然条件下,昆虫受到多种环境因素,比如温度、湿度以及光周期等的影响。研究表明,光周期作为自然界中最稳定的信号,能够调节昆虫的行为和节律[16],对昆虫的羽化、产卵、交配和取食都有影响[17]。在光周期L∥D=12 h∥12 h时,草地螟Loxostege sticticalis Linne会出现明显的滞育现象[18]。对于无滞育特性的昆虫来说,光周期对其生长发育也有影响,比如
在相同温度不同光周期条件下,甜菜夜蛾Spodoptera exigua Hübner的幼虫历期以及蛹期都有显著的差异[19],棉铃虫Helicoverpa armigera Hübner的羽化节律严格受到光周期的调控,无论光暗期如何调换,其均在暗期羽化[20]。光周期还对迁飞性昆虫的生殖能力有影响,短光照(L∥D=8 h∥16 h)和中等光照(L∥D=12 h∥12 h)条件下,黏虫Mythimna seperata Walker的产卵前期显著延长[21]。
目前对环境条件影响草地贪夜蛾生长发育的研究主要集中在温度、寄主等环境因子上[22-24]。关于光周期对草地贪夜蛾生长发育影响的研究,国内外还少有报道。因此研究不同光周期对草地贪夜蛾生长发育和生殖的影响具有重要意义。本文通过探索不同光周期对草地贪夜蛾生长发育和生殖的影响,建立种群生命表并对种群动态进行预测,旨在揭示我国不同纬度不同季节所对应的不同光周期条件下,草地贪夜蛾的生长发育、生殖以及发生为害情况。确定最适合其生长发育的光周期条件,为预测不同季节草地贪夜蛾在我国的分布提供理论依据。
1 材料与方法
1.1 供试虫源
本试验所用的草地贪夜蛾于2019年5月采自广西南宁野外玉米田(108.37°E, 22.82°N),在实验室繁殖后进行试验。室内饲养条件为:温度(25±1)℃、相对湿度(60±5)%、幼虫密度1头/孔(12孔板:南通健格实验器材有限公司)。幼虫用棉铃虫人工饲料饲养[25-26],在相对湿度15%的灭菌土里化蛹,成虫用10%(V/V)蜂蜜水作为补充营养。
1.2 试验设计与方法
设置5个光周期,分别为L∥D=16 h∥8 h、L∥D=14 h∥10 h、L∥D=12 h∥12 h、L∥D=10 h∥14 h和L∥D=8 h∥16 h。幼虫老熟时,在6孔板中放入相对湿度15%的灭菌土,让其化蛹,化蛹第4天,先用分析天平(METTLER TOLEDO,ER-182A)称蛹重,之后将蛹放回灭菌土中,等待其羽化。
幼虫龄期和蛹期的观察: 幼虫单独饲养,每天观察幼虫龄期,隔24 h统计1次,从化蛹到羽化所经历的时间记为蛹期。
成虫生殖的观察:待草地贪夜蛾羽化后,按照雌雄1∶1配对,放入直径5 cm、高12 cm的透明塑料筒中,每日用10%(V/V)蜂蜜水饲喂,在温度(25±1)℃、相对湿度(65±5)%,和幼虫期一致的光周期条件下饲养。每天清点并取出纱布或塑料筒上面的卵,记录产卵日期、产卵量,待其死亡后,记录死亡日期,并解剖雌虫,根据精珠数量确定交配次数[27]。其中,将产卵整齐度(period of first oviposition, PFO)定义为各个处理每个雌虫的产卵前期与该处理中最短产卵前期的差值[28-29]。
1.3 生命表参数
试验得到的草地贪夜蛾平均世代周期(T),净增殖率(R0),内禀增长率(r)和周限增长率(λ)用以下公式计算:
式中Sxj表示草地贪夜蛾从卵发育到年龄x及龄期j的概率,lx表示草地贪夜蛾种群在年龄x时的存活率,fxj表示草地贪夜蛾雌虫在年龄x及龄期j时的产卵量,mx表示草地贪夜蛾种群在年龄x时的平均产卵量,lxmx表示草地贪夜蛾种群在年龄x时的净繁殖力。
1.4 草地贪夜蛾种群动态预测
利用软件 TWOSEX-MSChart[30]得到生命表数据,使用 TIMING-MSChart[31]软件预测 5 个光周期条件下草地贪夜蛾未来 60 d的种群动态。
1.5 数据分析
本试验得到的数据用SPSS 23.0进行统计分析,用Origin Pro 9.1进行图形绘制,分析结果用平均值±标准误表示。对不同光周期下的生长发育及生殖参数进行单因素方差分析,对于差异性显著的数据用Tukey’s HSD法进行多重比较,显著性水平设置为0.05。相同处理不同性别之间的比较用独立样本t测验,显著性水平设置为0.05。交配率和各阶段存活率结果用卡方检验进行分析。
2 结果与分析
2.1 不同光周期对草地贪夜蛾生长发育的影响
2.1.1 不同光周期下草地贪夜蛾各阶段的发育历期
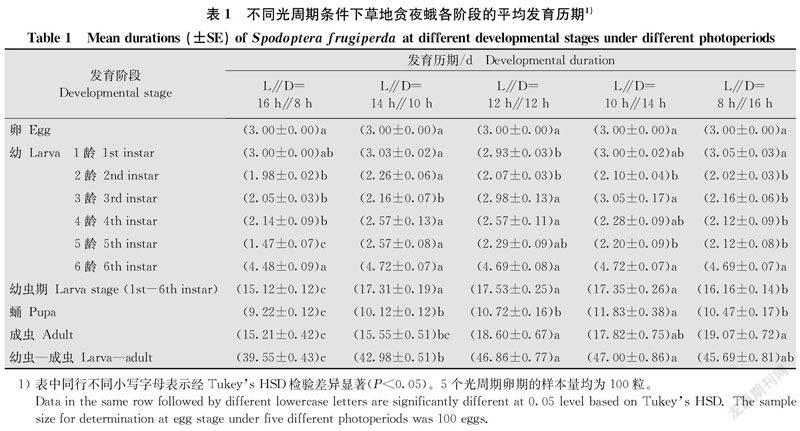
草地贪夜蛾在所设光周期下均能完成世代发育。光周期除了对卵和6龄幼虫的发育历期没有显著影响(P>0.05)之外,对其他各阶段发育历期均有显著影响(1龄:F4,287=3.39, P<0.05;2齡:F4,287=7.85, P<0.05;3龄:F4,287=21.11, P<0.05;4龄:F4,287=4.56, P<0.05;5龄:F4,287=26.04, P<0.05;1~6龄:F4,287=26.37, P<0.05;蛹:F4,287=19.67, P<0.05;成虫:F4,287=7.83, P<0.05;幼虫-成虫:F4,287=20.41, P<0.05)。
整体而言,随着光照时间的增加,草地贪夜蛾各阶段发育历期先延长后缩短,草地贪夜蛾幼虫在L∥D=16 h∥8 h时发育最快(15.12 d),蛹期和成虫寿命最短,分别为9.22 d和15.21 d。L∥D=12 h∥12 h下幼虫发育最慢(17.53 d),L∥D=10 h∥14 h的蛹期最长(11.83 d),L∥D=8 h∥16 h下成虫寿命最长(19.07 d)。在L∥D=10 h∥14 h条件下,草地贪夜蛾从1龄幼虫发育到成虫死亡所经历的时间最长(47.00 d),L∥D=16 h∥8 h条件下经历的时间最短(39.55 d)(表1)。
对草地贪夜蛾在同一光周期条件下相同龄期不同性别的发育历期进行独立样本t测验,结果表明,相同光周期条件下,卵期、1~4龄的雌、雄幼虫历期无显著差异(P>0.05),L∥D=12 h∥12 h条件下,5龄雄虫历期显著高于雌虫(t1,56=-2.78, P<0.05)。L∥D=16 h∥8 h(t1,56=-6.20, P<0.05), L∥D=14 h∥10 h(t1,56=-5.25, P<0.05), L∥D=12 h∥12 h(t1,56=-5.66, P<0.05)和L∥D=8 h∥16 h(t1,56=-9.28, P<0.05)的雄蛹蛹期均显著高于雌蛹的蛹期。雄蛾寿命在L∥D=12 h∥12 h(t1,56=-2.30, P<0.05)和L∥D=8 h∥16 h(t1,56=-3.33, P<0.05)时显著高于雌蛾寿命。草地贪夜蛾雄性幼虫-成虫历期在L∥D=12 h∥12 h(t1,56=-3.70, P<0.05), L∥D=10 h∥14 h(t1,56=-2.11, P<0.05)和L∥D=8 h∥16 h(t1,56=-4.44, P<0.05)时显著高于雌性幼虫-成虫的历期(表2)。
2.1.2 光周期对草地贪夜蛾蛹重和性比的影响
光周期对草地贪夜蛾雌蛹(F4,221=17.22,P<0.05)、雄蛹(F4,214=16.70,P<0.05)以及雌雄蛹的重量(F4,440=32.39,P<0.05)均有显著影响(表3)。在L∥D=14 h∥10 h时草地贪夜蛾的雌蛹(P<0.05)和雌雄平均蛹重(P<0.05)均最重,均显著高于L∥D=16 h∥8 h和L∥D=10 h∥14 h (P<0.05),但和L∥D=12 h∥12 h,L∥D=8 h∥16 h差异不显著(P>0.05)。L∥D=16 h∥8 h和L∥D=10 h∥14 h处理的雌蛹最轻(0.22 g),显著低于L∥D=14 h∥10 h的处理(P<0.05),与L∥D=12 h∥12 h和L∥D=8 h∥16 h 的处理差异不显著(P>0.05)。L∥D=10 h∥14 h和L∥D=16 h∥8 h条件下雄蛹的重量最轻(0.23 g),显著低于L∥D=14 h∥10 h和L∥D=12 h∥12 h的处理(P<0.05),与L∥D=8 h∥16 h的处理差异不显著(P>0.05)。
光周期对草地贪夜蛾的雌、雄性比没有显著影响(P>0.05),且各个光周期条件下草地贪夜蛾的雌雄性比均接近1∶1,其中 L∥D=10 h∥14 h时草地贪夜蛾的雌雄性比最高(1.27),L∥D=12 h∥12 h时草地贪夜蛾的雌雄性比最低(0.88)。
2.2 不同光周期对草地贪夜蛾生殖的影响
2.2.1 产卵前期
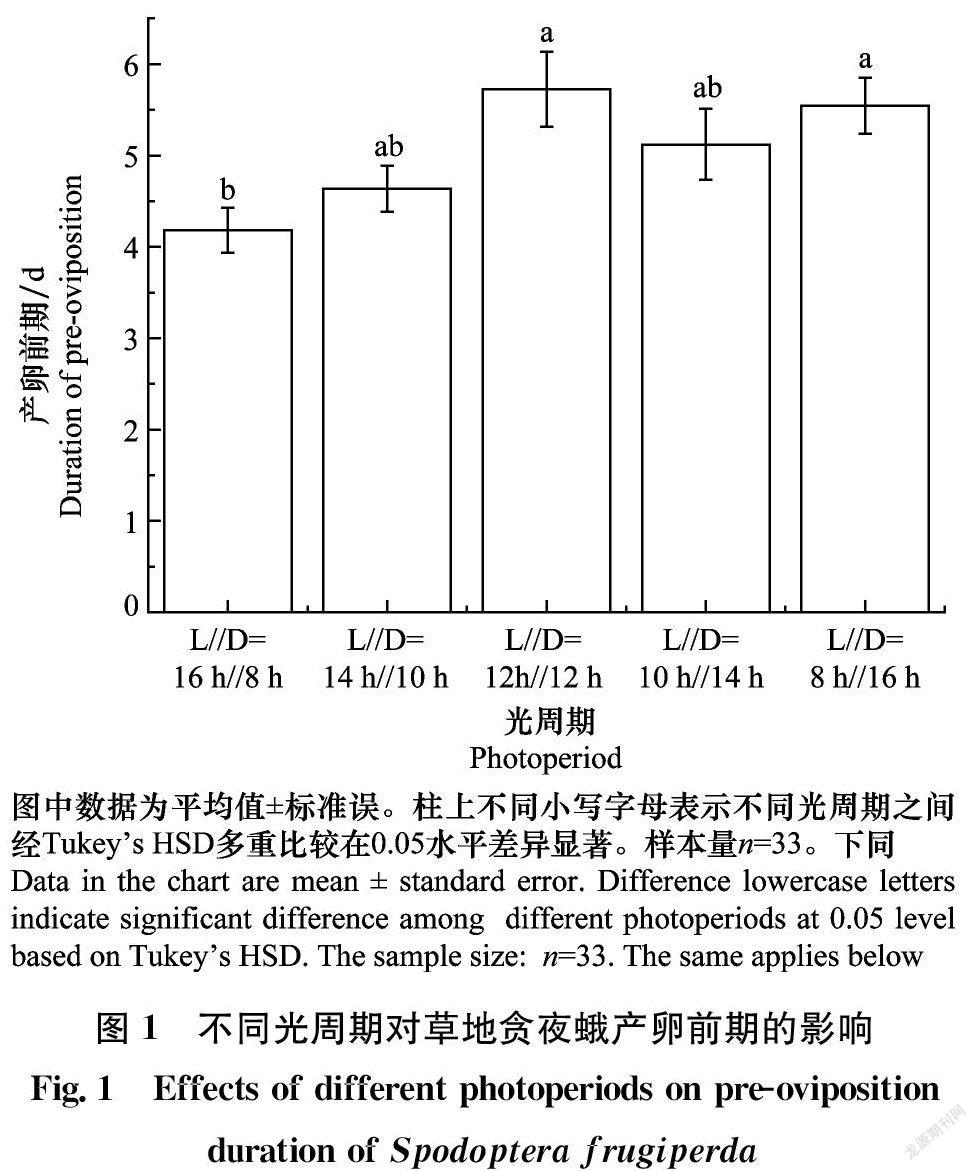
不同光周期条件下,草地贪夜蛾的产卵前期差异显著(F4,160=3.81,P<0.05;图1)。L∥D=12 h∥12 h时,雌蛾的产卵前期最长(5.73 d),显著高于L∥D=16 h∥8 h (P<0.05),但与另外3个光周期处理差异不显著(P>0.05)。L∥D=16 h∥8 h的产卵前期最短(4.18 d),显著低于L∥D=12 h∥12 h 和L∥D=8 h∥16 h处理(P<0.05),但与L∥D=14 h∥10 h (4.64 d)和L∥D=10 h∥14 h(5.12 d)处理组差异不显著(P>0.05)。
2.2.2 产卵整齐度
不同光周期对草地贪夜蛾的产卵整齐度有显著影响(F4,160=2.82,P<0.05;图2),其中在L∥D=10 h∥14 h时,草地贪夜蛾的PFO最大(2.82 d),显著高于L∥D=14 h∥10 h的PFO(P<0.05),与L∥D=16 h∥8 h、L∥D=12 h∥12 h和L∥D=8 h∥16 h之间的PFO没有显著差异(P>0.05),L∥D=14 h∥10 h时草地贪夜蛾的PFO最小(1.42 d),显著低于L∥D=10 h∥14 h(P<0.05),与L∥D=16 h∥8 h(2.18 d)、L∥D=12 h∥12 h(2.73 d)和L∥D=8 h∥16 h(2.55 d)处理组的PFO无显著差异(P>0.05)。
2.2.3 产卵历期、总产卵量、交配次数与交配率
草地贪夜蛾在不同光周期条件下的产卵历期显著不同(F4,160=3.94,P<0.05;表4),L∥D=14 h∥10 h时草地贪夜蛾的产卵历期最长,为7.48 d,显著高于L∥D=12 h∥12 h(5.64 d)(P<0.05),但两者均与L∥D=16 h∥8 h(6.58 d)、L∥D=10 h∥14 h(6.39 d)和L∥D=8 h∥16 h(6.67 d)处理组的产卵历期差异不显著(P>0.05)。
不同光周期条件下草地贪夜蛾的总产卵量差异显著(F4,160=13.01,P<0.05;表4),其中L∥D=14 h∥10 h时,草地贪夜蛾的产卵量最多,为1 950.64粒,显著高于L∥D=16 h∥8 h(1 516.85粒)、L∥D=12 h∥12 h(1 198.15粒)和L∥D=10 h∥14 h(1 328.91粒)处理(P<0.05),与L∥D=8 h∥16 h处理(1 758.55粒)差异不显著(P>0.05)。在L∥D=12 h∥12 h时,草地贪夜蛾的总产卵量最低,显著低于L∥D=14 h∥10 h和L∥D=8 h∥16 h处理(P<0.05),但与其余光周期处理差异不显著(P>0.05)。
光周期对草地贪夜蛾的交配次数(F4,160=8.72,P<0.05)和交配率(χ2=14.66,P<0.05)均有显著影响。L∥D=14 h∥10 h处理组的交配次数和交配率最高,分别为3.55次和95.25%,其中,其交配次数显著高于L∥D=12 h∥12 h、L∥D=10 h∥14 h和L∥D=8 h∥16 h处理(P<0.05),但与L∥D=16 h∥8 h处理差异不显著(P>0.05)。其交配率显著高于L∥D=12 h∥12 h和L∥D=10 h∥14 h处理(P<0.05),但与L∥D=16 h∥8 h和L∥D=8 h∥16 h处理差异不显著(P>0.05)(表4)。
圖3中的年龄-阶段特征存活率(Sxj)表示草地贪夜蛾从卵发育到年龄x及龄期j的概率。试验结果表明,不同光周期条件下草地贪夜蛾的年龄-阶段特征存活率存在显著差异。不同光周期下草地贪夜蛾从卵发育到1龄幼虫的概率存在显著差异(P<0.05),其中L∥D=16 h∥8 h条件下卵的孵化率最高(76.94%),显著高于L∥D=14 h∥10 h(64.93%)和L∥D=10 h∥14 h(64.93%)(P<0.05),与L∥D=12 h∥12 h(74.07%)以及L∥D=8 h∥16 h(69.91%)差异不显著(P>0.05)。在L∥D=16 h∥8 h条件下,草地贪夜蛾从卵发育到成虫的概率最高(36.00%),显著高于L∥D=14 h∥10 h(29.00%),L∥D=12 h∥12 h(27.00%),L∥D=10 h∥14 h(26.00%)和L∥D=8 h∥16 h(29.00%)。
5个光周期lx曲线的变化趋势基本相同,都是前期突降,中期平稳,后期突降的趋势,表明5个光周期条件下种群死亡主要集中在幼虫低龄期(图4)。5个光周期繁殖力曲线fxj开始出现的时间点依次为L∥D=16 h∥8 h为第28天,L∥D=14 h∥10 h为第31天,L∥D=12 h∥12 h为第33天,L∥D=10 h∥14 h为第30天,L∥D=8 h∥16 h为第31天。其中L∥D=16 h∥8 h的繁殖力曲线出现得最早,L∥D=12 h∥12 h出现得最晚,表明在L∥D=16 h∥8 h时,草地贪夜蛾种群进入繁殖期最快,而L∥D=12 h∥12 h条件下进入繁殖期最慢。不同光周期下草地贪夜蛾种群进入繁殖高峰的时间不同,L∥D=16 h∥8 h是第32天,L∥D=14 h∥10 h为第35天,L∥D=12 h∥12 h为第37天,L∥D=10 h∥14 h为第36天,L∥D=8 h∥16 h为第34天。在繁殖高峰的平均产卵量也不同,L∥D=14 h∥10 h时草地贪夜蛾的产卵量最高(314.79粒),L∥D=10 h∥14 h时产卵量最低(101.85粒),L∥D=16 h∥8 h、L∥D=12 h∥12 h和L∥D=8 h∥16 h的产卵量依次为268.79、111.08粒和288.63粒。在L∥D=14 h∥10 h条件下草地贪夜蛾的mx,lxmx曲线均高于其余光周期,表明在此条件下,草地贪夜蛾的繁殖力最强,而L∥D=12 h∥12 h时,草地贪夜蛾的mx,lxmx相对最低,因此这个条件不适合草地贪夜蛾繁殖。
2.4 不同光周期对草地贪夜蛾种群生命表参数的影响
草地贪夜蛾种群的平均世代周期(T)在L∥D=16 h∥8 h条件下最短(33.10 d),显著低于其他4个光周期处理组(P<0.05),在L∥D=12 h∥12 h和L∥D=10 h∥14 h条件下最长,分别为38.27 d和38.49 d。在L∥D=14 h∥10 h条件下,草地贪夜蛾的种群净增殖率(R0)最高(525.08),显著高于L∥D=12 h∥12 h(199.96)和L∥D=10 h∥14 h(186.04)(P<0.05),但与L∥D=16 h∥8 h(507.32)和L∥D=8 h∥16 h(463.07)差异不显著(P>0.05)。在L∥D=16 h∥8 h条件下,草地贪夜蛾的种群内禀增长率(r)(0.19)和周限增长率(λ)(1.21)均最高,显著高于其余光周期(P<0.05),在L∥D=12 h∥12 h和L∥D=10 h∥14 h条件下,草地贪夜蛾的种群内禀增长率(r)(0.14)和周限增长率(λ)(1.15)最低(表5)。
2.5 不同光周期下草地贪夜蛾种群动态预测
利用TIMING-MSChart 軟件对草地贪夜蛾未来60 d的种群动态进行预测,结果表明,在L∥D=16 h∥8 h条件下,草地贪夜蛾的种群发育最快,60 d后已经发育到第3代的卵期,而在另外4个光周期下均发育到第2代的蛹期(图5)。60 d后,不同光周期条件下草地贪夜蛾当代的种群数量也明显不同,L∥D=16 h∥8 h条件下草地贪夜蛾的三代卵已达到4.28个数量单位,这一时期的第二代的蛹(3.26)和雌、雄蛾(3.03)也占很大比例;L∥D=14 h∥10 h条件下草地贪夜蛾的蛹达到3.42个数量单位,6龄幼虫为2.74个数量单位;L∥D=12 h∥12 h时草地贪夜蛾的蛹为2.88个数量单位,6龄幼虫为2.62个数量单位;L∥D=10 h∥14 h时草地贪夜蛾的蛹为2.78个数量单位,6龄幼虫为2.54个数量单位;L∥D=8 h∥16 h时草地贪夜蛾的蛹为3.38个数量单位,6龄幼虫为2.39个数量单位。可以看出,虽然L∥D=14 h∥10 h条件下,草地贪夜蛾种群还未发育到第3代,但是此时二代蛹和6龄幼虫的数量单位均最高,因此此时草地贪夜蛾有最高的繁殖潜力。
3 讨论
本研究结果表明,光周期对草地贪夜蛾各虫态的发育历期均有显著影响,在光周期为L∥D=12 h∥12 h时,草地贪夜蛾的幼虫历期最长,雄性幼虫到成虫的历期也是最长,这与黏虫相似,黏虫在L∥D=12 h∥12 h时世代发育历期也是最长[21]。光周期对草地贪夜蛾的幼虫阶段各龄期死亡率、雌雄性比没有显著影响,但对草地贪夜蛾蛹期的死亡率有显著影响,在长光照(L∥D=16 h∥8 h和L∥D=14 h∥10 h)时,草地贪夜蛾蛹的羽化率最高,在相对短光照(L∥D=12 h∥12 h和L∥D=10 h∥14 h)时,草地贪夜蛾蛹的羽化率最低,而棉铃虫在相对短光照L∥D=12 h∥12 h时羽化率最高[20]。光周期对草地贪夜蛾雌雄蛹的重量有显著影响,L∥D=14 h∥10 h的雌蛹和雄蛹最重,这一结果也与棉铃虫类似[32],但黏虫在L∥D=12 h∥12 h时蛹最重[31]。光周期作为一种信号,其调节昆虫生长发育肯定有相应的信号传递途径,最近有研究指出,光周期能够通过调节直翅目蟋蟀Modicogryllus siamensis体内保幼激素的含量来影响蟋蟀的发育[33],其结果与本文结果一致,即短光照(L∥D=12 h∥12 h)下幼虫的发育历期比长光照(L∥D=16 h∥8 h)长,在草地贪夜蛾体内是否也存在这样的调节路径,值得进一步探究。草地贪夜蛾在短光照时的羽化率比长光照低,也许就是因为短光照促进保幼激素的释放,使草地贪夜蛾的蛹不能正常羽化,这一点类似于滞育昆虫的滞育,这一猜想还有待进一步研究证实。
光周期对草地贪夜蛾的生殖能力影响结果表明,不同光周期对迁飞性昆虫的生殖能力也有较大影响,从本文的研究结果可以看出,光周期对草地贪夜蛾的交配次数和交配率均有显著影响,其中两者在L∥D=14 h∥10 h时最高,在L∥D=12 h∥12 h时最低,棉铃虫的交配次数和交配率在L∥D=16 h∥8 h最高,L∥D=12 h∥12 h最低,与本研究类似[20]。有研究表明,光周期可以显著影响昆虫性信息素各组分的含量,比如光周期越短,棉铃虫性信息素各组分的含量越低[34],因此推测光周期能够调节草地贪夜蛾性信息素的释放,进而调节草地贪夜蛾的性行为,但还需要进一步的研究来验证。光周期对草地贪夜蛾的产卵前期、产卵历期、总产卵量均有显著影响,其中光周期为L∥D=12 h∥12 h时的产卵前期最长,这一光周期条件下的产卵历期和总产卵量相较于其余光周期也较弱,这一结果与黏虫的研究结果类似[31],而棉铃虫在短光照L∥D=12 h∥12 h和L∥D=8 h∥16 h时产卵前期最短,产卵量最高[20]。光周期对草地贪夜蛾的产卵整齐度也具有显著性影响,L∥D=10 h∥14 h下草地贪夜蛾的产卵整齐度最低,此时草地贪夜蛾的种群生殖期不集中,不具备暴发的潜力;而L∥D=14 h∥10 h的产卵整齐度最高,此时草地贪夜蛾能够集中产卵,下一代能够集中为害,具备暴发潜力,而且在5月-7月,我国大部分地区的光周期均接近L∥D=14 h∥10 h,因此在此期间要做好草地贪夜蛾的监测与防控工作,防止其在此时形成较大的种群。
不同光周期條件下,草地贪夜蛾的年龄-特征存活率和繁殖力不同,存活率的差异主要存在于卵期,L∥D=16 h∥8 h条件下草地贪夜蛾卵的孵化率最高,L∥D=14 h∥10 h最低,然而L∥D=14 h∥10 h的繁殖力却最高,造成这一现象的原因也许是草地贪夜蛾在L∥D=14 h∥10 h条件下通过繁殖力的增强来弥补卵孵化率的不足。一个种群增殖能力的强弱,可以通过净增殖率、内禀增长率和周限增长率来反映[34]。草地贪夜蛾在L∥D=16 h∥8 h条件下内禀增长率和周限增长率均最高,而净增殖率则低于L∥D=14 h∥10 h;在短光照L∥D=12 h∥12 h和L∥D=10 h∥14 h时,草地贪夜蛾的净增殖率、内禀增长率和周限增长率均最低,这一结果表明长光照更有利于草地贪夜蛾种群的繁殖。对草地贪夜蛾未来60 d的种群预测也有类似规律,即长光照种群的繁殖速度最快。
根据本文结果,在L∥D=14 h∥10 h时,草地贪夜蛾种群的增殖能力最强,而在L∥D=12 h∥12 h时,草地贪夜蛾的繁殖能力最低。L∥D=12 h∥12 h只有在3月的春分日和9月的秋分日会出现,而这两个时间点恰恰是北半球气候转变的关键时间点,有研究表明草地贪夜蛾在3月会随盛行西风向我国东北方向迁飞[14],根据迁飞昆虫飞行和生殖互作关系的研究[35-37],生殖能力较强的个体,其飞行能力会偏弱,结合本文结果,推测L∥D=12 h∥12 h条件下草地贪夜蛾的飞行能力会很强。而L∥D=14 h∥10 h这一光周期现象在我国中部地区是普遍存在的,从4月到7月之间,我国各地均会出现这一光周期,因此在此条件下,草地贪夜蛾更倾向于在本地繁殖为害。具体光周期对草地贪夜蛾飞行能力的影响有待进一步研究。
参考文献
[1] LUGINBILL P. The fall armyworm [R]. USDA Technology Bulletin, 1928: 34-91.
[2] SPARKS A N. A review of the biology of the fall armyworm [J]. The Florida Entomologist, 1979, 62(2): 82-87.
[3] TODD E L, POOLE R W. Keys and illustrations for the armyworm moths of the noctuid genus Spodoptera Guenée from the Western Hemisphere [J]. Annals of the Entomological Society of America, 1980, 73(6): 722-738.
[4] EARLY R, GONZLEZ-MORENO P, MURPHY S T, et al. Forecasting the global extent of invasion of the cereal pest Spodoptera frugiperda, the fall armyworm [J]. NeoBiota, 2018, 40: 25-50.
[5] JOHNSON S J. Migration and the life history strategy of the fall armyworm, Spodoptera frugiperda in the Western Hemisphere [J]. International Journal of Tropical Insect Science, 1987, 8(4/5/6): 543-549.
[6] PASHLEY D P. Host-associated genetic differentiation in fall army worm (Lepidoptera: Noctuidae): a sibling species complex?[J]. Annals of the Entomological Society of America, 1986, 79(6): 898-904.
[7] CASMUZE A, JUREZ M L, SOCAS M G, et al. Revisión de los hospederos del gusano cogollero del maíz, Spodoptera frugiperda (Lepidoptera: Noctuidae) [J]. Revista de la Sociedad Entomológical Argentina, 2010, 69: 209-231.
[8] GOERGEN G, KUMAR P L, SANKUNG S B, et al. First report of outbreaks of the fall armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa [J/OL]. PLoS ONE, 2016, 11(10): e0165632. DOI:10.1371/journal.pone.0165632.
[9] COCK M J W, BESEH P K, BUDDIE A G, et al. Molecular methods to detect Spodoptera frugiperda in Ghana, and implications for monitoring the spread of invasive species in developing countries [J/OL]. Scientific Reports, 2017, 7(1): 4103. DOI:10.1038/s41598-017-04238-y.
[10]NAGOSHI R N, GOERGEN G, TOUNOU K A, et al. Analysis of strain distribution, migratory potential, and invasion history of fall armyworm populations in northern Sub-Saharan Africa [J/OL]. Scientific Reports, 2018, 8(1): 3710. DOI:10.1038/s41598-018-21954-1.
[11]RWOMUSHANA I, BATEMAN M, BEALE T, et al. Fall armyworm: impacts and implications for Africa [R]. Evidence Note Update, October 2018. Report to DFID. Wallingford, UK: CAB International, 2018.
[12]STOKSTAD E. New crop pest takes Africa at lightning speed [J]. Science, 2017, 356(6337): 473-474.
[13]SHARANABASAPPA D, KALLESHWARASWAMY C M, ASOKAN R, et al. First report of the fall armyworm, Spodoptera frugiperda (J E Smith) (Lepidoptera: Noctuidae), an alien invasive pest on maize in India [J]. Pest Management in Horticultuml Ecosystems, 2018, 24(1): 23-29.
[14]吴秋琳, 姜玉英, 胡高, 等.中国热带和南亚热带地区草地贪夜蛾春夏两季迁飞轨迹的分析[J]. 植物保护, 2019, 45(3): 1-9.
[15]姜玉英, 刘杰, 朱晓明.草地贪夜蛾侵入我国的发生动态和未来趋势分析[J]. 中国植保导刊, 2019, 39(2): 33-35.
[16]吴少会, 向群, 薛芳森. 昆虫的行为节律[J].江西植保, 2006, 29(4): 147-157.
[17]童廖. 温度和光周期对美国白蛾羽化和交配影响[D].南昌: 江西农业大学, 2012.
[18]黄少虹, 江幸福, 罗礼智.光周期和温度对草地螟滞育诱导的影响[J].昆虫学报, 2009, 52(3): 274-280.
[19]何海敏, 杨慧中, 肖亮, 等.温度和光周期对甜菜夜蛾发育历期和繁殖的影响[J].江西植保, 2011, 34(3): 93-96.
[20]孙晓婷, 徐瑞斌, 葛世帅, 等.光周期对棉铃虫羽化、生殖和飞行的影响[J].环境昆虫学报, 2019, 41(5): 1045-1056.
[21]王熠, 庞士海, 纪薇, 等. 光周期对黏虫生长发育和生殖的影响[J].植物保护学报, 2019, 46(3): 542-548.
[22]鲁智慧, 和淑琪, 严乃胜, 等. 温度对草地贪夜蛾生长发育及繁殖的影响[J].植物保护, 2019, 45(5): 27-31.
[23]谢殿杰, 张蕾, 程云霞, 等. 不同温度下草地贪夜蛾年龄-阶段实验种群两性生命表的构建[J].植物保护, 2019, 45(6): 20-27.
[24]吴正伟, 师沛琼, 曾永辉, 等. 3种寄主植物饲养的草地贪夜蛾种群生命表[J].植物保护, 2019, 45(6): 59-64.
[25]梁革梅, 谭维嘉, 郭予元. 人工饲养棉铃虫技术改进[J].植物保护, 1999, 25(2): 15-17.
[26]李俊, 谭显胜, 谭泗桥, 等. 改进支持向量机在棉铃虫人工饲料配方优化中的应用[J].昆虫学报, 2010, 53(4): 420-426.
[27]吕伟祥, 江幸福, 张蕾, 等.一日龄粘虫不同时长吊飞对生殖及寿命的影响[J].应用昆虫学报, 2014, 51(4): 914-921.
[28]ZHANG Lei, PAN Pan, SAPPINGTON T W, et al. Accelerated and synchronized oviposition induced by flight of young females may intensify larval outbreaks of the rice leaf roller [J/OL]. PLoS ONE, 2015, 8(5): e63554. DOI:10.1371/journal.pone.0121821.
[29]CHENG Yunxia, LUO Lizhi, JIANG Xingfu, et al. Synchronized oviposition triggered by migratory flight intensifies larval outbreaks of beet webworm [J/OL]. PLoS ONE, 2012, 7(2): e31562. DOI: 10.1371/journal.pone.0031562.
[30]CHI H. TWOSEX-MSChart: a computer program for the age-stage, two-sex life table analysis [OL].http:∥140.120.197.173/Ecology/Download/TWOSEX.zip.
[31]CHI H. TIMING-MSChart: a computer program for the age-stage, two-sex life table analysis [OL].http:∥140.120.197.173/Ecology/ Download/TIMING.zip.
[32]陳元生, 段德康, 陈超, 等. 光周期和温度对棉铃虫发育历期及蛹重的影响[J].环境昆虫学报, 2012, 34(4): 407-414.
[33]MIKI T, SHINOHARA T, CHAFINO S, et al. Photoperiod and temperature separately regulate nymphal development through JH and insulin/TOR signaling pathways in an insect [J]. Proceedings of the National Academy of Sciences, 2020, 117(10): 5525-5531.
[34]鲁玉杰,张孝羲,翟保平. 温度和光周期对棉铃虫雌性信息素成分的含量与比例的影响[J].生态学报,2002,22(4):566-570.
[35]葛世帅. 草地贪夜蛾的飞行能力及其与生殖的互作关系研究[D].福州:福建农林大学, 2020.
[36]郑作涛. 二点委夜蛾飞行与生殖的互作[D].北京:中国农业科学院,2014.
[37]潘攀. 稻纵卷叶螟飞行行为及与生殖的互作关系[D].北京:中国农业科学院, 2013.
(责任编辑:杨明丽)
