类胡萝卜素代谢途径基因变异导致园艺植物色泽差异的研究进展
2022-06-06曾旭梅席婉朱琳琳袁金梅熊康舜王彩云郑日如
曾旭梅,席婉,朱琳琳,袁金梅,熊康舜,王彩云,郑日如
园艺植物生物学教育部重点实验室/华中农业大学园艺林学学院,武汉 430070
类胡萝卜素是自然界中广泛分布的一类次生代谢物,在植物的生长发育中发挥着重要的作用[1]。同时,在植物花瓣、果实和叶片等器官中由于类胡萝卜素种类及含量的差异,导致色泽的多样性[2-5],形成五彩缤纷的世界(图1)。此外,类胡萝卜素可裂解产生紫罗酮等重要香气物质[6]。对植物而言,鲜艳的颜色和芬芳的气味可有效吸引昆虫或其他动物为其传粉,提高生殖成功率[7]。在植物光合机构中,类胡萝卜素能辅助捕获光能并传递给叶绿素进行光合作用;也能有效清除自由基,避免光氧化对植物造成的损伤[8]。另外,类胡萝卜素的下游产物脱落酸(abscisic acid,ABA)和独角金内酯(strigolactone)是植物抵御逆境、调控生长发育等重要的内源信号物质[9]。类胡萝卜素也是一类具有抗氧化能力的高价值天然活性物质,是人体膳食营养的重要组成部分[10]。因此,类胡萝卜素代谢途径的分子机制一直是园艺植物研究的热点。

图1 含类胡萝卜素的常见园艺植物[2-5]Fig.1 Common horticultural plants containing carotenoids
在园艺植物中,类胡萝卜素的代谢通路已基本明晰。近年来,随着组学技术的不断发展以及与遗传学的深度融合,越来越多类胡萝卜素代谢途径的关键基因及其重要变异被挖掘并加以验证。这些研究表明,代谢途径中结构基因编码区和启动子的变异会导致基因功能及转录水平的显著变化,从而改变类胡萝卜素的种类和含量。分子标记辅助育种是未来种业的发展方向,解析优良性状形成的分子机制是开展定向改良的核心科学问题,开发可用于早期选择的分子标记是实现高效育种的关键技术手段。因此,本文总结了植物类胡萝卜素代谢通路、基因变异研究策略、基因变异对类胡萝卜素代谢的影响及转录调控的研究,以期加深对园艺植物类胡萝卜素多样性的生物学认识,同时为开发分子标记、提升育种效率提供理论依据。
1 园艺植物类胡萝卜素的多样性
类胡萝卜素的种类异常丰富,迄今为止已在植物、动物和微生物中发现800余种。它是一类含有40个碳的类异戊烯聚合物,由8个异戊二烯单位首尾相连形成。类胡萝卜素类物质主要分为两大类,一是只含碳氢元素的胡萝卜素类,包括番茄红素、八氢番茄红素、α- 胡萝卜素、β-胡萝卜素等物质;二是胡萝卜素类的含氧衍生物(即叶黄素类),包括叶黄素、虾青素、玉米黄质等[11]。类胡萝卜素也因其分子结构中存在的共轭双键而呈现不同的颜色[12]。园艺植物中常见的类胡萝卜素有红色的辣椒红素、橙红色的β-胡萝卜素、粉色的番茄红素、橙黄色的紫黄质、黄色的黄体素和玉米黄质等[2,13-15]。不同种类和含量的类胡萝卜素积累可形成园艺植物中美丽多彩的色泽[3,16]。同时,在柑橘、辣椒、西瓜、番茄等同种植物的不同品种间也可因类胡萝卜素种类的不同而呈现色泽的差异(表1)。如柑橘中的‘温州蜜柑’果实积累β-隐黄质而呈橙红色,‘红夏橙’果实积累紫黄质而呈黄色[35];辣椒中红色果实主要积累辣椒红素,橙色果实主要积累辣椒红素、玉米黄质,黄色果实主要积累叶黄素和β-胡萝卜素[20-21]。此外,同一种植物也可积累不同含量的类胡萝卜素而显示色泽多样性。如桂花中α-胡萝卜素和β-胡萝卜素含量的高低是区分丹桂(橙红色)、金桂(橙黄色)、银桂(黄白色)的关键因素[26,36]。因此,类胡萝卜素的种类和含量是决定园艺植物色泽多样性的重要原因。
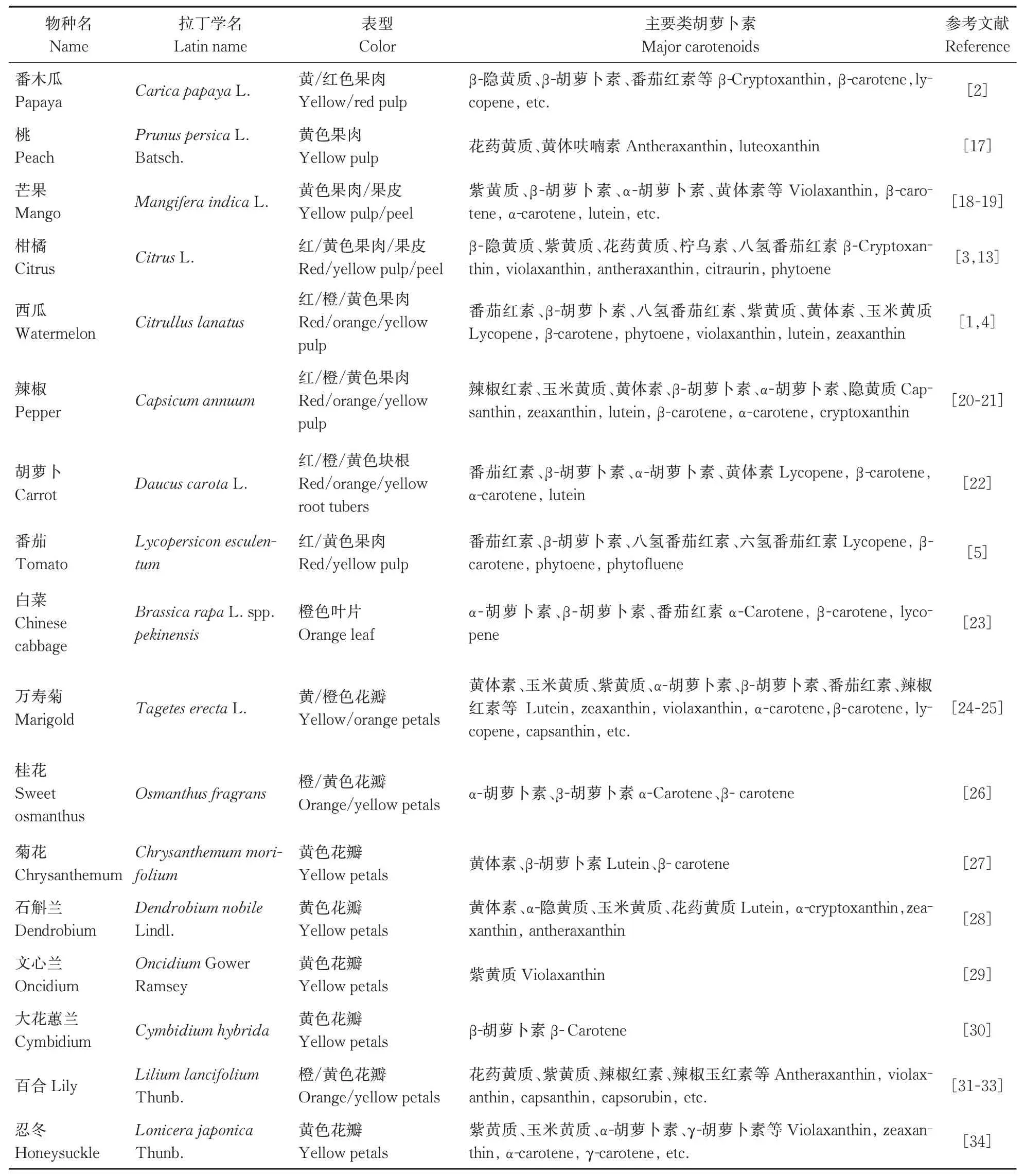
表1 园艺植物中的主要类胡萝卜素组分Table 1 Important carotenoids in different horticultural plants
2 植物类胡萝卜素的主要代谢通路
植物类胡萝卜素的代谢途径已被阐述清楚,主要包括合成及降解两个部分[37]。类胡萝卜素合成始于异戊二烯焦磷酸(IPP)与二甲基丙烯焦磷酸(DMAPP)的缩合,在牻牛儿牻牛儿基焦磷酸合酶(GGPPS)的作用下生成的前体物质——牻牛儿牻牛儿基焦磷酸(geranylgeranyl pyrophosphate,GGPP)。两分子的GGPP 通过八氢番茄红素合成酶(phytoene synthase,PSY)的作用产生无色的八氢番茄红素(phytoene),然后,在八氢番茄红素脱氢酶(phytoene desaturase,PDS)、δ-胡萝卜素异构酶(zeta-carotene isomerase,ZISO)、ζ-胡萝卜素脱氢酶(zeta-carotenedesaturase,ZDS)和胡萝卜素异构酶(carotenoid isomerase,CRTISO)等一系列酶的作用下生成番茄红素(lycopene)。随后,类胡萝卜素代谢途径被分为2 个分支点,由番茄红素ε-环化酶(lycopene epsilon cyclase,LCYe)和番茄红素β-环化酶(lycopene beta cyclase,LCYb)的环化分别产生α-胡萝卜素、β-胡萝卜素两个重要产物[38]。α-胡萝卜素在β-胡萝卜素羟化酶(β-carotene hydroxylase,BCH)和ε-胡萝卜素羟化酶(ECH)作用下生成叶黄素;而β-胡萝卜素在β-胡萝卜素羟化酶(BCH)、玉米黄质环氧化酶(zeaxanthin epoxidase,ZEP)、紫黄质脱环氧化酶(violaxanthin deepoxidase,VDE)、新黄质合成酶(neoxanthin synthase,NXS)等作用下,生成β-隐黄质、玉米黄质、新黄质等,其中新黄质是脱落酸(abscisic acid,ABA)的合成前体物质,也是β分支的最后一个产物。
植物类胡萝卜素的降解由酶促反应和非酶促反应完成,可生成多种脱辅基类胡萝卜素,如香气物质、激素、色素及信号分子等[9,39]。其中,酶促降解过程在植物中主要由类胡萝卜素裂解氧化酶(carotenoid cleavage oxygenases,CCOs)催化完成;非酶促降解过程通常与逆境胁迫应答相关。CCOs 可以特异性剪切C40类胡萝卜素多烯链的共轭双键,形成多种脱辅基类胡萝卜素,部分产物还可进一步裂解[12]。CCOs 家族具有双加氧酶特性,根据酶所裂解的底物是否发生环氧化,可将其分为类胡萝卜素裂解双加氧酶(carotenoid cleavage dioxygenases,CCDs)和 9-顺式-环氧类胡萝卜素双加氧酶(9-cis-epoxycarotenoid dioxygenases,NCEDs)[40]。在类胡萝卜素降解过程中,CCD1 和CCD4 在植物的色泽和香气物质代谢中发挥重要作用[6]。桂花、拟南芥、月季、矮牵牛等植物中,CCD1 和 CCD4 以 C40类胡萝卜素和 C27脱辅基类胡萝卜素为底物分解产生香叶基丙酮、β-紫罗兰酮、假紫罗酮、α-紫罗酮等 C13或 C14香气物质[41-46]。同时,类胡萝卜素被CCD4 所裂解产生胭脂素、β-柠乌素及β-citraurinene 等天然色素物质,使红木、柑橘等植物的种子或果实呈现诱人的红色[15,47-50]。此
外,植物激素ABA 和独角金内酯也是类胡萝卜素裂解的重要产物[9]。番茄、水稻、鳄梨、葡萄及草莓等植物中NCED基因参与到ABA 生物合成过程,导致果实内源ABA 含量变化,果实表皮呈现不同色泽[51-56]。
3 植物类胡萝卜素遗传变异的常用研究方法
为了揭示色泽的遗传变异规律,利用正向遗传学方法进行研究,一般采取以下3 种手段:其一是挖掘突变体材料,如在番茄[57-58]、桃[59]、白菜[60]等研究中通过比较野生型和突变型的差异锁定了关键变异基因。其二是用2 种不同表型的植株进行杂交产生性状分离,再结合组学等技术明确差异区域,筛选并验证关键基因[15,61-62]。如在柑橘的研究中,利用红橘(红果皮)和枳(黄果皮)F1代杂交子代混池RNA-seq 测序产生的SNP 进行混合样本分组分析(BSA,bulked segregant analysis),再结合分子标记进行精细定位,揭示出CCD4b对色泽变异起到关键作用[15]。在花椰菜的花色研究中,对白花和黄花杂交的F2分离群体进行了BSA 分析结合全基因组重测序,筛选到了关键变异基因CCD4[63]。其三是利用丰富的自然品种寻找普遍规律。如在菊花研究中发现CmCCD4a在黄色品种中无法表达,而在白色品种中大量表达,进一步通过RNAi 验证了CmCCD4a对类胡萝卜素降解起重要作用,是黄色和白色花瓣差异的关键[16]。在桃中也利用不同基因型种质资源筛选得到果肉色泽差异基因[59]。
4 类胡萝卜素代谢途径关键基因变异研究与应用
关键代谢通路基因的编码区及启动子变异是造成植物类胡萝卜素种类和含量差异的重要原因(表 2)。在花椰菜[63]、胡萝卜[72]及菊花[77]等植物中证实,重要基因编码区的变异可使基因功能和基因表达量发生变化,从而导致表型差异。PSY 是类胡萝卜素生物合成途径的限速酶,研究显示在木薯和辣椒中,PSY编码区多个 SNPs 和 InDels 的变异,可影响类胡萝卜素的合成,使其呈现丰富的色泽[71,73]。番木瓜中的红色果肉是由番茄红素环化酶基因LCYb2的编码区突变引起番茄红素大量积累而产生[68]。再如,桃子黄色与白色果实相比,CCD4编码区发生了多处SNPs、转座子及逆转座子插入、氨基酸替代等变化,从而导致酶活性丧失,类胡萝卜素积累[59,64-67]。油菜的黄色花瓣中,由于CCD4基因编码区插入了一个CACTA 类型转座子导致降解功能丧失,下游的紫黄质大量积累;而未发生转座子插入的白色花瓣中类胡萝卜素裂解产生大量香气物质紫罗酮[60]。拟南芥种子、甘蓝花瓣中CCD4编码区的SNPs 变异导致其裂解功能下降,类胡萝卜素含量升高[62,78]。

表2 植物类胡萝卜素代谢关键基因启动子和/或编码区变异的遗传学机制Table 2 Genetic mechanism of variation of promoter and/or coding regions of key genes of carotenoids metabolism in plants
除了编码区之外,启动子的变异则可直接影响基因的转录水平,从而改变类胡萝卜素代谢。如柑橘中的CCD4b基因启动子中增加了Harbinger 类型MITE 转座子。经烟草双荧光素酶和GUS 酶活性检测表明,MITE 转座子上的SNP2G(CAACTG)可增强启动子活性,促进CCD4b大量表达形成红皮柑橘中特有的红色β-柠乌素[15]。除此之外,还有部分植物类胡萝卜素代谢途径关键基因的编码区和启动子同时发生了变异,如白菜的类胡萝卜素异构酶基因CRTISO的编码区突变导致功能丧失,同时启动子的88 bp缺失导致调控它的转录因子及代谢通路其他基因表达均受影响,最终前番茄红素(prolycopene)在突变体中大量积累[60]。拟南芥种子中的CCD4[78]、ZEP[79]的启动子和编码区变异影响了种子类胡萝卜素的积累。玉米LCYe[75]的改变则直接影响了α-和β-分支的流向。大量研究表明,植物类胡萝卜素的种类及含量与相关基因表达量密切相关[1,9,12],而启动子活性直接调控基因表达,因此启动子变异是造成色泽差异的另一主因。
在深入解析类胡萝卜素变异的分子机制基础上,利用差异片段开发可用于早期选择的分子标记能有效提升育种效率。如在白菜的研究中发现,CRTISO启动子88 bp缺失可导致白色叶片突变为黄色叶片,由此开发了InDel 共显性分子标记Br-Pro-Indel,可有效鉴别叶色,缩短育种周期[60]。玉米中的类胡萝卜素含量与LCYe紧密相关,它的编码区变异可引起α-胡萝卜素、β-胡萝卜素、β-隐黄质等重要类胡萝卜素代谢变化。根据编码区变异片段设计特异引物,对后代进行PCR 扩增可快速鉴定变异并推断表型[75]。因此,对类胡萝卜素代谢通路基因变异的解析不仅可以丰富色泽多样性的生物学理论,还可有效设计用于定向改良的分子标记。
5 植物类胡萝卜素代谢途径中关键基因的转录调控
类胡萝卜素代谢调控一直是园艺植物研究的热点,而转录因子通过直接或间接调控植物类胡萝卜素代谢途径关键基因的表达量,影响类胡萝卜素的含量。在柑橘中,CsMADS5 既可直接结合PSY、PDS、LCYB1的启动子,从而激活其转录,也可与CsMADS6形成增强子复合物,协同促进类胡萝卜素的积累,调控果实着色[80];CsERF061 激活PSY1、PDS、CRTISO、LCYb1、BCH、ZEP、NCED3、CCD1和CCD4等9 个关键类胡萝卜素途径基因的启动子,参与类胡萝卜素代谢的多靶点调节[81]。在番茄果实中证实,SlBBX20 可通过直接结合PSY1启动子中G-box 基序激活类胡萝卜素生物合成关键酶基因的表达,从而导致类胡萝卜素水平升高[82];再如SlMYB72 与八氢番茄红素合成酶、胡萝卜素异构酶和番茄红素β 环化酶基因结合,调控类胡萝卜素含量,使番茄果实呈现不均匀着色[83]。同时,在猕猴桃[84]、番木瓜[85]等果实中,AdMYB7、CpMYB1/2分别调控类胡萝卜素的合成,影响果实的风味与色泽。转录因子也调节类胡萝卜素裂解酶基因,导致园艺植物色泽和香气物质不同。桂花ERF61[86]和WRKY3[87]与OfCCD4启动子上的作用元件结合促进类胡萝卜素裂解双加氧酶基因的表达,调节花瓣中挥发性香气物质(如β-紫罗兰酮)的合成,从而导致品种间色泽和香气的差异。葡萄MADS4 转录因子负调控VvCCD4b的表达,影响其主要香气成分(如β-大马士酮、β-紫罗兰酮及α-紫罗兰酮等)[88]。此外,番茄果实成熟过程中,研究显示RIN 也可激活与成熟相关的基因,间接促进类胡萝卜素的积累[89]。因此,转录因子调控类胡萝卜代谢通路的关键基因,对园艺植物色泽差异也具有重要作用。
6 展 望
园艺植物与人们生活密切相关,色泽改良一直是园艺植物的重要育种目标之一。在柑橘、桃、葡萄、番茄、油菜等园艺植物及作物中,学者们已经逐渐探明类胡萝卜素代谢途径关键基因变异的分子机制,并在此基础上开发了一系列辅助育种分子标记。分子标记辅助育种可直接针对特定农艺性状对大量杂交后代开展快速且精确的检测,大幅提高育种效率,具有重要的产业价值。
