内蒙古森林—草原交错带中野生东北马鹿冬季食物组成模式
2022-06-06周良俊王琳魏楷丽张明海张玮琪
周良俊 王琳 魏楷丽 张明海 张玮琪
(东北林业大学野生动物与自然保护地学院,哈尔滨 150040)
野生动物的食物具有多物种、多成分和多类型的特征,不同食物功用差异显著(Raubenheimer and Simpson, 2009),且食物的种类、质量和数量在时间和空间上多变。因此,动物在不同的时间段,选择合适的地点,采食所需的食物,并摄入一定的数量,经过体内一系列协同、拮抗作用,所食食物经同化和重新分配等代谢过程,以满足身体的营养需求,使其能够生存(Feltonet al.,2009, 2016; Raubenheimeret al., 2009; Sperfeldet al.,2012;Wamet al.,2018)。食物的处理和消化吸收过程会影响种群中的每个个体,从而使整个种群表现出特定的食物组成模式,且这种食物组成模式会在不同时间和不同环境因子(气候、海拔、纬度、水深等) 影响下发生变化(Graham and Vrijenhoek, 1988; Brewsteret al., 2016; Goldmanet al.,2016;Whitehouseet al.,2017)。
东北马鹿(Cervus elaphus xanthopygus)主要分布于东北大、小兴安岭及长白山等林区,是高格斯台罕乌拉国家级自然保护区的主要保护对象,同时也是内蒙古森林—草原交错带中的关键物种。内蒙古高格斯台罕乌拉国家级自然保护区为典型的蒙古栎(Quercus mongolica) 林、山杨(Populus davidiana)林等森林植被类型与大针茅(Stipa grandis)草原、贝加尔针茅(Stipa baicalensis)草原等草原植被类型共存的森林—草原交错带(邓芳,2005)。生态交错带(Ecotone)即相邻生态系统的过渡带,其特征由相邻生态系统之间相互作用的空间、时间和强度所决定(Holland,1988)。通常,在生态交错带中植物多样性越高,群落结构更复杂,这为动物提供了更多的隐蔽和摄食条件(王庆锁等,1997)。生态交错带对干扰或环境变化具有较强的敏感性,天气、气候、降雨量和自然大火等非生物因素以及食草动物的选择性采食行为等会对生态交错带中的植物群落产生影响。同时,植物群落结构的改变也会影响食草动物的食物组成、分布、种群数量、行为乃至营养策略(Dublinet al., 1990; Olofssonet al., 2004; De Micheleet al.,2011; Dioufet al., 2012; Mooney and Singer, 2012)。北方冬季寒冷低温,积雪较深,以及食物的营养质量和可利用性降低,致使东北马鹿遭受严重的营养摄取胁迫。森林—草原交错带中植物的群落组成不同于北方典型的针阔混交林,木本植物占比较低(黄晨,2015),而野生东北马鹿冬季主要以木本植物的嫩枝为食(黄晨等,2014;钟林强,2020)。因此,为保证冬季营养摄入均衡,分布于森林—草原交错带内的野生东北马鹿种群可能呈现特定的食物组成模式。
野生有蹄类为满足营养需求在不同的时间段和环境中对食物进行选择,且这种选择逐渐积累形成相对稳定的食物组成模式,因此,对有蹄类的食物组成模式进行分析是研究和监测有蹄类种群营养状态的基础和重要组成部分。而目前,森林—草原交错带中东北马鹿的食物组成研究大多局限于冬季和种群整体,并没有对不同年份之间以及东北马鹿种群内冬季食物组成的相似性和差异性进行比较与讨论(何欢,2015;邢博,2016;孙悦,2018)。本研究于2019年和2020年,分析内蒙古森林—草原交错带中东北马鹿个体间冬季食物组成的相似性,探索东北马鹿种群呈现何种食物组成模式及种群内食物组成模式是否存在差异,为森林—草原交错带中东北马鹿营养生态学的进一步研究与保护提供基础数据。
1 研究方法
1.1 研究地区概况
内蒙古高格斯台罕乌拉国家级自然保护区位于内蒙古赤峰市境内(北纬44°41′03″~45°08′44″,东经119°03′30″~119°39′08″),以罕山林场为主体,海拔900 ~1 500 m,总面积106 284 hm2。该保护区与大兴安岭北部山地相连,西南部与燕山北部山地相邻,是东亚阔叶林向西伯利亚寒温带针叶林、草原向森林双重交错的过渡地带。保护区内植物区系复杂,呈现典型的森林—草原交错带结构,山体背阴处以白桦(Betula platyphylla)林和山杨林为主要优势植被,其林下灌木以兴安杜鹃(Rhododendron dauricum)、绣 线菊(Spiraea salicifolia) 和胡枝子(Lespedeza bicolor)为主;山体阳坡和半阳坡以蒙古栎林和家榆(Ulmus pumila) 林为主,林下灌木以山杏(Armeniaca sibirica)、绣线菊和虎榛子(Ostryopsis davidiana) 等为主;山体顶部向阳处则以草原为主(邓芳,2005)(图1)。保护区的动物区系,呈现出华北区、东北区和蒙新区3 个区域之间相互渗透的状态,主要野生动物有东北马鹿、狍(Capreolus pygargus)、野猪(Sus scrofa)、猞猁(Lynx lynx)、狼(Canis lupus)、黑鹳(Ciconia nigra)和大鸨(Otis tarda)等(郭金昊,2020)。

图1 研究区域地理位置及植被分布Fig.1 The study area and vegetation distribution
1.2 粪便样品采集
2019年3月和2020年1月,在内蒙古高格斯台罕乌拉国家级自然保护区内,分别进行为期10 d的样品采集。根据保护区内东北马鹿的分布状况,设计不同生境类型的采样区域,由于高格斯台地区东北马鹿冬季的家域面积为55.87 ~114.58 hm2(杨淼,2018),因此不同采样区域之间间隔2 km以上。在一个采样区域内的随机样线上发现东北马鹿的新鲜足迹链时,通过反向跟踪足迹链,使用PE 手套采集新鲜的粪便样品置于封口袋中,采集粪样后,再寻找新的足迹链跟踪,以保证在这个区域内所采集的粪样代表一个个体,寻找及跟踪足迹链时每隔100 ~150 m 测定雪深(朱明月等,2019)。
1.3 植物样品采集及鉴定
寻找及跟踪东北马鹿足迹链时,将所有含啃食痕迹的植物进行采集,采集时保留该植物的花、茎、叶等具有辨识特征的器官,并详细记录采集日期、地点和生境等信息。如果采样时部分植物的花、果实、叶枯萎凋落,则夏季在原采集地对同种植物进行补采。每种植物样本分成两份,一份装入牛皮纸档案袋中,用于粪便显微分析中植物细胞壁的对照;另一份用标本夹收集,先依据《内蒙古植物志》(赵一之等,2020)中的植物分类检索表逐次逐项检索,对植物物种进行初步的判定,再与保护区的植物标本馆核对标本进行检验。若结果一致,即确定其物种名,不一致,则请保护区的专业人员鉴定其物种名。
1.4 显微装片制备及拍摄
将采集到的植物样品置于105℃烘箱中烘48 h至恒重,粉碎机粉碎后,使用80 ~100目的分样筛分筛。取0.5 g 粉末样品于培养皿中,加入浓度10%的次氯酸钠溶液40 mL进行消化。夹取消化良好的样品于载玻片中,蒸馏水清洗3遍,使用甘油封片,加拿大树脂封边后置于10×40 显微镜下观察,选取细胞结构清晰,无杂质的视野进行拍照(朱明月等,2019)。每种植物样本制片10张,拍摄50 张照片,构建植物细胞结构形态图谱,用于粪便显微装片镜检时鉴定植物组织碎片。
将采集到的粪便样品置于105℃烘箱中烘48 h至恒重,用研钵研磨后以上述相同方法制备显微装片。每个粪便样本制片15 张。将制作好的粪便显微装片置于10 × 40 的电子照相显微镜下镜检,每张装片按由左到右、由下到上的方向选取20 个视野拍照,拍摄时两个视野间留出1 个视野不拍摄,1个植物碎片记为1次。
1.5 统计分析
通过频率转换法F= 100(1 -e-Di)(Johnson,1982),计算每个粪堆中每种植物可辨认碎片的平均密度(D),每种植物的相对密度(relative density,RD)通过公式:RD=Di∑Di计算得出,并以RD作为野生东北马鹿食物组成的标准。使用Shapiro-Wilk 检验数据是否符合正态分布。采用Mann-WhitneyUtest比较两年冬季中各食物组成RD值之间的差异,显著性水平设置为α=0.05。
通过Shannon-Weiner 多样性指数(H)、Pielou均匀度指数(J)和营养生态位指数(B)分析冬季东北马鹿食物组成的多样性(吕忠海等,2019)。

式中,Pi为植物i碎片与总碎片的比值,HMAX=lnS,S为植物种数。以东北马鹿2019 年和2020 年粪便显微分析中各植物的多度为原始数据(行代表粪便样本,列代表植物种类),由于原始数据为多度数据,且量纲一致,同时为避免排序时可能出现的弓形效应,使用去趋势分析(Detrended correspondence analysis,DCA)对数据进行排序,以分析东北马鹿冬季食物组成的相似性模式(Graham and Vrijenhoek, 1988; Loureiro-Crippa and Hahn, 2006;张玮琪等,2019)。所有统计在R 软件(4.1.0 版本) 中实现。
2 结果
2.1 东北马鹿冬季食物组成
2019 年3 月,平均雪深3.2 cm,共采集新鲜粪便33 堆,辨认植物组织碎片11 205 个。共采食21 种(科) 植物,RD 值大于1%的食物有9 种,其中 禾 本 科 (Poaceae spp.) (26.44%)、 蒙 古 栎(20.63%)、山杏(15.23%)、家榆(14.48%)和细叶沼柳(Salix rosmarinifolia)(8.22%)等为东北马鹿在该时间段内的主要食物,共占85.00% (图2)。在取食植物类别上以木本植物为主,为67.16%;草本植物共占30.70%,其中禾本科占26.44%,非禾本科占4.26%。
2020 年1 月,平均雪深6.7 cm,共采集新鲜粪便65 堆,辨认植物组织碎片20 197 个。共采食22 种(科)植物,RD 值大于1%的食物有12 种,其中蒙古栎(23.94%)、细叶沼柳(16.77%)、山杏(15.13%)、禾本科(10.86%)、云杉(Picea asperata) (9.88%) 和家榆(7.07%) 等为东北马鹿在该时间段内的主要食物,共占83.66% (图2)。在取食植物类别上,木本植物较2019 年显著提高,占比达83.76%;而草本植物取食较少,共占15.62%,其中禾本科占10.86%,非禾本科占4.76%。
2019 年和2020 年冬季东北马鹿采食的食物种类相似,均以蒙古栎、山杏、细叶沼柳、家榆以及禾本科等植物为主要食物,但两年冬季中云杉、家榆、白桦等部分食物的RD 值存在显著差异(图2)。

图2 森林—草原交错带东北马鹿冬季食物组成及差异显著分析.*P <0.05,**P <0.01,***P <0.001Fig. 2 Difference of food composition of red deer in forest-grassland ecotone in winter.*P <0.05,**P <0.01,***P <0.001
2020 年冬季东北马鹿的可食植物种数、多样性指数、均匀度指数以及营养生态位指数均大于2019年冬季(表1)。

表1 森林—草原交错带东北马鹿冬季食物多样性、均匀度及营养生态位指数Table 1 Food diversity,evenness,and torphic niche index of red deer in forest-grassland ecotone in winter
2.2 东北马鹿冬季食物组成去趋势分析
将两年冬季中98 堆东北马鹿粪样的食物组成按食物种类通过去趋势分析进行排序并绘制了95%的置信椭圆区间,4 个轴的特征值分别是0.465 0、0.198 4、0.125 7 及0.102 5,轴1 和轴2能够解释数据74.41%的变化(表2,图3)。2019年和2020 年的样本在第二象限以及第三象限中有较大重叠,即两年冬季中东北马鹿大部分个体呈现出以蒙古栎、山杏、细叶沼柳、家榆和禾本科为主要食物的食物组成模式,而2020年部分个体在1轴具有较高评分,且远离大部分个体,表明这些个体具有不同的以云杉为主的食物组成模式(图3a)。铁杆蒿(Artemisia gmelinii)、卫矛(Euonymus alatus)、山刺玫(Rosa davurica) 和益母草(Leonurusjaponicus)等RD 值小于1%的偶食食物在排序图中均离原点较远,表明偶食食物具有较高信息荷载量,即东北马鹿个体间偶食食物所占的比例存在较大变化(图3b)。而蒙古栎、山杏等虽为东北马鹿的主要食物,但离原点相对较近,信息荷载量较低,表明冬季东北马鹿种群对蒙古栎、山杏等食物采食的比例较为稳定。毛莲蒿(Artemisia vestita)、落叶松(Larix gmelinii)和益母草在1 轴和2 轴上的评分相似,表明毛莲蒿、落叶松和益母草倾向于同时出现在东北马鹿食物中,与之类似,艾蒿(Artemisia lavandulifolia)和山杏也倾向于同时出现在东北马鹿食物中。

图3 东北马鹿冬季食物组成按食物种类DCA排序. a:粪便样本;b:食物物种Fig. 3 DCA of the food composition of red deer by food species in winter. A:fecal samples;b:food species
按东北马鹿食物类别进行去趋势分析结果显示,4 个轴的特征值分别为0.551 2、0.149 8、0.072 3 及0.247 9,轴1 和轴2 能够解释数据68.65%的变化(表3,图4)。1 轴能够区分针叶乔木和落叶乔木,2 轴能够区分禾本科草本与其他植物类别。落叶乔木和灌木倾向于同时出现在东北马鹿食物中,而禾本科草本、非禾本科草本及针叶乔木都不倾向于与其他植物类别同时出现。落叶乔木和灌木在1 轴和2 轴上的评分均较低,离原点较近,暗示这些植物是东北马鹿的主要食物但不是导致食物组成变化的主要因素。针叶乔木和禾本科草本分别在1 轴及2 轴上有较高的评分,且离原点较远,而针叶乔木在1轴上具有较高评分是由于2020 年部分个体大量取食云杉所致。因此禾本科和云杉是引起东北马鹿冬季食物组成变化的主因。
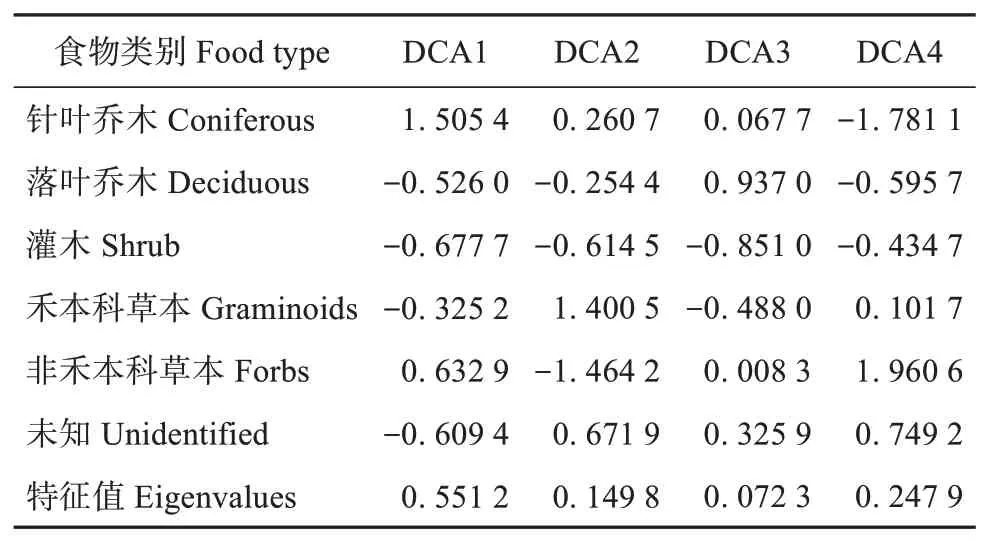
表3 以食物类别分类进行DCA的物种得分表和特征根Table 3 Table of species scores and eigenvalues for DCA by food type classification

图4 东北马鹿冬季食物组成按食物类别DCA排序. a:粪便样本;b:食物类别Fig. 4 DCA of the food composition of red deer by food type in winter. a:fecal samples;b:food type
3 讨论
3.1 东北马鹿冬季食物组成
2019 年和2020 年冬季,东北马鹿取食的植物物种基本稳定,均以蒙古栎、山杏、细叶沼柳、家榆和禾本科草本为主要食物,这与此前内蒙古森林—草原交错带中东北马鹿冬季的食性研究结果相似(黄晨,2015;何欢,2015;邢博,2016;孙悦,2018),但较高比例采食云杉(9.88%) 这一现象并未在此前研究中发现。云杉属针叶树种,含有较高的次生代谢产物并适口性较差,有蹄类通常会避免过多取食此类植物(Verheyden-Tixieret al., 2008;Azoritet al., 2012)。而本研究中东北马鹿大量采食云杉的现象可能与有蹄类在食物短缺时会以针叶树种作为补充食物的采食策略有关(Cręte and Courtois, 1997; Stormset al., 2008)。2020 年冬季,内蒙古森林—草原交错带中雪被较厚,许多低矮的草本植物及落叶等食物被积雪覆盖,且交错带中木本植物占比较低,东北马鹿食物的可获得性降低,而云杉含有较高的蛋白质、脂肪和碳水化合物等营养物质(钟林强,2020),因此东北马鹿可能通过采食较高比例的云杉来弥补主要食物不足(窦红亮,2013;张玮琪,2013)。
两年冬季的东北马鹿食物组成中,部分食物的RD 值差异显著,2020年冬季东北马鹿显著取食更多比例的细叶沼柳、白桦以及绣线菊;而2019年冬季则显著取食更多比例的家榆、卫矛、禾本科以及叉分蓼(Polygonum divaricatum)。造成这种差异的原因可能是环境,与2020 年相比,2019 年冬季雪被较浅,气温较高,低矮的草本植物及家榆落叶等食物裸露在地表未被积雪覆盖,可获得性高。同时,2020 年冬季东北马鹿的食物多样性指数、均匀度指数以及营养生态位指数均较大,与Dumont 等(2005) 和Seto 等(2015) 的研究结果相似,这也验证了有蹄类在食物资源较为丰富时,只取食那些优质食物;而在食物资源匮乏时,增加食物种类来弥补主要食物摄入不足(Nudds,1980)。虽然上述食物的RD 值差异显著,但家榆、细叶沼柳和禾本科在两年冬季中均作为东北马鹿主要食物出现,而白桦、卫矛、绣线菊和叉分蓼也均为偶食食物,表明森林—草原交错带中东北马鹿冬季的食物组成较为稳定。
3.2 冬季东北马鹿食物组成模式
结合东北马鹿冬季食物种类排序图、食物类别排序图以及食物组成数据,表明在2019 年和2020 年冬季,大多数东北马鹿的主要食物组成是蒙古栎、山杏、细叶沼柳、家榆等落叶乔木和灌木,同时采食较高比例禾本科草本植物的模式。这种食物组成模式与主要由山地森林植被类型、灌丛植被类型和草原植被类型组成的内蒙古森林—草原交错带的生境结构相符合(邓芳,2005)。但与黑龙江典型森林生态系统中东北马鹿的食物组成模式存在较大差异,在典型森林生态系统的东北马鹿冬季食物中木本植物占90%以上,较少甚至不取食禾本科草本植物(李言阔和张明海,2005;张玮琪,2013;黄晨等,2014;黄晨,2015;冯源,2018;朱明月等,2019)。这说明当植被类型的空间格局发生变化时,有蹄类的食物选择将会发生相应的改变(李俊生等,2003),进而导致食物组成模式也发生变化(Hu,2002)。
研究结果显示,2020 年冬季部分东北马鹿呈现出以云杉为主要食物(RD 值>60%)的食物组成模式。这种模式与东北地区的驼鹿(Alces alces)在营养摄取胁迫期间通过采食大量云杉来弥补食物不足的采食策略相同(张玮琪,2013;窦红亮,2013),且本研究中大量采食云杉的样本数量占2020 年冬季样本量的12.3%,占两年冬季总样本量的8.2%。可见,部分东北马鹿个体大量采食云杉并不是偶然行为,而是应对冬季食物短缺时,所呈现出的特殊食物组成模式。
