低氧对非小细胞肺癌细胞PDHA1表达及生物学行为的影响
2022-06-05曾宪旭雷冬梅班振英
曹 静,曾宪旭,雷冬梅,班振英,张 威
郑州大学第三附属医院病理科 郑州 450052
肺鳞状细胞癌(简称肺鳞癌)及腺癌约占非小细胞肺癌总数的90%以上,是其主要的组织学类型,严重危害人类健康[1]。已知三羧酸循环氧化磷酸化及糖酵解是人体细胞中的主要产能方式;线粒体中三羧酸循环正常进行的前提为丙酮酸脱氢酶E1α亚单位(pyruvate dehydrogenase E1α subunit,PDHA1)正常表达,PDHA1表达下降能使氧化磷酸化受阻,代谢途径转为糖酵解;大多数肿瘤依赖糖酵解代谢,被称为Warburg效应[2]。此外,PDHA1表达下调在致癌信号整合、代谢重编程、促进细胞增殖及抑制凋亡中也发挥作用[3]。肿瘤组织一般处于低氧的微环境。研究[4-5]表明低氧导致肿瘤细胞“未分化”,甚至是“去分化”而获得干性,促进肿瘤的生长、浸润、转移和复发。本研究旨在探索低氧对非小细胞肺癌细胞中PDHA1表达的影响,及其与恶性生物学行为之间的关系。
1 材料与方法
1.1 材料肺鳞癌细胞株LC-42、肺腺癌细胞株SELS均为挪威奥斯陆大学肿瘤研究实验室冻存株。Xvivo低氧细胞工作站(美国BioSpherix公司),超净工作台(美国AirClean公司),CO2细胞培养箱(美国Thermo Fisher公司),Transwell小室(美国Corning公司),PDHA1(C54G1)一抗(Cell Signaling Technology公司)。
1.2 细胞培养条件常氧:细胞在体积分数20%O2、5%CO2、75%N2,37 ℃及饱和湿度下培养。低氧:细胞常氧培养24 h,待细胞贴壁转移至Xvivo低氧细胞工作站,在体积分数1%O2、5%CO2、94%N2,37 ℃及饱和湿度下培养。
1.3 细胞形态学观察、生长曲线绘制及倍增时间测定LC-42、SELS细胞分别接种于6孔板中,调整细胞密度为1.5×104个/mL,每孔约接种3 mL,分为2组,分别于低氧及常氧条件下培养,每组6孔。连续培养6 d,观察细胞形态学变化。每天消化1孔,细胞计数仪计算细胞数量,绘制生长曲线。按Patterson公式[6]计算细胞倍增时间。实验重复4次。
1.4 细胞侵袭能力检测LC-42、SELS细胞分为2组,分别于低氧及常氧条件下连续培养72 h后,制备细胞悬液,调整细胞密度为5×105个/mL。Matrigel基质胶稀释后铺满Transwell小室,上室加入100 μL细胞悬液,下室加入500 μL含体积分数20%胎牛血清的培养液,常氧环境下培养48 h。取出培养板,PBS洗2遍,以体积分数5%戊二醛固定,20 g/L结晶紫染色,PBS洗2遍,低倍镜(×100)下选取10个视野,Image J软件计数穿膜细胞,取总数。实验重复6次。
1.5 细胞克隆形成能力检测LC-42、SELS细胞分为2组,分别于低氧及常氧条件下连续培养72 h,消化细胞,加入培养液重悬细胞。取细胞悬液10 μL并加入等体积台盼蓝染液1~2 min后将细胞接种于60 mm培养皿,每个培养皿接种约1 000个细胞。常氧培养2周后40 g/L多聚甲醛固定,20 g/L结晶紫染色。记录克隆数,克隆形成率=克隆数/接种细胞数×100%。实验重复6次。
1.6 细胞中PDHA1蛋白表达水平检测LC-42、SELS细胞分别于低氧及常氧条件下连续培养72 h,胰蛋白酶消化收集细胞,加入150 μL RIPA裂解液冰上裂解30 min,4 ℃15 000 r/min离心5 min后抽提上清,Bradford法测定蛋白浓度。加入上样缓冲液,沸水加热5 min。聚丙烯酰胺凝胶电泳、转膜,用含50 g/L脱脂牛奶的TBST封闭1 h。加PDHA1抗体 (按1∶1 000稀释),4 ℃过夜孵育,加二抗,室温孵育2 h,洗膜并曝光。使用Image J软件进行灰度值分析,以目的条带与内参(GAPDH)条带灰度值的比值作为目的蛋白的相对表达量。实验重复4次。
1.7 统计学处理应用SPSS 19.0处理数据。采用两独立样本t检验比较低氧组及常氧组细胞倍增时间、穿膜细胞数、克隆形成率、PDHA1蛋白表达的差异,采用Pearson相关分析低氧条件下两种细胞株PDHA1蛋白表达与穿膜细胞数及克隆形成率的相关性。检验水准α=0.05。
2 结果
2.1 各组细胞形态及生长情况培养第6天,镜下(图1)可见常氧组LC-42、SELS细胞为多角形、梭形、椭圆形,低氧组LC-42、SELS细胞间隙增大,细胞膜模糊。生长曲线(图2)表明低氧组细胞生长缓慢,细胞倍增时间延长(表1)。
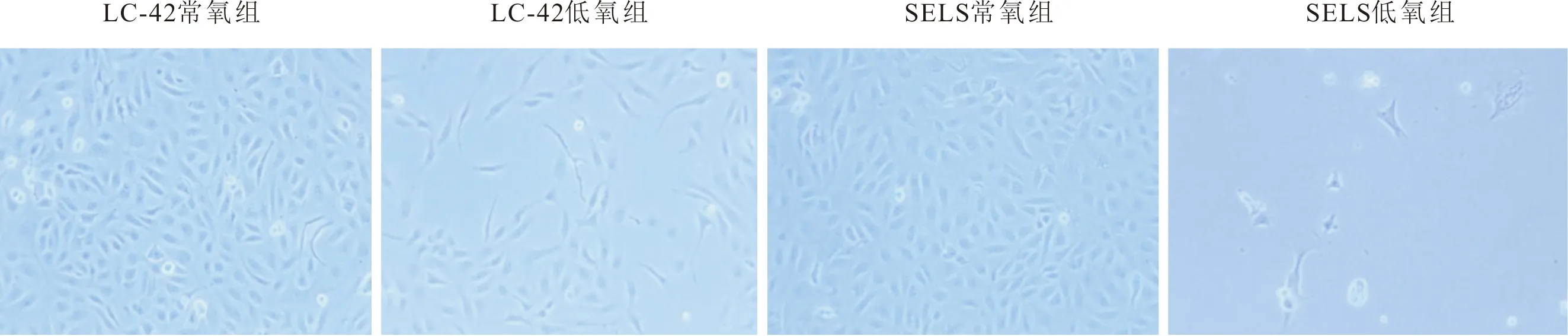
图1 肺癌细胞株低氧组及常氧组细胞形态(×400)

图2 LC-42(左)、SELS(右)细胞常氧组(a)与低氧组(b)的生长曲线

表1 各组细胞倍增时间比较 h
2.2 各组细胞侵袭能力及克隆形成能力比较与常氧组相比,两种细胞低氧组的穿膜细胞数及克隆形成率均增加,见图3、表2。
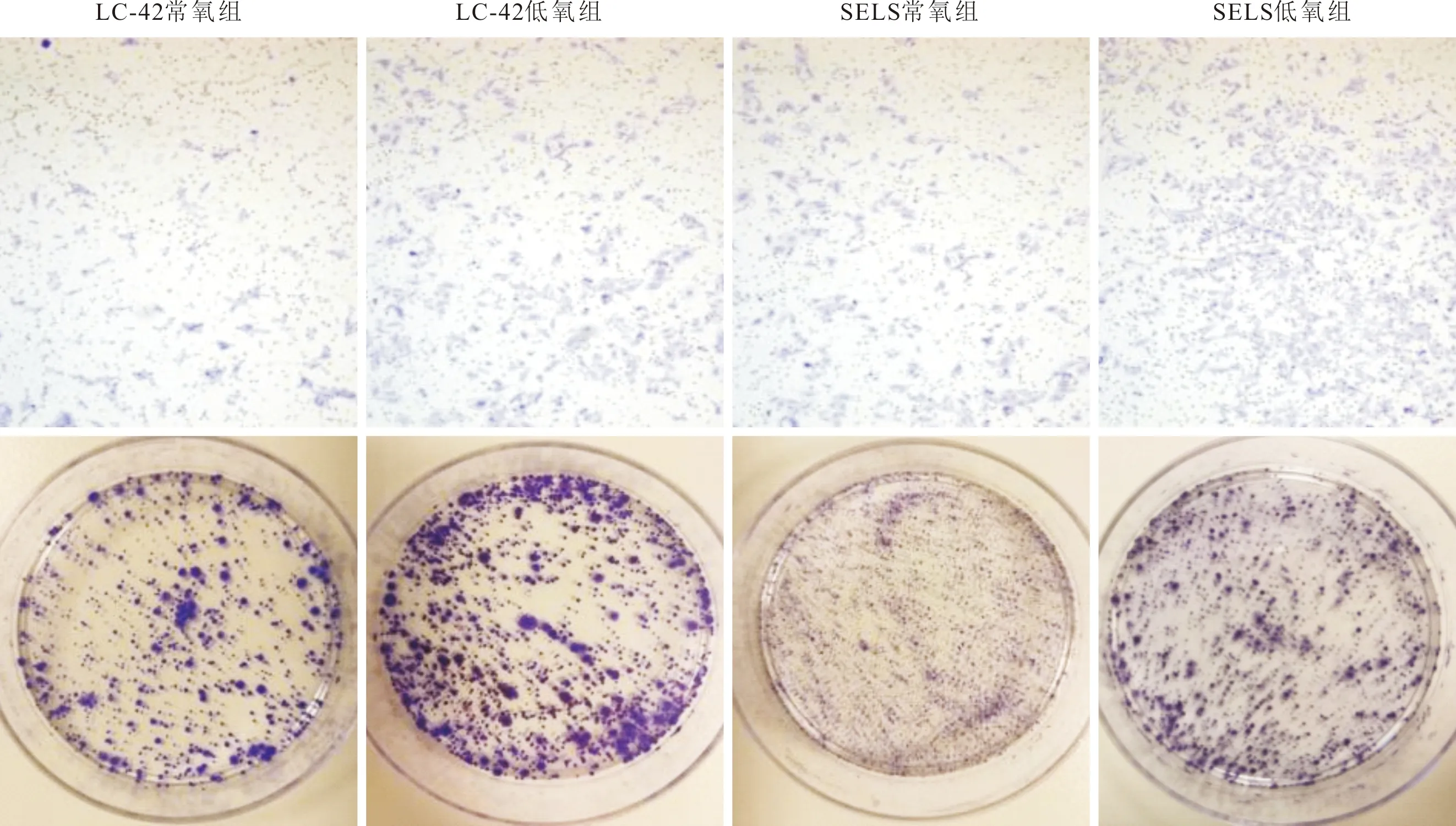
上排:侵袭实验结果;下排:克隆形成实验结果

2.3 各组细胞PDHA1蛋白表达的比较与常氧组相比,两种细胞低氧组PDHA1蛋白的表达均降低(P<0.05)(图4、表3)。
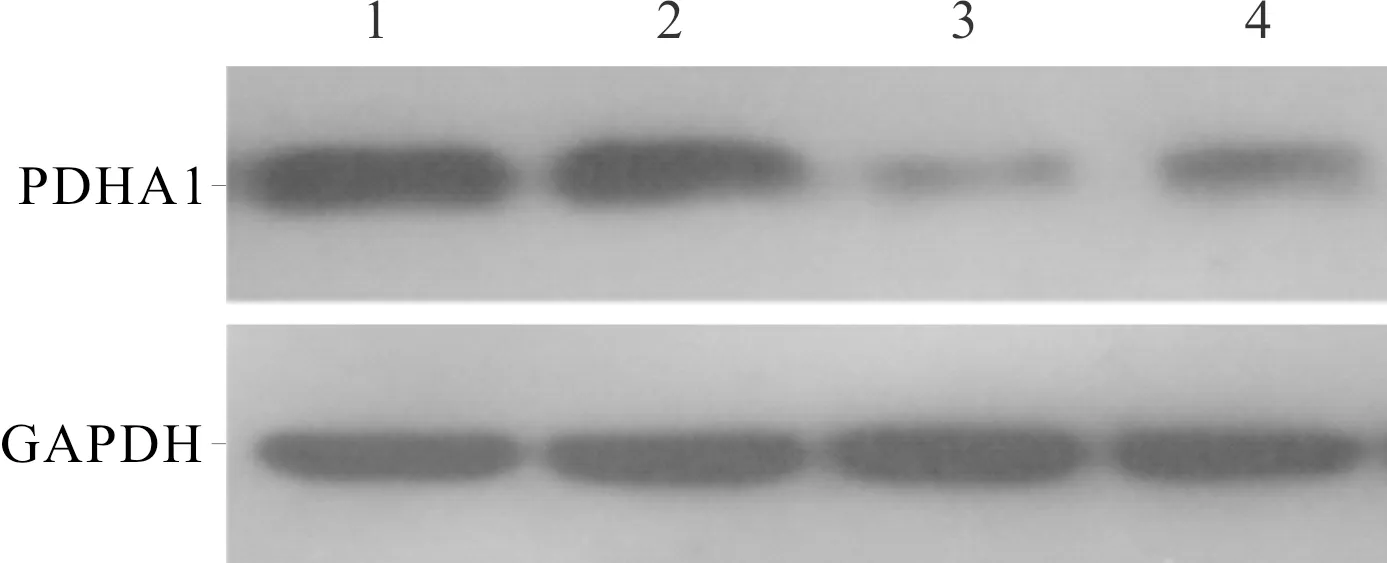
1~4:分别为LC-42常氧组、SELS常氧组、LC-42低氧组、SELS低氧组

表3 各组细胞PDHA1蛋白表达的比较
2.4 低氧组细胞PDHA1蛋白表达与穿膜细胞数及克隆形成率的相关性Pearson相关分析表明,两种细胞低氧组中PDHA1蛋白表达与穿膜细胞数及克隆形成率均呈负相关(表4)。

表4 低氧组PDHA1蛋白表达与穿膜细胞数及克隆形成率的相关性 r(P)
3 讨论
本研究对比分析了常氧及低氧培养条件下两种非小细胞肺癌细胞株生物学行为及PDHA1表达的差异,结果表明低氧会导致这两种细胞增殖变慢,侵袭能力及克隆形成能力增强。分析机制可能为低氧刺激细胞干性增强,且调动了其中部分侧种群细胞[7]。有研究[8]表明实体肿瘤的低氧区普遍存在侧种群细胞。相关研究[9]发现,荷瘤鼠瘤组织低氧区存在侧种群细胞集中性分布特征,由此推测低氧促进侧种群细胞形成肿瘤。此外,干性基因Oct-4等表达增强亦证明肿瘤细胞的干性增强[10]。
低氧可以诱导很多基因产物如干细胞标记物(Oct-3/4、Nanog等),糖酵解及血管源性标记物等的表达[10]。本研究显示低氧处理后肺鳞癌及腺癌细胞中PDHA1表达下降,且PDHA1表达与穿膜细胞数及克隆形成率呈负相关。PDHA1在糖代谢中的重要功能是连接糖酵解和线粒体三羧酸循环,PDHA1表达下降可使氧化磷酸化受阻,糖酵解增加。此外,PDHA1下调在致癌信号整合、代谢重编程、促进细胞增殖及抑制凋亡中也发挥作用。一些学者[11-14]发现PDHA1蛋白表达减少与肝细胞癌、前列腺癌、食管鳞癌预后差有关,PDHA1敲除的前列腺癌细胞增殖、迁移能力增强。本研究结果与上述研究一致。
总之,低氧导致非小细胞肺癌细胞中PDHA1表达下降及恶性生物学行为增强的结果有助于理解PDHA1在Warburg效应中的重要功能。在抗肿瘤研究领域,肿瘤的PDHA1表达下降导致的糖酵解增强已经引起重视。有研究[15]表明二氯乙酸盐可通过激活PDHA1,使糖代谢转向氧化磷酸化,抑制肿瘤的增殖。随着此方面机制的进一步明确,未来将能研发出更多的PDHA1靶点药物用于非小细胞肺癌的治疗。
