高原鼠兔干扰对高寒草地碳循环的影响研究进展
2022-06-02邵梓桐
邵梓桐,秦 彧
(1.西北农林科技大学,草业与草原学院,陕西 杨凌 712100; 2. 中国科学院西北生态环境资源研究院,冰冻圈科学国家重点实验室,甘肃 兰州 730000)
全球变化及扰动对陆地生态系统碳循环的影响及其反馈作用是全球变化研究等相关学科的重点科学问题,亦是各国学术界和政府关注的热点问题。陆地生态系统作为全球碳循环的重要组成部分,在全球碳收支平衡中占有主导地位[1]。高寒草地作为全球陆地生态系统重要的碳库之一[2-4],土壤有机碳储量约为全球碳库的2.5%[5],在未来全球碳循环调控中发挥着重要的作用[6]。准确估算青藏高原高寒草地生态系统碳储量和碳通量是评估该区碳循环对全球气候变化反馈效应的基础[7-8],其研究成果可为全球碳平衡研究提供科学依据,提高全球碳源汇评估的精度[9]。
在过去的几十年,约有1/3的高寒草地发生了不同程度的退化[10],导致大约42%±2%的土壤有机碳损失[11]。除了全球气候变化和人类活动的干扰[12],栖息在高寒草地的掘土动物对生态系统碳平衡的扰动也在持续增加,影响了整个生态系统碳循环过程和时空分布格局[13]。高原鼠兔(Ochotonacurzoniae)是高寒草地生态系统中的关键种[14-15],在高寒草地生态系统物质循环和能量流动中具有重要作用[16-17]。高原鼠兔扰动导致地表形成新鼠丘、旧鼠丘、植被和裸地等不同斑块镶嵌共存的景观格局[18]。这种异质性的下垫面改变了土壤的水热属性[19],加剧了高寒草地植被生产力和土壤有机碳的空间异质性分布[20],改变了生态系统碳排放[21-22]和碳交换过程[23-34]。
然而,目前关于高原鼠兔干扰对陆地生态系统碳循环关系的研究还很薄弱且相对滞后,尤其是在定量评估方面还存在很大的不确定性[25],成为制约高寒草地生态系统碳收支以及碳源汇功能准确评估的掣肘。为此,笔者试图从高原鼠兔干扰影响高寒草地的主要途径,高原鼠兔干扰对生态系统有机碳储量及其生态系统碳排放的影响过程和作用机制等方面,综述高原鼠兔干扰对高寒草地碳循环影响的研究进展,以期为高寒草地碳源汇准确评估和碳汇管理对策提供理论依据和科学参考。同时,明晰高原鼠兔在高寒草地生态系统碳循环中的作用,有望在小型啮齿动物与生态系统功能维持之间的关系方面取得新的认识和进展,对于制定合理的高寒草地生态系统管理措施和生物多样性保护对策具有重要的理论和现实意义。
1 高原鼠兔生物学特性
高原鼠兔为兔形目鼠兔科鼠兔属动物,成年鼠兔体长约为10~17 cm,平均体重约为150~200 g,耳大、额隆、鼻尖、上下唇为黑色,因此又名黑唇鼠兔[26-27]。化石证据显示,高原鼠兔由距今约3 700万年的古鼠兔亚科(SubfamilySinolagomyinae)演化而来,是一种经过长期的自然选择形成适应高寒缺氧环境而保留下来的青藏高原特有的小型植食性啮齿类动物[28]。高原鼠兔广泛分布于青藏高原且一般营家族式生活[14,29],种群密度可以达到每公顷300~350只[30]。高原鼠兔偏向于选择植被高度较低,视野开阔且土壤疏松的环境作为栖息地[31],这种栖息地有利于高原鼠兔减少能量支出,扩展其生存空间和躲避天敌[32]。尽管研究指出高原鼠兔广泛分布在海拔3 100~5 100 m的高寒草甸、高寒草原、草甸草原及高寒荒漠草原地带[33],但是在高寒草甸高原鼠兔种群密度显著高于其他草地类型[34-35]。
高原鼠兔繁殖具有明显的季节性,主要集中在每年4—8月,性成熟的雌鼠每年繁殖1~2胎[36]。为了维持种群数量的稳定,当青藏高原恶劣天气致使高原鼠兔第一胎繁殖失败时,其第二胎会持续增加可食性资源以及空间资源来改变自身体重增长模式[37]。高原鼠兔每胎产仔数为3~7只[38],繁殖周期约为3周,雌鼠分娩结束后即进入下一个繁殖周期[39-40]。高原鼠兔在冷季的死亡率要高于暖季,冷季和暖季平均死亡率分别为0.446和0.283[41]。一般认为食物短缺和低温是导致高原鼠兔冷季死亡的主要原因[42],而暖季大量的降水是影响高原鼠兔存活主要原因[40]。高原鼠兔最大寿命为2~3年,显著低于其他鼠兔,例如Ochotonapricei和Ochotonaprinceps的最大寿命分别为4年和6~7年[43]。
2 高原鼠兔影响高寒草地的主要方式
高原鼠兔长期生活在景观开阔的草地上,隐蔽条件差,加上严酷的高寒气候,高原鼠兔形成了在地下挖掘洞道,洞穴栖息的生活方式[44-45]。高原鼠兔挖掘洞道时,一方面在地表形成与地下洞道系统连通的洞口[46],另一方面将草皮层下的土壤堆至洞外形成裸露疏松的鼠丘[21,47]。高原鼠兔洞道系统和鼠丘改变了这种区域内的水热过程,养分的垂直分布格局和植被物种组成[48-50],客观上构造了有别于周围植被斑块的微生境。尽管营洞穴生活,但是成年鼠兔觅食的时间大约占其在地面活动总时间的74%[51]。此外,高原鼠兔通常会刈割洞穴周围的高大植物[52-53],这不仅易于其发现和躲避捕食者,同时也释放了低矮植被的生长潜力[54]。同时高原鼠兔具有选择性采食行为且其食物生态位与家畜高度重叠[55]。据估算,70只成年鼠兔年采食牧草量相当于4.8只藏绵羊一年的饲草量[56]。由此可见,高原鼠兔是除家畜之外影响高寒草地生态系统最重要的生物因子之一,采食、堆土和挖掘是其影响高寒草地最主要的方式(图1)。

图1 高原鼠兔干扰对高寒草地生态系统功能影响的方式Fig.1 Influence ways of pikas disturbance on ecosystem function of alpine grassland
除了对高寒草地的直接影响外,高原鼠兔堆土形成的鼠丘和挖掘形成的洞道侵蚀和坍塌后会形成秃斑(图2),因而高原鼠兔扰动被认为是高寒草地秃斑化的驱动力之一[57-58]。秃斑由于缺少植被覆盖,易受侵蚀的影响而引起土壤和养分的流失[59-60]。同时斑块的进一步扩张导致生产力下降[61],植被恢复的过程就会变得异常缓慢[47]。秃斑化下高寒草地植被生长除了受土壤养分、土壤水分和热量的影响外[62],秃斑的热岛效应增加了土壤的容积热容量,导热率和热扩散率[63-64],在近地表形成持续加热状态。秃斑周边植被为了维持其适宜的生长温度,加快了土壤水分蒸散[65],加剧了土壤养分和生产力的异质性分布[20,66]。由此可见,高原鼠兔还可以通过与其扰动相关联的秃斑化这种间接的方式对高寒草地生态系统功能产生影响(图2)。

图2 高原鼠兔扰动下秃斑的形成机制及其对高寒草地的影响Fig.2 The formation and development of patchiness by pikas’ disturbance and its cascade effect on alpine grassland
3 高原鼠兔干扰对高寒草地生态系统有机碳储量的影响
3.1 高原鼠兔干扰对高寒草地生态系统有机碳储量的积极影响
青藏高原高寒区由于缓慢的成土作用,土壤层普遍发育较薄[67],而低温下缓慢的分解速率导致高寒草地生态系统碳主要累积在表层土壤[68]。高原鼠兔作为高寒区生态系统中的一类活跃组分[50,69],其构筑的地下洞道主要分布于距地表30~50 cm的土壤层[45,70]。由此可见,高原鼠兔挖掘能力有限,其扰动对高寒草地土壤有机碳的影响主要集中在表层。高原鼠兔通过挖掘及排泄粪尿等活动,在改变高寒草地的地表生态水文过程的同时[19],也促进了生态系统的物质循环[16]。微生物是影响陆地生态系统碳循环的重要生物因子,其不同的代谢途径和生物周转过程影响了碳的生物地球化学循环过程[71]。土壤微生物群落结构和组成变化主要受土壤和植被特性的影响[72],而微生物群落对环境因子的响应甚至会对全球范围碳循环和温室气体的排放产生影响[71]。高原鼠兔的挖掘活动疏松和改善了土壤条件,增加深层土壤的氧气含量和透气性,改变了土壤微生物群落结构和活性,进而促进有机质的分解和碳素的矿化[73-74]。此外,高原鼠兔会为冬季贮藏干草以减少出外觅食被捕食的风险[75],而且伴随觅食及领域等活动,常有选择地在其巢区内排泄粪、尿[16]。贮藏的剩余干草碎屑以及埋在土层中的粪、尿不仅为土壤提供了大量有机质[39,76],同时也为微生物提供了更多的呼吸底物,增加了高寒草地生态系统的异养呼吸(图3)。值得注意的是,高原鼠兔干扰对高寒草地土壤有机碳储量的积极影响依赖于其干扰强度,适量的干扰增加了土壤粉粒比例,提高了土壤微生物对动植物残体分解速率,导致土壤有机碳储量增加,而高强度扰动下土壤微生物对动植物残体的分解速率下降,整体上降低了土壤有机碳储量[77-78]。
3.2 高原鼠兔干扰对高寒草地生态系统有机碳储量的消极影响
然而除了对高寒草地生态系统有机碳累积的积极作用外,高原鼠兔干扰也会引起潜在的生态系统有机碳流失(图3)。高原鼠兔的挖掘活动使地表堆积大量松散和干燥的土丘,这些松散的土丘在外营力作用下会被逐渐侵蚀[47,79],导致土壤细粒物质减少,砾石含量增加,由于粘粉粒含量的损失量大于补偿量[80-81],从而引起土壤有机碳的损失[82]。最近在样地尺度的研究表明高原鼠兔挖掘导致地表堆积1~5 t·ha-1的疏松土壤,潜在的土壤有机碳流失量约为20~70 kg·ha-1[83]。尽管有学者就高原鼠丘风力侵蚀作用及土壤流失规律进行了野外人工模拟风蚀试验[84],但由于缺乏自然状态下鼠丘土壤侵蚀速率以及与鼠丘相关联的斑块演变的直接证据,导致就高原鼠兔挖掘活动对土壤有机碳的量化研究依然不足。例如,高原鼠兔干扰对土壤碳流失的直接贡献以及与斑块化相关联的间接贡献分别是多少?鼠丘以及斑块土壤侵蚀导致直接迁移出生态系统的碳损失量和由于沉降重新进入生态系统的返还量分别是多少?等这些关键科学问题还需要进一步的量化研究。此外,洞口土丘压伏牧草,导致其死亡引起植被碳的减少[85]。最新的研究表明高原鼠兔挖掘活动引起植被生物量的减少不足2%[86],而其年采食牧草量大约为高寒草地总生产力的21%~40%[83]。但是,由于缺少大量的样地尺度高原鼠兔种群密度数据,其采食活动对植被碳库的影响未见报道。此外,高原鼠兔采食活动导致群落物种组成以及群落物种间竞争发生改变[53,61,87],由此引发的植被超补偿生长量也没有得到量化。
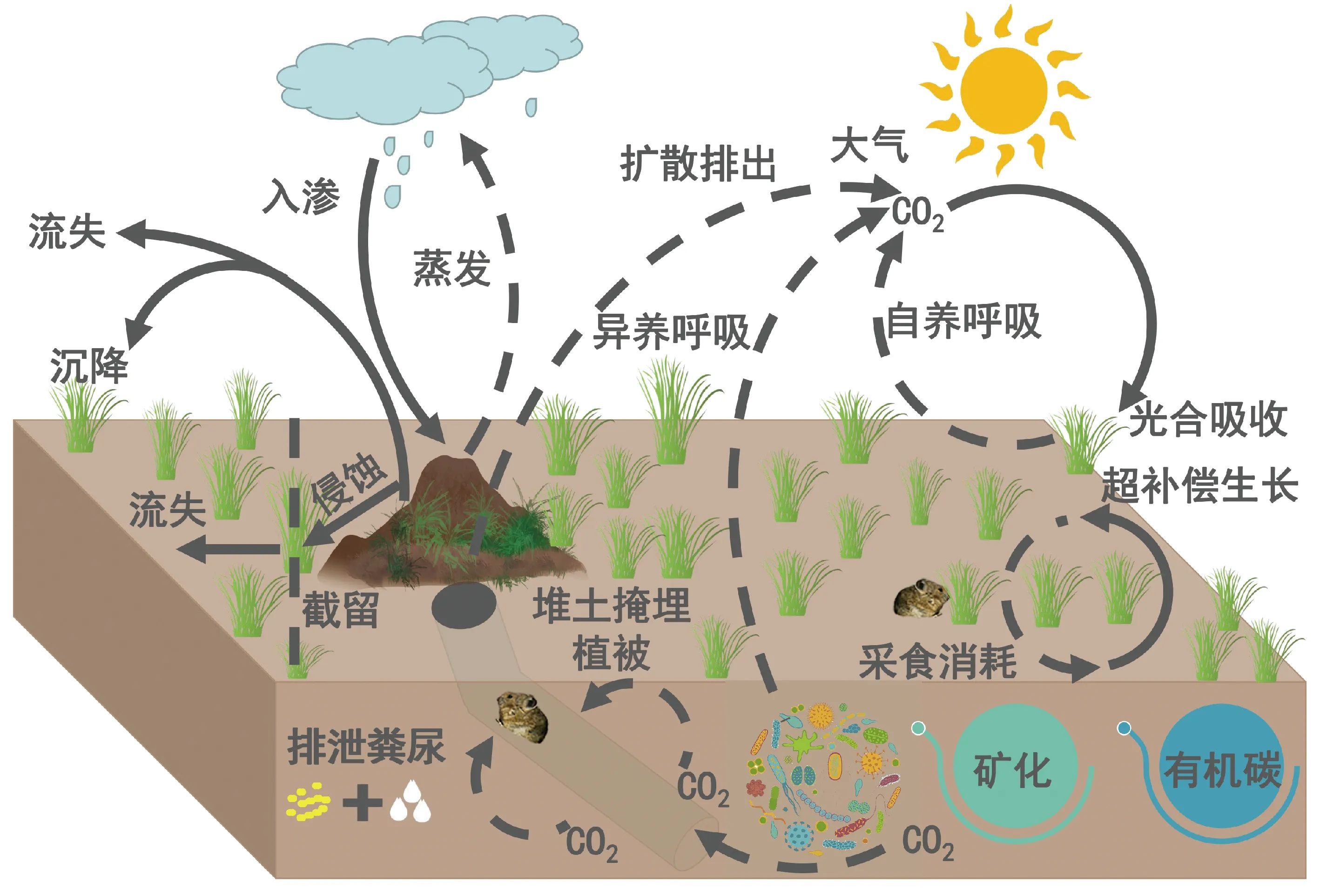
图3 高原鼠兔干扰对高寒草地碳储量和碳交换的影响Fig.3 Influence of pikas’ disturbance on carbon storage and exchange of alpine grassland
4 高原鼠兔干扰对生态系统碳收支的影响
生态系统呼吸(Ecosystem respiration,Re)是碳循环中仅次于生态系统总初级生产力(Gross primary productivity,GPP)的第二大通量组分[88]。净生态系统交换(Net ecosystem CO2exchange,NEE)受到生态系统呼吸作用导致的碳排放和光合作用形成的碳吸收两个过程的共同调节[89],它体现了生态系统的碳源或碳汇能力[90]。生态系统呼吸碳排放和光合作用碳吸收两个过程受到诸多生物和环境因子的控制[91]。因此,任何改变环境条件和影响植被生长的外部扰动都会导致生态系统碳交换发生改变,进而影响生态系统的碳收支。
洞穴是高原鼠兔栖居、繁殖、规避捕食风险和应对恶劣天气等所依赖的基本设施[46]。高原鼠兔挖掘洞穴打破了生态系统原有的碳交换平衡,加速了CO2通过鼠兔洞道扩散排出[92](图3)。伴随着采食活动改变了下垫面的微地形结构[19],这种异质性的微生境在短期内可以显著改善土壤的水分和养分环境[93-94]。但是从长期来看,由于鼠丘和裸地斑块缺少植被覆盖,不利于土壤养分和水分的保持[95]。最近的研究表明高原鼠兔干扰影响下生态系统呼吸底物供应不足和土壤暖干化等环境因子的改变会导致生态系统碳排放显著减少[21-22]。除了对限制植被生长的环境因子的影响外,高原鼠兔的干扰也改变了高寒草地的土壤微生物和植物群落结构[96-97]。高原鼠兔干扰导致植物物种组成和地上、地下生物量分配格局的变化是高寒草地净生态系统交换降低的主要原因[23-24](图3)。但是也有研究指出高原鼠兔干扰导致土壤呼吸速率增加10%,而土壤呼吸对温度的敏感性降低了大约20%[98]。
综上所述,高原鼠兔干扰改变了高寒草地生态系统呼吸和净生态系统交换过程,但是关于该方面的研究未能充分展开。例如,高原鼠兔干扰下生态系统碳排放和碳吸收哪个过程占主导地位?生态系统呼吸排放减少的碳是否可以抵消光合作用固定的碳损失?此外,高原鼠兔干扰下涉及碳排放和碳吸收这两个过程的关键驱动机制是什么?这些问题还需要进一步的深入研究。
5 展望
要客观辨析高原鼠兔在高寒草地碳循环中的作用,需要在大尺度评估其干扰对生态系统碳储量和碳交换的影响,其关键在于量化高原鼠兔种群密度及其扰动痕迹(新、旧鼠丘和裸地斑块)。但是目前关于大范围样地尺度高原鼠兔种群密度和扰动痕迹的研究还很匮乏[83,86]。缺乏行之有效的观测手段是造成以上研究不足的主要原因。高原鼠兔个体小且异常警觉,而其扰动痕迹形状和大小不一。在空间尺度上,传统样方或样带调查法费时费力,空间覆盖范围较大时,调查结果的空间代表性不足,而遥感反演数据又受到空间分辨率的限制,难以与样地尺度的实测结果精确匹配。在时间尺度上,遥感手段虽可重复获取数据,但受传感器重访周期、高原天气情况等的影响,获取高质量的时序数据也存在诸多困难。因此,这两种方法在获取大范围样地尺度高原鼠兔种群密度和高原鼠兔扰动痕迹时存在较大局限。
由于缺少大范围样地尺度、长时间序列高原鼠兔种群密度和高原鼠兔扰动痕迹数据,现有关于高原鼠兔干扰对高寒草地碳循环的影响工作大都是基于样方尺度(0.25~1 m2)短期的取样和观测研究,即在不同的高原鼠兔种群密度地区或者高原鼠兔扰动区域和非扰动区域设置样地(1 000~10 000 m2),基于样方法调查和取样分析不同鼠兔密度区土壤有机碳含量[99-100]、生产力[101-102]、生态系统呼吸和净交换过程[23-24]。这些研究结果反映了不同高原鼠兔种群密度区、扰动和非扰动区生态系统碳库和碳交换特征,对高原鼠兔驱动形成的异质性下垫面的影响考虑不足。尽管有研究分析了这种异质性下垫面的土壤有机碳和生态系统呼吸特征[21-22],但是依然缺乏针对不同下垫面净生态系统交换的研究。此外,青藏高原作为中国最大的草地畜牧业基地之一,放牧是高寒草地最主要的利用方式,而且超过一半的高寒草地地下存在着多年冻土。多年冻土和放牧是除高原鼠兔干扰外影响高寒草地的主要因素。因此,现有基于不同鼠兔种群密度区、扰动和非扰动区所得的结果可能会掩盖高原鼠兔对生态系统碳循环的真实影响。
近年来,研究者注意到为了更为客观、合理地评估高原鼠兔干扰对高寒草地碳收支的影响,需要量化高原鼠兔驱动形成的不同下垫面的比例,并将样方尺度的观测准确地升尺度到样地。为此,研究者在样地尺度估算了高原鼠兔扰动区和非扰动区土壤有机碳、无机碳和总碳储量[103],分析了高原鼠兔对高寒草地生物量[83]、植被盖度[104]和土壤理化性质[77]的影响。但是已有在样地尺度的研究并没有量化高原鼠兔驱动形成的下垫面对生态系统碳储量、碳排放和碳吸收的贡献,获取的研究结果无法评估高原鼠兔干扰对高寒草地碳收支的影响。因此,亟需在样地尺度开展兼顾碳库和碳交换的系统研究,这是合理评价高原鼠兔干扰对高寒草地生态系统碳收支和碳源汇功能影响的必要条件。
小型无人机技术在最近几年的发展十分迅速,并在各行各业得到了广泛的应用[105]。在草地研究和管理方面,无人机航拍可以提供高分辨率的地表影像,因而在样地尺度能够更准确地提取草地的植被盖度、斑块和鼠洞等信息[106]。在区域尺度高原鼠兔空间分布[107],高原鼠兔干扰对高寒草地的影响[58],高寒草地植物物种调查[108],生物量估算[109],生态系统碳储量和碳排放估算[66]等方面得到了广泛的应用。秦彧等[66]采用无人机航拍、遥感反演和样方取样调查相结合的方法在样地尺度分析了斑块对高寒草地生态系统碳储量和碳排放估算结果的影响,结果表明不考虑斑块的影响会导致对高寒草地生态系统碳储量和生态系统碳排放显著的高估,结合遥感反演,无人机航拍和样地调查可以提高高寒草地碳储量和排放估算的精度。因此,基于大量的无人机近地面航拍和实地调查相结合的方法,通过细化不同下垫面生态系统碳库和碳交换特征,可以明确高原鼠兔干扰对生态系统碳储量、碳排放和碳吸收的贡献量,提高高寒草地碳收支的评价精度。
